

Abstract
Gut-associated lymphoid tissue is supposed to play a central role in both the organization of colonic repair mechanisms and colorectal carcinogenesis. In inflammatory conditions, the number, diameter and density of isolated lymphoid follicles (ILFs) increases. They are not only involved in immune surveillance, but their presence is also indispensable in normal mucosal regeneration of the colon. In carcinogenesis, ILFs may play a dual role. On the one hand they may support tumor growth and the metastatic process by vascular endothelial growth factor receptor signaling and producing a specific cytokine and cellular milieu, but on the other hand their presence is sometimes associated with a better prognosis. The relation of ILFs to bone marrow derived stem cells, follicular dendritic cells, subepithelial myofibroblasts or crypt formation, which are all involved in mucosal repair and carcinogenesis, has not been directly studied. Data about the putative organizer role of ILFs is scattered in scientific literature.
Keywords: Isolated lymphoid follicle, Colon, Mucosal repair, Colorectal cancer, Epithelial stem cell, Myofibroblast, Follicular dendritic cell, Mesenchymal-epithelial transition, Epithelial-mesenchymal transition
INTRODUCTION
The imbalance of colonic epithelial proliferation and apoptosis may lead to both ulcer- and carcinoma development of the mucosa. The final direction of this imbalance depends on complex pathogenetic pathways in which isolated lymphoid follicles (ILFs) seem to have a specific role.
Some steps of colonic epithelial regeneration are known, but the connection among them is not fully understood. The continuous reformation of the epithelial layer is important in avoiding the aggregation of pernicious mutations induced by intraluminal factors. In inflammation, the lack of regenerative factors and the disturbance of the regulation of regenerative mechanisms favour ulcer development. It has also been observed that in colonic inflammation there is a tight connection between the degree of epithelial damage and the number, diameter and cellular compounds of subepithelial lymphoid follicles[1,2]. The more severe the epithelial destruction that develops, the higher the number of ILFs that can be found in adjacent mucosa.
It has recently been reported that lymphoid follicles are also present in carcinomas of the lung[3], endometrium[4], liver[5], and colon[1]. They are supposed to have immune-mediated anti-tumoral effects, as their elevated number is in positive correlation with a better prognosis and a longer survival[3]. However, the density of lymphoid follicle-associated flat dysplastic aberrant crypt foci was significantly higher compared to the rest of the mucosa in azoxymethane-treated rats[6]. Several reports have investigated the association between lymphoid aggregates and colonic tumors in rodents[7,8]. The results indicate that colonic crypts overlying ILFs show a significantly higher proliferative activity, which may also influence genetically defected epithelial cells. Hence, the risk of carcinoma is increased in the colonic mucosa of ILFs compared to mucosa without ILFs. It has also been shown that the incidence of ILFs in early human colorectal cancers significantly differs by gender, location, macroscopic type and histology, but moreover, their localization significantly differs by their macroscopic type[9].
However, the exact role of ILFs in colonic epithelial repair and colorectal carcinogenesis is not yet known. Some data show[10] that the lack of lymphoid follicles results in abnormal crypt formation in the case of epithelial destruction. On the other hand, Apc gene mutation causes impairment of developmental and apparent differentiation blockade in proliferative tissues, including those of the lymphoid follicles[11]. Whether ILFs act as a regenerative pool containing putative stem cells in case of mucosal damage, or they are responsible only for the optimal cytokine milieu for the differentiation of immigrating stem cells or invasive carcinoma cells[12] need to be further examined.
THE ORGANIZATION OF THE GUT-ASSOCIATED LYMPHOID TISSUE
The gut-associated lymphoid tissue (GALT) is a component of the mucosa-associated lymphoid tissue, in which approximately 70 percent of the body’s immune cells are found[13,14]. GALT differentiates between pathogens and commensal bacteria.
The majority of GALT is composed of isolated and aggregated lymphoid follicles dispersed throughout the small and large intestines[15]. These lymphoid follicles, including Peyer’s patches (PPs) of the small intestine and ILFs of the large intestine, are composed of a specialised follicle associated epithelium (FAE), which overlies a subepithelial dome containing numerous macrophages, dendritic cells, T, B lymphocytes, and special antigen sampling microfold/M/cells[15-17]. The FAE has a crucial role in the initiation of the mucosal and systemic immune response[18]. ILFs have, in general, an average diameter of 0.1-0.7 mm and number of around 30 000 in humans[19].
ILFs are innervated sites of GALT. Functionally, antigen-triggered mast cell and eosinophil activation affects both the secretory and motor functions of the intestines[20], and these defensive reactions can be modulated by the enteric nervous system[21]. It has been recently recognised that there is a dense neuronal network at the level of the supra-follicular dome region, but not within the germinal centers in lymphoid follicles[22]. Neuronal alterations of PPs and ILFs, such as nerve-eosinophil associations or increasing neuronal cell adhesion molecule expression, may have consequences on the uptake of particular pathogens[16,23].
VASCULARIZATION OF ILFS
ILFs have rich blood and lymphatic vascularization[14,19]. Vasculogenesis may play a dual role in mucosal organization, in that it is not only necessary for nutritional and metabolic processes, but the homing of the repopulating bone marrow derived stem cells to the site of tissue damage may happen via blood vessels. In the case of cancer development, the vascular system is essentially involved in tumor growth, invasion and metastasis formation.
Revascularization is a key point of colonic mucosal repair. During inflammatory stages, due to cytokine action and intercellular adhesion, molecules signalling some of the vessels differentiate into high endothelial venules (HEVs)[24,25]. In the case of lymphocytes and neutrophils, it is supposed that they firstly reach the inflammatory sites via a transcellular pathway through the HEVs[26], but an intercellular pathway is also known[27]. Upon epithelial injury the circulating bone marrow derived cells (BMDCs) migrate to the stromal layer of the damaged colonic wall, presumably via HEVs at an increased number regulated by overexpressed inflammatory chemokines[28].
Based on the result of Witmer et al[29], it has been suggested that in lymphoid tissues, including GALT, the signaling system of the vascular endothelial growth factor (VEGF) and its receptor play a permanent role in the vasculogenesis of ILFs. Whereas the inhibition of VEGF has shown promising results in sporadic colon cancer, it has been recently published that VEGF receptor signaling acts as a direct growth factor for tumor cells in colitis-associated cancer, providing a molecular link between inflammation and the development of colon cancer[30].
BONE MARROW DERIVED STEM CELLS OF ILFS
Based on the former results[31-33], emerging evidence suggests that bone marrow derived stem cells contribute to tissue regeneration partly by promoting neovascularization or arteriogenesis. After human hematopoietic cell transplantation epithelial tissue chimerism appears[34-36].
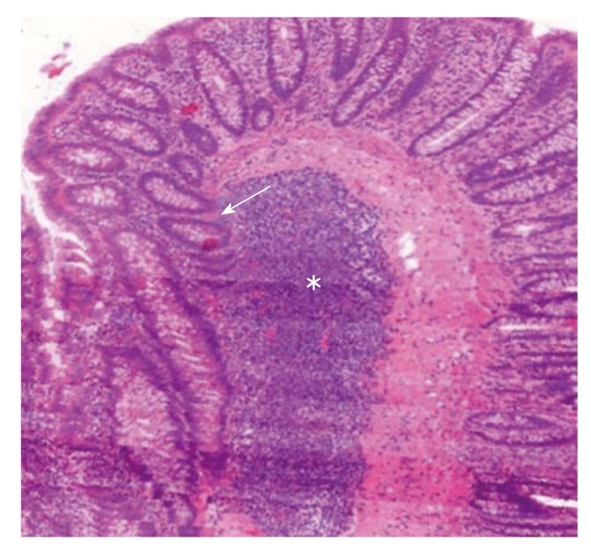
The bone marrow origin of epithelial cells may be supposed by observations in which epithelial cell markers and leukocyte markers showed that double positive cells were found in inflamed mucosa adjacent to lymphoid aggregates[2,37-39]. The presence of cytokeratin, epithelial growth factor receptor, hepatocyte-derived growth factor receptor or CDX2 co-expression in CD45+ cells of ILFs may support the mesenchymal origin of epithelial stem cells. Based on these results, it seems that ILFs are involved in the homing and differentiation of BMDCs in the case of colonic mucosal damage (Figure 1).
Figure 1.

Intraepithelial male donor bone marrow origin CD45-/Y-FISH+ cell (white arrow) and CD45+/Y-FISH+ intraepithelial lymphocyte (red arrow) in the colonic biopsy specimen of a female acceptor. A: Chromosomal detection (green: Y-chromosome, red: X-chromosome; fluorescence in situ hybridization); B: CD45 and cytokeratin (green: cytokeratin, red: CD45; fluorescence immunohistochemistry; 130 × magnification).
The cause of metastasis remains elusive despite a vast amount of information on cancer cells. According to recent research, cancer cell fusion with macrophages or immigrating BMDCs provides an explanation[40,41]. BMDCs fused with tumor cells were present not just in animal tumor xenografts where they were associated with metastases, but in human carcinomas, including colon cancer. BMDC-tumor cell fusion explains the epidermal-mesenchymal transition in cancer since BMDCs express mesodermal traits and epithelial-mesenchymal transition regulators (i.e.: Twist, SPARC). If BMDC-tumor cell fusion underlies invasion and metastasis in human cancer, new therapeutic strategies would be mandated.
DENDRITIC CELLS IN ILFS
Follicular dendritic cells (FDCs) in lymphoid follicles retain native antigens in the form of immune complexes on their membrane for months, and present these antigens to B cells during the secondary response[42,43]. The origin and cell lineage of FDCs are controversial. Whereas their immune functions and expression of hemopoietic cell-associated antigens suggest that they belong to the hemopoietic lineage[44], their spindle-shaped morphology “in vitro”, lack of CD45, and presence of antigens expressed by fibroblasts[45] indicate that FDCs may be mesenchymal cells. Based on studies with mouse radiation chimeras, Humphrey et al[46] concluded that FDCs were not derived from the bone marrow, but came from a local mesenchymal precursor. However, Kapasi et al[44], using mice homozygous for the SCID mutation, which lack T, B lymphocytes, and FDCs, demonstrated that after reconstitution with bone marrow from donor mice, the FDCs of the reconstituted mice expressed the donor phenotype. These authors concluded that FDC precursors came from bone marrow.
According to the results of Muñoz-Fernández et al[47], FDCs seem to be a specialized form of myofibroblasts and derive from bone marrow stromal cell progenitors. The authors were able to isolate and culture 18 follicular dendritic cell lines from human tonsils. These cells were CD45-negative and expressed antigens associated with FDCs (CD21, CD23, CD35, CD40, CD73, BAFF, ICAM-1, and VCAM-1) and antigens specific for FDC (DRC-1, CNA.42, and HJ2). These cell lines were also able to bind B cells and secrete CXCL13, and they had functional activities characteristic of FDCs. Nevertheless, the additional expression of STRO-1, together with CD10, CD13, CD29, CD34, CD63, CD73, CD90, ICAM-1, VCAM-1, HLA-DR, alkaline phosphatase, and α-smooth muscle actin (α-SMA) indicated that FDCs are closely related to bone marrow stromal cell progenitors. The expression of α-SMA also relates FDCs with myofibroblasts. Like myofibroblasts, FDC lines expressed stress fibers containing α-SMA and were able to contract collagen gels under the effect of TGFβ1 and platelet-derived growth factor.
In various inflammation models, tissue-derived dendritic cells have been shown to migrate from the inflammatory site via lymphatics to secondary lymphoid organs where they interact with lymphocytes[48]. Based on their dual phenotype, follicular dendritic cells may represent a transformation switch point among immigrating bone marrow derived stem cells in ILFs and the surrounding subepithelial myofibroblasts.
The origin of dendritic cells (DCs) in tumors remains obscure. Recent studies indicate that conventional DCs in lymphoid tissues arise from a distinct population of committed conventional DC precursors (pre-cDCs) that originate in bone marrow and migrate via blood. Diao et al[49] showed that pre-cDCs are precursors for conventional DCs in tumors, and they migrate from blood into the tumor where they generate conventional DCs. The chemokine CCL3, which is markedly upregulated in tumors (including colon cancer) and in tumor-infiltrating stromal and immune cells, promotes pre-cDC recruitment. Both pre-cDCs and their conventional DC progeny actively proliferate within the tumor, and have the ability to mature and stimulate Ag-specific lymphocytes. This finding suggests that in several cases the migration of pre-cDCs to tumors may represent a normal response to inflammation. Further studies are needed to delineate the role of pre-cDCs in other inflammatory processes and to compare them with monocytes, which are currently considered the main source of inflammatory DCs in peripheral tissues[50,51].
MYOFIBROBLASTS SORROUNDING ILF ADJACENT EPITHELIUM
Subepithelial myofibroblasts (SEMFs) exist as a syncytium that extends throughout the colonic lamina propria, merging with the pericytes surrounding the blood vessels[52,53]. SEMFs are involved in two epithelial repair processes[54,55]. One process is called restitution[56]. This is an important response to minor to moderate injury. The other process is observed when the wound is deep, and the subepithelial tissues and the basement membrane need to be reconstituted[55].
According to recent studies[54,57,58], myofibroblasts are thought to derive from two major sources, bone marrow or locally activated fibroblasts, in response to transforming growth factor-β1. In the case of serious tissue injury (i.e. active ulcerative colitis) the regeneration capacity of local stem cells is not enough to complete tissue repair. In this case, bone marrow derived mesenchymal stem cells migrate into the gastrointestinal wall where they may contribute to the repair progress[59,60] as differentiated mesenchymal cells (e.g. myofibroblasts)[61].
Despite the increasing number of publications illustrating the role of tumor-associated stromal cells in cancer progression, there still exists a significant ambiguity with respect to the identification of cancer-associated fibroblasts, myofibroblasts and peritumoral fibroblasts in the cancer tissue. SEMFs appear early in the cancer’s development. The mutual interaction (through direct cell-cell contacts and paracrine signals) between cancer cells and SEMFs is essential for invasive growth and is translated into a poor clinical prognosis[62].
TOLL-LIKE RECEPTOR EXPRESSION IN ILFS
Beside immune functions, PPs and ILFs are supposed to be involved in mucosal repair via Toll-like receptors (TLRs). In ILFs, TLRs are expressed on the cells of the monocyte/macrophage system, on some kinds of T cells, as well as on intestinal epithelial, endothelial and stromal cells[63]. Using the dextran sodium sulfate (DSS) model of colitis, mice lacking TLR2, TLR4 or MyD88 all developed more severe colitis than wild type mice when exposed to orally administered DSS[64]. These findings suggest that signaling from commensal bacteria throughout TLRs resulted in protection from DSS colitis through enhanced epithelial cell proliferation, and worked as a compensatory factor against epithelial damage[64].
TLRs can also bind endogenous ligands including necrotic cells, heat shock proteins, and extracellular matrix components[65-67]. Necrotic cells may activate NF-κB through TLR2, leading to the expression of tissue repair-associated genes[65]. It is supposed that necrosis induced inflammation in tissue damage may provide danger signals functioning as inducers of tissue repair responses through TLRs. The TLR ligands released from necrotic cells have not been identified, although heat shock proteins produced by damaged cells are known to be TLR ligands[66]. Components of the extracellular matrix, such as hyaluronan, can be an endogenous ligand for TLR4[67]. Increased hyaluronan production has been demonstrated in both DSS colitis in mice and in human Crohn’s disease[68]. It is possible that TLR activation may occur in the absence of microbial products[68]. In the case of inflammatory mucosal damage, ILFs may induce repair mechanisms via endogenous TLR activation.
TLR4 was also shown to be expressed on human colon carcinoma cells and functionally active. It may play important roles in promoting immune escape of human colon carcinoma cells by inducing immunosuppressive factors and apoptosis resistance, and it may also promote the proliferation and migration of cancer cells[69,70].
The analysis of isolated tumor cells from primary colon cancers showed co-expression of TLR7 and TLR8 with CD133 and gave evidence for a subpopulation of colon cancer-initiating cells[71]. Persistent TLR-specific activation of NF-κB in colorectal cancer, particularly in tumor-initiating cells, may sustain further tumor growth and progression through perpetuated signaling known from inflammatory and tissue repair mechanisms with consecutive self-renewal in pluripotent tumor cells. Activation through self-ligands or viral RNA fragments from tumor-associated lymphoid aggregates may putatively maintain this inflammatory process, suggesting a key role in cancer progression.
THE EFFECT OF THE PRESENCE OF ILFS ON MUCOSAL REPAIR
The epithelia of intestinal crypts associated with ILFs and PPs have an increased proliferation rate[10,14]. Saxena et al[10] showed that PPs in rats have a facilitative effect on the healing of intestinal wounds by promoting both epithelial cell migration on the defect and epithelial cell proliferation in the crypts adjacent to the wound and by decreasing the rate of wound contraction.
In rats, a difference in epithelial apoptosis between the FAE of PPs and intestinal villi was described[72]. Onishi et al[72] showed that the progression of the apoptotic process in the epithelial cells of FAE occurs later than in the intestinal villi, so the possibility of epithelial differentiation might remain in FAE, unlike in intestinal villi. PPs are supposed to have a regulatory effect on the epithelial proliferation as well[73].
The Wnt signaling pathway is critical for regulating a number of basic cell functions, such as cell proliferation, cell fate, polarity, differentiation, and migration, leading to morphogenesis and organogenesis[74,75]. There is strong genetic evidence that Wnt signaling play critical roles in the regulation of epithelial stem cells in the intestinal tract[76]. The Wnt target gene Lgr5 has been recently identified as a novel stem cell marker of the intestinal epithelium and the hair follicle[77]. In the intestine, Lgr5 is exclusively expressed in cycling crypt base columnar cells[77]. Many Wnt family proteins are expressed in hematopoietic tissues, and can also be secreted by lymphoid cells[78,79]. The Wnt-Lgr5 pathway may be a potential switch between the ILFs and colonic epithelial renewal. Lymphoid cells of ILFs may produce Wnts which are essential components of a milieu in which bone marrow derived stem cells immigrated to ILFs to engage in epithelial differentiation (Figure 2).
Figure 2.
3D reconstruction of a human colonic surgical sample (MIRAX Viewer, 3D, 3DHISTECH Ltd., Budapest). A large subepithelial isolated lymphoid follicle (white star) can be seen. Colonic crypts (white arrow) with no connection to the luminal surface “outgrow” from the isolated lymphoid follicle.
THE EFFECT OF THE PRESENCE OF ILFS ON COLORECTAL CARCINOGENESIS
Results from experimental colon cancer studies indicated that ILFs might promote the development of adenocarcinomas[7,8]. However, studies in experimental animals have also shown that the intestinal lymphoid system plays an important role in immunologic defense mechanisms; that is, antigenic stimuli result in germinal center formation, antibody production, and finally enlargement of the follicles[80]. In humans, the presence of tumor-infiltrating lymphocytes is associated with an improved prognosis in colorectal cancers, as does the presence of high level DNA microsatellite instability[81]. These results suggest that ILFs in early colorectal neoplasms play an important role in defense rather than in promotion.
In a recent study, Fu et al[9] found that the incidence of ILFs in early human colorectal neoplasms significantly differs by gender, location, macroscopic type, and histology, but moreover, their localization significantly differs by macroscopic type.
In squamous cell carcinomas of the esophagus, cyclin A expression in the germinal center cells of ILFs beneath the superficial tumorous lesions was shown to be an immunological signal toward the proliferation and progression of the tumors[82]. Gutfeld et al[83] found that the cells of colonic ILFs, inflammatory cells, ganglion cells, and endothelial cells express serum amyloid A, an acute phase reactant, whose level in the blood is elevated in response to trauma, infection, inflammation, and neoplasia, on both mRNA and protein levels. The serum amyloid A mRNA expression in epithelial cells was found to gradually increase as it progressed through different stages of dysplasia to overt carcinoma. While expression of the serum amyloid A1 and -4 genes in colon carcinomas was confirmed by RT-PCR analysis, this expression was barely detectable in normal colon tissues. Their findings indicate local and differential expression of serum amyloid A in human colon cancer and tumor-associated ILFs, and suggest its role in colorectal carcinogenesis.
MESENCHYMAL-EPITHELIAL AND EPITHELIAL-MESENCHYMAL TRANSITION IN ILFS
Epithelial-mesenchymal transition (EMT) is a physiological mechanism present during development, and is also encountered in several pathological situations such as renal interstitial fibrosis, endometrial adhesion, and cancer metastasis[84]. A reverse phenomenon, mesenchymal-epithelial transition (MET) also takes place during normal development in processes such as somitogenesis, kidney development and coelomic cavity formation[85]. In adult organisms, it has been proposed that restrictive mechanisms repress EMT and MET[86]. During tumor development, these mechanisms appear to fail, allowing EMT described in metastasis generation[87].
In inflammation, MET can also be altered because mesenchymal stem cells are mobilized to these sites of injury and consequently subjected to the inflammatory response[88]. BMDCs could differentiate into mature-appearing epithelial cells in response to tissue damage[89]. It was recently published that versican, a large chondroitin sulfate proteoglycan, mediates MET[90]. The results of Hirose et al[91] indicate that versican can bind specific chemokines through its chondroitin sulfate chains and that the binding tends to down-regulate the chemokine function. This raises the possibility that versican may act as a regenerative factor in colonic mucosa, and may be an important switch point between ILFs and MET. The presence of CDX2 and cytokeratin positive subepithelial cells in the marginal zone of ILFs also suggests that MET may take place in these immune formations[2].
Stroma-tissue, including lymphoid aggregates and ILFs surrounding the cancer cells, plays an important role in the tumor behavior. Mesker et al[92] analyzed the expression of markers involved in pathways related to stroma production and EMT (β-catenin, TGF-β-R2, SMAD4) in high-risk colorectal cancer patients, and found that patients with stroma-high and SMAD4 loss are of high risk. The anti-EMT effect of SMAD4 was also proven in colon carcinoma cells[93].
CONCLUSION
Based on the summarized results of literature, it seems that ILFs act like a switch between colonic mucosal regeneration and colorectal carcinogenesis.
Subepithelial revascularization after mucosal damage takes place partly under the direction of ILFs with the prominent help of vascular endothelial growth factor and its receptors. Immigrating stem cells from bone marrow may leave circulation via high endothelial venules in ILFs and their surroundings. Their differentiation throughout mesenchymal-to-epithelial transition may also happen in ILFs, and follicular dendritic cells, as well as the subepithelial myofibroblasts, seem to be crucial parts of colonic crypt formation and epithelial renewal.
Vasculogenesis in ILFs supports not just tumor growth and the metastatic process, but the VEGF receptor signaling acts like a direct growth factor for tumor cells. The fusion of BMDCs immigrating to ILFs with tumor cells may explain EMT in colorectal cancers. The presence of ILFs, dendritic cells and subepithelial myofibroblasts may also result in a specific milieu for tumor formation, growth and invasion.
Better understanding of the role of ILFs in mucosal repair may lead to the development of new therapeutic agents for inflammatory colon diseases that not only decrease the activity of inflammation, but also accelerate epithelial barrier recovery, hence dramatically decreasing clinical symptoms. Moreover, by revealing the exact connections between ILFs and colorectal carcinogenesis, the basis of individualized anti-cancer immunotherapies may be established.
Footnotes
Peer reviewer: Akira Andoh, MD, Department of Internal Medicine, Shiga University of Medical Science, Seta Tukinowa, Otsu 520-2192, Japan
S- Editor Tian L L- Editor Rutherford A E- Editor Ma WH
References
- 1.Nascimbeni R, Di Fabio F, Di Betta E, Mariani P, Fisogni S, Villanacci V. Morphology of colorectal lymphoid aggregates in cancer, diverticular and inflammatory bowel diseases. Mod Pathol. 2005;18:681–685. doi: 10.1038/modpathol.3800343. [DOI] [PubMed] [Google Scholar]
- 2.Sipos F, Muzes G, Valcz G, Galamb O, Tóth K, Leiszter K, Krenács T, Tulassay Z, Molnár B. Regeneration associated growth factor receptor and epithelial marker expression in lymphoid aggregates of ulcerative colitis. Scand J Gastroenterol. 2010;45:440–448. doi: 10.3109/00365521003624144. [DOI] [PubMed] [Google Scholar]
- 3.Dieu-Nosjean MC, Antoine M, Danel C, Heudes D, Wislez M, Poulot V, Rabbe N, Laurans L, Tartour E, de Chaisemartin L, et al. Long-term survival for patients with non-small-cell lung cancer with intratumoral lymphoid structures. J Clin Oncol. 2008;26:4410–4417. doi: 10.1200/JCO.2007.15.0284. [DOI] [PubMed] [Google Scholar]
- 4.Shia J, Black D, Hummer AJ, Boyd J, Soslow RA. Routinely assessed morphological features correlate with microsatellite instability status in endometrial cancer. Hum Pathol. 2008;39:116–125. doi: 10.1016/j.humpath.2007.05.022. [DOI] [PubMed] [Google Scholar]
- 5.Wada Y, Nakashima O, Kutami R, Yamamoto O, Kojiro M. Clinicopathological study on hepatocellular carcinoma with lymphocytic infiltration. Hepatology. 1998;27:407–414. doi: 10.1002/hep.510270214. [DOI] [PubMed] [Google Scholar]
- 6.Paulsen JE, Steffensen IL, Olstørn HB, Alexander J. Prevalent location of flat dysplastic aberrant crypt foci near lymphoid follicles in the colon of azoxymethane-treated rats. Anticancer Res. 2006;26:1803–1807. [PubMed] [Google Scholar]
- 7.Hardman WE, Cameron IL. Colonic crypts located over lymphoid nodules of 1,2-dimethylhydrazine-treated rats are hyperplastic and at high risk of forming adenocarcinomas. Carcinogenesis. 1994;15:2353–2361. doi: 10.1093/carcin/15.10.2353. [DOI] [PubMed] [Google Scholar]
- 8.Nauss KM, Locniskar M, Pavlina T, Newberne PM. Morphology and distribution of 1,2-dimethylhydrazine dihydrochloride-induced colon tumors and their relationship to gut-associated lymphoid tissue in the rat. J Natl Cancer Inst. 1984;73:915–924. [PubMed] [Google Scholar]
- 9.Fu KI, Sano Y, Kato S, Fujii T, Koba I, Yoshino T, Ochiai A, Yoshida S, Fujimori T. Incidence and localization of lymphoid follicles in early colorectal neoplasms. World J Gastroenterol. 2005;11:6863–6866. doi: 10.3748/wjg.v11.i43.6863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Saxena SK, Thompson JS, Sharp JG. Role of organized intestinal lymphoid aggregates in intestinal regeneration. J Invest Surg. 1997;10:97–103. doi: 10.3109/08941939709032139. [DOI] [PubMed] [Google Scholar]
- 11.You S, Ohmori M, Peña MM, Nassri B, Quiton J, Al-Assad ZA, Liu L, Wood PA, Berger SH, Liu Z, et al. Developmental abnormalities in multiple proliferative tissues of Apc(Min/+) mice. Int J Exp Pathol. 2006;87:227–236. doi: 10.1111/j.1365-2613.2006.00477.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Raclot G, Devalland C, Potet F, Bernard F, Monath C. [Colonic carcinoma arising de novo near a lymphoid follicle] Gastroenterol Clin Biol. 1991;15:775–776. [PubMed] [Google Scholar]
- 13.Heel KA, McCauley RD, Papadimitriou JM, Hall JC. Review: Peyer’s patches. J Gastroenterol Hepatol. 1997;12:122–136. doi: 10.1111/j.1440-1746.1997.tb00395.x. [DOI] [PubMed] [Google Scholar]
- 14.Sipos F, Muzes G, Galamb O, Spisák S, Krenács T, Tóth K, Tulassay Z, Molnár B. The possible role of isolated lymphoid follicles in colonic mucosal repair. Pathol Oncol Res. 2010;16:11–18. doi: 10.1007/s12253-009-9181-x. [DOI] [PubMed] [Google Scholar]
- 15.Neutra MR, Mantis NJ, Kraehenbuhl JP. Collaboration of epithelial cells with organized mucosal lymphoid tissues. Nat Immunol. 2001;2:1004–1009. doi: 10.1038/ni1101-1004. [DOI] [PubMed] [Google Scholar]
- 16.O’Brien LM, Fitzpatrick E, Baird AW, Campion DP. Eosinophil-nerve interactions and neuronal plasticity in rat gut associated lymphoid tissue (GALT) in response to enteric parasitism. J Neuroimmunol. 2008;197:1–9. doi: 10.1016/j.jneuroim.2008.04.002. [DOI] [PubMed] [Google Scholar]
- 17.Neutra MR, Frey A, Kraehenbuhl JP. Epithelial M cells: gateways for mucosal infection and immunization. Cell. 1996;86:345–348. doi: 10.1016/s0092-8674(00)80106-3. [DOI] [PubMed] [Google Scholar]
- 18.Niedergang F, Kraehenbuhl JP. Much ado about M cells. Trends Cell Biol. 2000;10:137–141. doi: 10.1016/s0962-8924(00)01731-1. [DOI] [PubMed] [Google Scholar]
- 19.Azzali G. Structure, lymphatic vascularization and lymphocyte migration in mucosa-associated lymphoid tissue. Immunol Rev. 2003;195:178–189. doi: 10.1034/j.1600-065x.2003.00072.x. [DOI] [PubMed] [Google Scholar]
- 20.Baird AW, Cuthbert AW. Neuronal involvement in type 1 hypersensitivity reactions in gut epithelia. Br J Pharmacol. 1987;92:647–655. doi: 10.1111/j.1476-5381.1987.tb11368.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.O’Malley KE, Sloan T, Joyce P, Baird AW. Type I hypersensitivity reactions in intestinal mucosae from rats infected with Fasciola hepatica. Parasite Immunol. 1993;15:449–453. doi: 10.1111/j.1365-3024.1993.tb00630.x. [DOI] [PubMed] [Google Scholar]
- 22.Vulchanova L, Casey MA, Crabb GW, Kennedy WR, Brown DR. Anatomical evidence for enteric neuroimmune interactions in Peyer’s patches. J Neuroimmunol. 2007;185:64–74. doi: 10.1016/j.jneuroim.2007.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chiocchetti R, Mazzuoli G, Albanese V, Mazzoni M, Clavenzani P, Lalatta-Costerbosa G, Lucchi ML, Di Guardo G, Marruchella G, Furness JB. Anatomical evidence for ileal Peyer’s patches innervation by enteric nervous system: a potential route for prion neuroinvasion? Cell Tissue Res. 2008;332:185–194. doi: 10.1007/s00441-008-0583-y. [DOI] [PubMed] [Google Scholar]
- 24.Freemont AJ, Ford WL. Functional and morphological changes in post-capillary venules in relation to lymphocytic infiltration into BCG-induced granulomata in rat skin. J Pathol. 1985;147:1–12. doi: 10.1002/path.1711470102. [DOI] [PubMed] [Google Scholar]
- 25.Cavender DE, Haskard DO, Joseph B, Ziff M. Interleukin 1 increases the binding of human B and T lymphocytes to endothelial cell monolayers. J Immunol. 1986;136:203–207. [PubMed] [Google Scholar]
- 26.Perry ME, Mustafa Y, Brown KA. The microvasculature of the human palatine tonsil and its role in the homing of lymphocytes. Adv Otorhinolaryngol. 1992;47:11–15. doi: 10.1159/000421711. [DOI] [PubMed] [Google Scholar]
- 27.Kvietys PR, Sandig M. Neutrophil diapedesis: paracellular or transcellular? News Physiol Sci. 2001;16:15–19. doi: 10.1152/physiologyonline.2001.16.1.15. [DOI] [PubMed] [Google Scholar]
- 28.Chamberlain G, Fox J, Ashton B, Middleton J. Concise review: mesenchymal stem cells: their phenotype, differentiation capacity, immunological features, and potential for homing. Stem Cells. 2007;25:2739–2749. doi: 10.1634/stemcells.2007-0197. [DOI] [PubMed] [Google Scholar]
- 29.Witmer AN, Dai J, Weich HA, Vrensen GF, Schlingemann RO. Expression of vascular endothelial growth factor receptors 1, 2, and 3 in quiescent endothelia. J Histochem Cytochem. 2002;50:767–777. doi: 10.1177/002215540205000603. [DOI] [PubMed] [Google Scholar]
- 30.Waldner MJ, Wirtz S, Jefremow A, Warntjen M, Neufert C, Atreya R, Becker C, Weigmann B, Vieth M, Rose-John S, et al. VEGF receptor signaling links inflammation and tumorigenesis in colitis-associated cancer. J Exp Med. 2010;207:2855–2868. doi: 10.1084/jem.20100438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rafii S, Lyden D. Therapeutic stem and progenitor cell transplantation for organ vascularization and regeneration. Nat Med. 2003;9:702–712. doi: 10.1038/nm0603-702. [DOI] [PubMed] [Google Scholar]
- 32.Takahashi T, Kalka C, Masuda H, Chen D, Silver M, Kearney M, Magner M, Isner JM, Asahara T. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med. 1999;5:434–438. doi: 10.1038/7434. [DOI] [PubMed] [Google Scholar]
- 33.MacDermott RP, Sanderson IR, Reinecker HC. The central role of chemokines (chemotactic cytokines) in the immunopathogenesis of ulcerative colitis and Crohn’s disease. Inflamm Bowel Dis. 1998;4:54–67. doi: 10.1097/00054725-199802000-00009. [DOI] [PubMed] [Google Scholar]
- 34.Spyridonidis A, Schmitt-Gräff A, Tomann T, Dwenger A, Follo M, Behringer D, Finke J. Epithelial tissue chimerism after human hematopoietic cell transplantation is a real phenomenon. Am J Pathol. 2004;164:1147–1155. doi: 10.1016/S0002-9440(10)63203-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Körbling M, Katz RL, Khanna A, Ruifrok AC, Rondon G, Albitar M, Champlin RE, Estrov Z. Hepatocytes and epithelial cells of donor origin in recipients of peripheral-blood stem cells. N Engl J Med. 2002;346:738–746. doi: 10.1056/NEJMoa3461002. [DOI] [PubMed] [Google Scholar]
- 36.Okamoto R, Yajima T, Yamazaki M, Kanai T, Mukai M, Okamoto S, Ikeda Y, Hibi T, Inazawa J, Watanabe M. Damaged epithelia regenerated by bone marrow-derived cells in the human gastrointestinal tract. Nat Med. 2002;8:1011–1017. doi: 10.1038/nm755. [DOI] [PubMed] [Google Scholar]
- 37.Gould VE, Bloom KJ, Franke WW, Warren WH, Moll R. Increased numbers of cytokeratin-positive interstitial reticulum cells (CIRC) in reactive, inflammatory and neoplastic lymphadenopathies: hyperplasia or induced expression? Virchows Arch. 1995;425:617–629. doi: 10.1007/BF00199352. [DOI] [PubMed] [Google Scholar]
- 38.Sipos F, Molnár B, Zágoni T, Berczi L, Tulassay Z. Growth in epithelial cell proliferation and apoptosis correlates specifically to the inflammation activity of inflammatory bowel diseases: ulcerative colitis shows specific p53- and EGFR expression alterations. Dis Colon Rectum. 2005;48:775–786. doi: 10.1007/s10350-004-0831-5. [DOI] [PubMed] [Google Scholar]
- 39.Sipos F, Galamb O, Herszényi L, Molnár B, Solymosi N, Zágoni T, Berczi L, Tulassay Z. Elevated insulin-like growth factor 1 receptor, hepatocyte growth factor receptor and telomerase protein expression in mild ulcerative colitis. Scand J Gastroenterol. 2008;43:289–298. doi: 10.1080/00365520701714434. [DOI] [PubMed] [Google Scholar]
- 40.Pawelek JM, Chakraborty AK. The cancer cell--leukocyte fusion theory of metastasis. Adv Cancer Res. 2008;101:397–444. doi: 10.1016/S0065-230X(08)00410-7. [DOI] [PubMed] [Google Scholar]
- 41.Pawelek JM, Chakraborty AK. Fusion of tumour cells with bone marrow-derived cells: a unifying explanation for metastasis. Nat Rev Cancer. 2008;8:377–386. doi: 10.1038/nrc2371. [DOI] [PubMed] [Google Scholar]
- 42.van Nierop K, de Groot C. Human follicular dendritic cells: function, origin and development. Semin Immunol. 2002;14:251–257. doi: 10.1016/s1044-5323(02)00057-x. [DOI] [PubMed] [Google Scholar]
- 43.Park CS, Choi YS. How do follicular dendritic cells interact intimately with B cells in the germinal centre? Immunology. 2005;114:2–10. doi: 10.1111/j.1365-2567.2004.02075.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kapasi ZF, Qin D, Kerr WG, Kosco-Vilbois MH, Shultz LD, Tew JG, Szakal AK. Follicular dendritic cell (FDC) precursors in primary lymphoid tissues. J Immunol. 1998;160:1078–1084. [PubMed] [Google Scholar]
- 45.Lindhout E, de Groot C. Follicular dendritic cells and apoptosis: life and death in the germinal centre. Histochem J. 1995;27:167–183. [PubMed] [Google Scholar]
- 46.Humphrey JH, Grennan D, Sundaram V. The origin of follicular dendritic cells in the mouse and the mechanism of trapping of immune complexes on them. Eur J Immunol. 1984;14:859–864. doi: 10.1002/eji.1830140916. [DOI] [PubMed] [Google Scholar]
- 47.Muñoz-Fernández R, Blanco FJ, Frecha C, Martín F, Kimatrai M, Abadía-Molina AC, García-Pacheco JM, Olivares EG. Follicular dendritic cells are related to bone marrow stromal cell progenitors and to myofibroblasts. J Immunol. 2006;177:280–289. doi: 10.4049/jimmunol.177.1.280. [DOI] [PubMed] [Google Scholar]
- 48.Jakubzick C, Bogunovic M, Bonito AJ, Kuan EL, Merad M, Randolph GJ. Lymph-migrating, tissue-derived dendritic cells are minor constituents within steady-state lymph nodes. J Exp Med. 2008;205:2839–2850. doi: 10.1084/jem.20081430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Diao J, Zhao J, Winter E, Cattral MS. Recruitment and differentiation of conventional dendritic cell precursors in tumors. J Immunol. 2010;184:1261–1267. doi: 10.4049/jimmunol.0903050. [DOI] [PubMed] [Google Scholar]
- 50.Auffray C, Fogg DK, Narni-Mancinelli E, Senechal B, Trouillet C, Saederup N, Leemput J, Bigot K, Campisi L, Abitbol M, et al. CX3CR1+ CD115+ CD135+ common macrophage/DC precursors and the role of CX3CR1 in their response to inflammation. J Exp Med. 2009;206:595–606. doi: 10.1084/jem.20081385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Le Borgne M, Etchart N, Goubier A, Lira SA, Sirard JC, van Rooijen N, Caux C, Aït-Yahia S, Vicari A, Kaiserlian D, et al. Dendritic cells rapidly recruited into epithelial tissues via CCR6/CCL20 are responsible for CD8+ T cell crosspriming in vivo. Immunity. 2006;24:191–201. doi: 10.1016/j.immuni.2006.01.005. [DOI] [PubMed] [Google Scholar]
- 52.Powell DW, Mifflin RC, Valentich JD, Crowe SE, Saada JI, West AB. Myofibroblasts. II. Intestinal subepithelial myofibroblasts. Am J Physiol. 1999;277:C183–C201. doi: 10.1152/ajpcell.1999.277.2.C183. [DOI] [PubMed] [Google Scholar]
- 53.Joyce NC, Haire MF, Palade GE. Morphologic and biochemical evidence for a contractile cell network within the rat intestinal mucosa. Gastroenterology. 1987;92:68–81. doi: 10.1016/0016-5085(87)90841-9. [DOI] [PubMed] [Google Scholar]
- 54.Andoh A, Bamba S, Fujiyama Y, Brittan M, Wright NA. Colonic subepithelial myofibroblasts in mucosal inflammation and repair: contribution of bone marrow-derived stem cells to the gut regenerative response. J Gastroenterol. 2005;40:1089–1099. doi: 10.1007/s00535-005-1727-4. [DOI] [PubMed] [Google Scholar]
- 55.Wallace JL, Granger DN. The cellular and molecular basis of gastric mucosal defense. FASEB J. 1996;10:731–740. doi: 10.1096/fasebj.10.7.8635690. [DOI] [PubMed] [Google Scholar]
- 56.Podolsky DK. Healing the epithelium: solving the problem from two sides. J Gastroenterol. 1997;32:122–126. doi: 10.1007/BF01213309. [DOI] [PubMed] [Google Scholar]
- 57.Brittan M, Chance V, Elia G, Poulsom R, Alison MR, MacDonald TT, Wright NA. A regenerative rolefor bone marrow following experimental colitis: contribution to neovasculogenesis and myofibroblasts. Gastroenterology. 2005;128:1984–1995. doi: 10.1053/j.gastro.2005.03.028. [DOI] [PubMed] [Google Scholar]
- 58.Fritsch C, Simon-Assmann P, Kedinger M, Evans GS. Cytokines modulate fibroblast phenotype and epithelial-stroma interactions in rat intestine. Gastroenterology. 1997;112:826–838. doi: 10.1053/gast.1997.v112.pm9041244. [DOI] [PubMed] [Google Scholar]
- 59.Brittan M, Wright NA. Stem cell in gastrointestinal structure and neoplastic development. Gut. 2004;53:899–910. doi: 10.1136/gut.2003.025478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tanaka F, Tominaga K, Ochi M, Tanigawa T, Watanabe T, Fujiwara Y, Ohta K, Oshitani N, Higuchi K, Arakawa T. Exogenous administration of mesenchymal stem cells ameliorates dextran sulfate sodium-induced colitis via anti-inflammatory action in damaged tissue in rats. Life Sci. 2008;83:771–779. doi: 10.1016/j.lfs.2008.09.016. [DOI] [PubMed] [Google Scholar]
- 61.Valcz G, Krenács T, Sipos F, Leiszter K, Tóth K, Balogh Z, Csizmadia A, Műzes G, Molnár B, Tulassay Z. The Role of the Bone Marrow Derived Mesenchymal Stem Cells in Colonic Epithelial Regeneration. Pathol Oncol Res. 2011;17:11–16. doi: 10.1007/s12253-010-9262-x. [DOI] [PubMed] [Google Scholar]
- 62.De Wever O, Demetter P, Mareel M, Bracke M. Stromal myofibroblasts are drivers of invasive cancer growth. Int J Cancer. 2008;123:2229–2238. doi: 10.1002/ijc.23925. [DOI] [PubMed] [Google Scholar]
- 63.Stenson WF. Toll-like receptors and intestinal epithelial repair. Curr Opin Gastroenterol. 2008;24:103–107. doi: 10.1097/MOG.0b013e3282f44a2a. [DOI] [PubMed] [Google Scholar]
- 64.Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F, Edberg S, Medzhitov R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell. 2004;118:229–241. doi: 10.1016/j.cell.2004.07.002. [DOI] [PubMed] [Google Scholar]
- 65.Li M, Carpio DF, Zheng Y, Bruzzo P, Singh V, Ouaaz F, Medzhitov RM, Beg AA. An essential role of the NF-kappa B/Toll-like receptor pathway in induction of inflammatory and tissue-repair gene expression by necrotic cells. J Immunol. 2001;166:7128–7135. doi: 10.4049/jimmunol.166.12.7128. [DOI] [PubMed] [Google Scholar]
- 66.Beg AA. Endogenous ligands of Toll-like receptors: implications for regulating inflammatory and immune responses. Trends Immunol. 2002;23:509–512. doi: 10.1016/s1471-4906(02)02317-7. [DOI] [PubMed] [Google Scholar]
- 67.Taylor KR, Trowbridge JM, Rudisill JA, Termeer CC, Simon JC, Gallo RL. Hyaluronan fragments stimulate endothelial recognition of injury through TLR4. J Biol Chem. 2004;279:17079–17084. doi: 10.1074/jbc.M310859200. [DOI] [PubMed] [Google Scholar]
- 68.Majors AK, Austin RC, de la Motte CA, Pyeritz RE, Hascall VC, Kessler SP, Sen G, Strong SA. Endoplasmic reticulum stress induces hyaluronan deposition and leukocyte adhesion. J Biol Chem. 2003;278:47223–47231. doi: 10.1074/jbc.M304871200. [DOI] [PubMed] [Google Scholar]
- 69.Tang XY, Zhu YQ, Wei B, Wang H. Expression and functional research of TLR4 in human colon carcinoma. Am J Med Sci. 2010;339:319–326. doi: 10.1097/MAJ.0b013e3181cef1b7. [DOI] [PubMed] [Google Scholar]
- 70.Zhou B, Zhou H, Ling S, Guo D, Yan Y, Zhou F, Wu Y. Activation of PAR2 or/and TLR4 promotes SW620 cell proliferation and migration via phosphorylation of ERK1/2. Oncol Rep. 2011;25:503–511. doi: 10.3892/or.2010.1077. [DOI] [PubMed] [Google Scholar]
- 71.Gasser M, Grimm M, Kim M, Rosenwald A, Germer CT, Waaga-Gasser A. Toll-like receptor (TLR) 7 and TLR8 expression in CD133+ cells in colorectal cancer and role for inflammation-induced TLRs in tumorigenesis and tumor progression. J Clin Oncol. 2010;28:15s. doi: 10.1016/j.ejca.2010.07.017. [DOI] [PubMed] [Google Scholar]
- 72.Onishi S, Miyata H, Inamoto T, Qi WM, Yamamoto K, Yokoyama T, Warita K, Hoshi N, Kitagawa H. Immunohistochemical study on the delayed progression of epithelial apoptosis in follicle-associated epithelium of rat Peyer’s patch. J Vet Med Sci. 2007;69:1123–1129. doi: 10.1292/jvms.69.1123. [DOI] [PubMed] [Google Scholar]
- 73.Renes IB, Verburg M, Bulsing NP, Ferdinandusse S, Büller HA, Dekker J, Einerhand AW. Protection of the Peyer’s patch-associated crypt and villus epithelium against methotrexate-induced damage is based on its distinct regulation of proliferation. J Pathol. 2002;198:60–68. doi: 10.1002/path.1183. [DOI] [PubMed] [Google Scholar]
- 74.Cadigan KM, Nusse R. Wnt signaling: a common theme in animal development. Genes Dev. 1997;11:3286–3305. doi: 10.1101/gad.11.24.3286. [DOI] [PubMed] [Google Scholar]
- 75.DasGupta R, Fuchs E. Multiple roles for activated LEF/TCF transcription complexes during hair follicle development and differentiation. Development. 1999;126:4557–4568. doi: 10.1242/dev.126.20.4557. [DOI] [PubMed] [Google Scholar]
- 76.Korinek V, Barker N, Moerer P, van Donselaar E, Huls G, Peters PJ, Clevers H. Depletion of epithelial stem-cell compartments in the small intestine of mice lacking Tcf-4. Nat Genet. 1998;19:379–383. doi: 10.1038/1270. [DOI] [PubMed] [Google Scholar]
- 77.Haegebarth A, Clevers H. Wnt signaling, lgr5, and stem cells in the intestine and skin. Am J Pathol. 2009;174:715–721. doi: 10.2353/ajpath.2009.080758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ouji Y, Yoshikawa M, Shiroi A, Ishizaka S. Wnt-10b secreted from lymphocytes promotes differentiation of skin epithelial cells. Biochem Biophys Res Commun. 2006;342:1063–1069. doi: 10.1016/j.bbrc.2006.02.028. [DOI] [PubMed] [Google Scholar]
- 79.Malhotra S, Kincade PW. Wnt-related molecules and signaling pathway equilibrium in hematopoiesis. Cell Stem Cell. 2009;4:27–36. doi: 10.1016/j.stem.2008.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.O’Leary AD, Sweeney EC. Lymphoglandular complexes of the colon: structure and distribution. Histopathology. 1986;10:267–283. doi: 10.1111/j.1365-2559.1986.tb02481.x. [DOI] [PubMed] [Google Scholar]
- 81.Michael-Robinson JM, Biemer-Hüttmann A, Purdie DM, Walsh MD, Simms LA, Biden KG, Young JP, Leggett BA, Jass JR, Radford-Smith GL. Tumour infiltrating lymphocytes and apoptosis are independent features in colorectal cancer stratified according to microsatellite instability status. Gut. 2001;48:360–366. doi: 10.1136/gut.48.3.360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Nozoe T, Korenaga D, Futatsugi M, Saeki H, Ohga T, Sugimachi K. Cyclin A expression in superficial squamous cell carcinoma of the esophagus and coexisting infiltrated lymphocyte follicle. Cancer Lett. 2002;188:221–229. doi: 10.1016/s0304-3835(02)00434-2. [DOI] [PubMed] [Google Scholar]
- 83.Gutfeld O, Prus D, Ackerman Z, Dishon S, Linke RP, Levin M, Urieli-Shoval S. Expression of serum amyloid A, in normal, dysplastic, and neoplastic human colonic mucosa: implication for a role in colonic tumorigenesis. J Histochem Cytochem. 2006;54:63–73. doi: 10.1369/jhc.5A6645.2005. [DOI] [PubMed] [Google Scholar]
- 84.Thiery JP. Epithelial-mesenchymal transitions in development and pathologies. Curr Opin Cell Biol. 2003;15:740–746. doi: 10.1016/j.ceb.2003.10.006. [DOI] [PubMed] [Google Scholar]
- 85.Rubio D, Garcia S, De la Cueva T, Paz MF, Lloyd AC, Bernad A, Garcia-Castro J. Human mesenchymal stem cell transformation is associated with a mesenchymal-epithelial transition. Exp Cell Res. 2008;314:691–698. doi: 10.1016/j.yexcr.2007.11.017. [DOI] [PubMed] [Google Scholar]
- 86.Prindull G, Zipori D. Environmental guidance of normal and tumor cell plasticity: epithelial mesenchymal transitions as a paradigm. Blood. 2004;103:2892–2899. doi: 10.1182/blood-2003-08-2807. [DOI] [PubMed] [Google Scholar]
- 87.Kang Y, Massagué J. Epithelial-mesenchymal transitions: twist in development and metastasis. Cell. 2004;118:277–279. doi: 10.1016/j.cell.2004.07.011. [DOI] [PubMed] [Google Scholar]
- 88.Spees JL, Olson SD, Ylostalo J, Lynch PJ, Smith J, Perry A, Peister A, Wang MY, Prockop DJ. Differentiation, cell fusion, and nuclear fusion during ex vivo repair of epithelium by human adult stem cells from bone marrow stroma. Proc Natl Acad Sci USA. 2003;100:2397–2402. doi: 10.1073/pnas.0437997100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Van Arnam JS, Herzog E, Grove J, Bruscia E, Ziegler E, Swenson S, Krause DS. Engraftment of bone marrow-derived epithelial cells. Stem Cell Rev. 2005;1:21–27. doi: 10.1385/SCR:1:1:021. [DOI] [PubMed] [Google Scholar]
- 90.Sheng W, Wang G, La Pierre DP, Wen J, Deng Z, Wong CK, Lee DY, Yang BB. Versican mediates mesenchymal-epithelial transition. Mol Biol Cell. 2006;17:2009–2020. doi: 10.1091/mbc.E05-10-0951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Hirose J, Kawashima H, Yoshie O, Tashiro K, Miyasaka M. Versican interacts with chemokines and modulates cellular responses. J Biol Chem. 2001;276:5228–5234. doi: 10.1074/jbc.M007542200. [DOI] [PubMed] [Google Scholar]
- 92.Mesker WE, Liefers GJ, Junggeburt JM, van Pelt GW, Alberici P, Kuppen PJ, Miranda NF, van Leeuwen KA, Morreau H, Szuhai K, et al. Presence of a high amount of stroma and downregulation of SMAD4 predict for worse survival for stage I-II colon cancer patients. Cell Oncol. 2009;31:169–178. doi: 10.3233/CLO-2009-0478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Pohl M, Radacz Y, Pawlik N, Schoeneck A, Baldus SE, Munding J, Schmiegel W, Schwarte-Waldhoff I, Reinacher-Schick A. SMAD4 mediates mesenchymal-epithelial reversion in SW480 colon carcinoma cells. Anticancer Res. 2010;30:2603–2613. [PubMed] [Google Scholar]