

Abstract
The beneficial effects of mindful awareness and mindfulness meditation training on physical and psychological health are thought to be mediated in part through changes in underlying brain processes. Functional connectivity MRI (fcMRI) allows identification of functional networks in the brain. It has been used to examine state-dependent activity and is well-suited for studying states such as meditation. We applied fcMRI to determine if Mindfulness-Based Stress Reduction (MBSR) training is effective in altering intrinsic connectivity networks (ICNs). Healthy women were randomly assigned to participate in an 8 week Mindfulness-Based Stress Reduction (MBSR) training course or an 8 week waiting period. After 8 weeks, fMRI data (1.5 T) was acquired while subjects rested with eyes closed, with the instruction to pay attention to the sounds of the scanner environment. Group independent component analysis was performed to investigate training-related changes in functional connectivity. Significant MBSR-related differences in functional connectivity were found mainly in auditory/salience and medial visual networks. Relative to findings in the control group, MBSR subjects showed (1) increased functional connectivity within auditory and visual networks, (2) increased functional connectivity between auditory cortex and areas associated with attentional and self-referential processes, (3) greater anticorrelation between auditory and visual cortex, and (4) greater anticorrelation between visual cortex and areas associated with attentional and self-referential processes. These findings suggest that 8 weeks of mindfulness meditation training alters intrinsic functional connectivity in ways that may reflect a more consistent attentional focus, enhanced sensory processing, and reflective awareness of sensory experience.
Keywords: mindfulness, meditation, functional connectivity, resting state fMRI, auditory, attention
1. Introduction
Mindfulness meditation is an attention-training technique that is receiving increased research and public interest in the West (Ludwig and Kabat-Zinn, 2008; Lutz et al., 2008b). It consists of training practitioners in how to be mindful, an attentional stance of openly and non-judgmentally observing one’s moment-by-moment experiences. A growing body of evidence suggests that mindfulness meditation training may reduce stress and improve some aspects of physical and psychological health in patient populations (Brown et al., 2007; Chiesa and Serretti, 2009, 2010; Creswell et al., 2009). Moreover, new neuroimaging studies suggest that mindfulness meditation training, such as that offered in the standardized and manualized 8-week Mindfulness-Based Stress Reduction (MBSR) intervention (Kabat-Zinn, 1990), can alter functional neural responding to affective tasks in healthy individuals (Farb et al., 2010).
A major component of mindfulness meditation training is focusing and refocusing awareness on present moment internal and external experience (e.g. breathing, thoughts, ambient sounds, interoceptive sensations) as opposed to mind wandering or active problem solving. MBSR training includes breath awareness, body awareness, attention to the transient nature of sensory experience, and shifting attention across sensory modalities. It is therefore hypothesized that meditation practice should lead to altered functioning of brain networks involved in attention and sensitivity to internal and external sensations and emotions. To date only a small number of studies examined brain mechanisms involved in mindfulness meditation using functional brain imaging. Although the results across these small studies are not completely consistent they do suggest mindfulness meditation leads to increased activations in frontal brain regions involved in attention (Farb et al., 2007; Goldin and Gross, 2010; Holzel et al., 2007) and increased activations in temporal regions during processing of sounds (Lutz et al., 2008a). In addition, mindfulness meditation experience is associated with increased cortical thickness in anterior insula (involved in interoception), frontal cortex (Brodmann’s area 9/10; involved in the integration of emotion and cognition) and sensory cortices (Lazar et al., 2005). Thus there is evidence that mindfulness meditation is associated with changes in attention and perceptual processing circuits, perhaps reflecting the emphasis on conscious direction and redirection of attention to present moment experience and increased awareness of sensory stimuli.
The examination of intrinsic connectivity networks (ICNs), sometimes referred to as resting state networks (Seeley et al., 2007), provides information about the integrity and organization of major functional systems of the brain. Even in the absence of explicit, phasic changes induced by a sensory or cognitive task, intrinsic brain activity is characterized by slow fluctuations in the BOLD signal (<0.1 Hz) which exhibit a high degree of spatial structure reminiscent of task-related activation maps (Damoiseaux et al., 2006; Smith et al., 2009). Coherent functional networks most consistently identified by fcMRI include: the default mode network (DMN) associated with episodic memory and self-referential processing; the executive control network associated with salience processing; lateralized fronto-parietal networks associated with working memory and attentional processes; and sensory-related networks (medial visual, lateral visual, auditory, sensorimotor) (Damoiseaux et al., 2006; Smith et al., 2009).
Intrinsic connectivity fMRI has been used to characterize neural networks in healthy and patient populations (Buckner et al., 2008; Greicius, 2008), and it may be also particularly relevant for the study of changes in functional connectivity that are hypothesized to result from regular practice of mindfulness meditation. Intrinsic connectivity reflects both anatomically constrained spontaneous fluctuations and state-dependent activity (Gopinath et al., 2010) and is well-suited for studying states such as meditation. Intrinsic activity has been shown to be modulated in terms of connectivity, amplitude or spectral power distribution by various conditions such as rest with eyes open or closed, sleep, transcutaneous electrical nerve stimulation, muscle fatigue and by pharmological manipulations (Bianciardi et al., 2009; Esposito et al., 2010; Gopinath et al., 2010; Horovitz et al., 2008; Peltier et al., 2005; Rack-Gomer et al., 2009; Yan et al., 2009). Also, intrinsic connectivity changes have been found following intense training on a difficult visual discrimination task (Lewis et al., 2009). Thus, fcMRI provides a tool for examining training-related plasticity and neural mechanisms underlying states such as meditation. The aim of the present study was to determine if Mindfulness-Based Stress Reduction (MBSR) training alters the functional connectivity of intrinsic connectivity networks during mindful awareness of auditory sensation. Given that the formal practice of MBSR highlights attention to sensory experience, we hypothesized that ICNs associated with attention and sensory processing would show training-related changes.
2. Materials and methods
2.1 Subjects
32 healthy, meditation-naïve women (ages 21–55; mean = 34.1; standard deviation = 9.8) were recruited by advertisement. Subjects were screened via medical exam for absence of significant health or psychiatric conditions. All study protocols were performed after approval by UCLA and informed consent was obtained from all subjects. Subjects were compensated for participation by the provision of the MBSR training program free of charge. One additional subject completed the MBSR training; however, due to scanner technical difficulties, her fMRI data was excluded from all analyses.
2.2 Behavioral Measures
To ensure that the MBSR program was indeed increasing mindfulness (compared to wait-list controls), a 15-item measure of trait mindfulness was administered at baseline and after the 8-week treatment period (Brown and Ryan, 2003). Trait mindfulness, defined as an open or receptive awareness to present moment experience, was assessed by the Mindfulness Attention Awareness Scale (MAAS) (Brown and Ryan, 2003). The MAAS is a 15-item self-report measure that asks participants to rate the frequency of their mindful states in day-to-day life. In addition, subjects completed the State-Trait Anxiety Inventory (STAI) as a measure of state and trait anxiety (Spielberger et al., 1970).
2.3 Procedures
Subjects were randomly assigned to participate in an 8 week Mindfulness-Based Stress Reduction (MBSR) training course (n=17) or to an 8 week waiting period (n=15). The two groups did not differ in age (t(30)=1.48, p>.05) or state anxiety (t(30)=.647, p>.05) and trait anxiety (t(30)=.05, p>.05) as measured by the State Train Anxiety Inventory (Table 1). Subjects in the MBSR group immediately participated in the 8 week MBSR training course. Subjects in the wait-list control group had an 8 week waiting period before receiving MBSR training. The wait-list condition was intended to control for subjects’ level of interest in meditation training and general motivation to participate in the study, while keeping their current meditation experience at a baseline (zero) level. After the 8 week training/waiting period, subjects completed intrinsic connectivity fMRI scans. Subjects in the wait-list group were then provided MBSR training. Subjects completed questionnaires at the randomization visit and the fMRI visit.
Table 1.
Group means and standard deviations for age and STAI measures at the randomization visit
| Age | STAI-state | STAI-trait | |
|---|---|---|---|
| MBSR | 31.7 (9.6) | 37.1(8.4) | 38.4(9.3) |
| Controls | 36.7(9.6) | 35.3(6.9) | 38.5(11.0) |
2.4 MBSR Training
MBSR training consisted of eight weekly 150-minute group sessions, a day-long retreat in the sixth/seventh week, and daily home practice. During each group session, an instructor lead subjects in different guided meditations, awareness exercises, mindful movement, and group discussions, with the intent of fostering mindful awareness of how one responds to stress. The day-long retreat (7 hours) focused on integrating and elaborating on exercises learned during the course. Participants were expected to do 30 minutes of daily home practice six days a week during the course with guided meditations on tape and to keep a record to allow assessment of level of compliance to this aspect of the training. The training was conducted by two experienced MBSR instructors (http://www.insightla.org/).
2.5 fMRI Acquisition
MRI scanning was performed using a 1.5T MRI scanner (Siemens Sonata; Siemens, Erlangen, Germany). A high resolution structural image was acquired from each subject with a magnetization-prepared rapid gradient-echo (MP-RAGE) sequence, repetition time (TR) = 2300 ms, echo time (TE) = 2.85 ms, slice thickness = 1 mm, 160 slices, 128 × 128 voxel matrices, and 13 mm voxel size. Subjects completed two mindfulness state scans in which functional blood oxygen-level dependent (BOLD) images were acquired for 5.5 minutes each in 38-slice whole brain volumes, slice thickness = 3 mm, TR = 3000 ms, TE = 28 ms, flip angle = 90°.
Prior to entering the scanner, subjects completed the MAAS questionnaire. Subjects completed two 5.5 minute mindfulness state scans; one with ambient scanner noise only and one with modulated white noise presented through headphones in addition to the ambient scanner noise, counterbalanced for order. The modulated white noise scan was originally included to potentially aid in the development of a novel analytical technique but this was not attempted in the current analysis. During the mindfulness state scans all subjects received the following instructions: “During this baseline scan of about 5 min, we would like you to again stay as still as possible and be mindfully aware of your surroundings. Please keep your eyes closed during this procedure. Continue to be mindfully aware of whatever you notice in your surroundings and your own sensations. Mindful awareness means that you pay attention to your present moment experience, in this case the changing sounds of the scanner/changing background sounds played through the headphones, and to bring interest and curiosity to how you are responding to them.” Following the mindfulness state scans, subjects completed additional fMRI scans involving a pain-related task; these data are not included in the current report, and will be reported elsewhere.
2.6 fMRI Preprocessing
Using SPM5 software (Welcome Department of Cognitive Neurology, London, UK), data were slice-time and motion corrected, spatially normalized to the MNI template using the structural imgaes, and spatially smoothed with a 5 mm3 Gaussian kernel. The first two volumes were discarded to allow for stabilization of the magnetic field. In addition, the last 8 scans were excluded from analyses as movement in the scanner increased towards the end of the acquisition for several subjects. Therefore, fMRI analyses were based on two 5 minute runs (100 timepoints each) of state fMRI data.
2.7 Statistical Analyses
Group independent component analysis (gICA) was performed to investigate training-related changes in functional connectivity among ICNs during mindful rest. All 32 subjects were entered into the gICA implemented in GIFT (http://icatb.sourceforge.com). The number of independent components was estimated to be 18 using the MDL criteria (Li et al., 2006). Eighteen independent components were extracted by ICA decomposition using the infomax algorithm (Bell and Sejnowski, 1995). Multiple runs (20 iterations) of ICA were performed using ICASSO to determine the reliability of the ICA algorithm (Himberg et al., 2004). ICASSO outputs a final set of independent components representing the centroid of the results from the multiple iterations. Each independent component consists of a spatial map and an associated timecourse. The component containing the default mode network (DMN) was identified by spatial correlation with a weighted template provided in GIFT (Franco et al., 2009). Additional components were selected that displayed spatial correlation with the SPM grey matter template greater than 0.15 and the majority of power spectral distribution in the lower frequencies (<.08 Hz). In total, eight components were selected for second-level analyses (Figure 1). Individual subject maps were back-reconstructed. These maps contained z-score values that represented the degree of correlation between the voxel signal and the timecourse of the component (Greicius et al., 2007).
Figure 1.

Intrinsic connectivity networks identified by ICA: a) auditory/salience network; b) medial visual network; c) lateral visual network; d) sensorimotor network; e) default mode network; f) executive control network; g & h) frontal parietal networks.
Subject maps were entered into SPM random-effects analyses. For each component, single group maps were created by performing one-way repeated measure (Ambient Noise Only; Modulated White Noise) ANOVAs. To detect possible regional differences in ICN functional connectivity, a Group (MBSR; controls) × Scan (Ambient Noise Only; White Noise) Mixed Model ANOVA was performed for each component with an explicit mask created from the single group maps (|T-value|≥3.0 on one or both single group maps). Results were thresholded at a corrected cluster significance level of p=0.05. No significant differences were found between the white noise and ambient noise scans on any component (i.e. no main effect for scan or interaction); thus, only group effects are reported. Behavioral data was analyzed by mixed linear model analysis using PASW 17 (Chicago, IL). All functional data were overlaid on the MNI template available in MRIcro (http://www.sph.s.c.edu/comd/rorden/mricro.html) for presentation purposes.
Additional post hoc, seed-based correlation analyses were performed on significant clusters reported in 3.3.1 using the CONN fMRI connectivity toolbox (v12.1, http://web.mit.edu/swg/software.htm). The toolbox removes confounding effects related to white matter/CSF signal as well as motion parameters (six dimensions with first order derivative). Functional data was bandpass filtered (0.01 to 0.10 Hz). Fisher transformed maps of the bivariate correlation between the seed ROI time-course and all other voxels were created and contrasted at the group level in SPM5 using an explicit mask from the identified ICN. Results were thresholded at a height of p<.001 uncorrected and extent > 10 voxels.
3. Results
3.1 MBSR Training Compliance
All MBSR subjects completed at least six of the eight classes. Meditation diaries were not received from 2 MBSR subjects. Of the remaining 15 MBSR subjects, 10 attended all eight classes; three attended seven classes; two attended six classes and 10 attended the daylong retreat. Diary data indicated that MBSR subjects completed an average of 2716.5 minutes of meditation during the course (± 184.6 standard error; range: 1520–3925 minutes).
3.2 MAAS
MAAS scores for three of the wait-list control subjects were not collected at the fMRI visit due to an oversight. A mixed linear model demonstrated a significant interaction between Group (MBSR; Control) and Visit (Randomization; fMRI) (F(1,30)=4.982, p=.033). At the randomization visit, subjects assigned to the MBSR group did not differ from wait-list controls in trait mindfulness as assessed by the MAAS (p=.718). However, at the fMRI visit, the MBSR subjects had significantly greater MAAS scores compared to wait list controls (p=.009). Estimated means and standard errors are presented in Table 2. These results show the effectiveness of MBSR training in increasing mindfulness while the wait-list group maintained baseline mindfulness.
Table 2.
Estimated means and standard errors for MAAS scores
| Randomization Visit | fMRI Visit | |
|---|---|---|
| MBSR | 3.878 (.203) | 4.686(.171) |
| Controls | 3.987(.216) | 3.931(.204) |
3.3 Intrinsic Connectivity Networks
Of the eight selected components of interest (Figure 1), six displayed significant MBSR-related changes in the strength of regional functional connectivity (Table 3). The components with the most widespread differences (in terms of spatial extent) included the auditory/salience ICN and the medial visual ICN.
Table 3.
Significant MBSR-related changes in functional connectivity for each Intrinsic Connectivity Network
| ICN | Region | Hemisphere | Cluster Size | MNI coordinates | TMBSR | TWLC | Tdiff | pdiff |
|---|---|---|---|---|---|---|---|---|
| Auditory/Salience | BA 9/32 | right | 60 | 12 42 18 | 3.33 | −3.62 | 4.73 | .008 |
| OP | left | 42 | −50 −20 16 | 9.29 | 4.98 | 4.82 | .045 | |
| BA 18 | bilateral | 458 | 0 −74 18 | −3.01 | 4.27 | 5.12 | <.001 | |
| BA 19 | left | 59 | −24 −96 20 | −6.29 | 1.19 | 5.22 | .009 | |
| Medial Visual | BA 30/17 | left | 395 | −20 −68 4 | 15.73 | 5.06 | 5.89 | <.001 |
| BA 30 | right | 158 | 20 −72 12 | 9.19 | 4.10 | 5.42 | <.001 | |
| BA 24/32 | right | 43 | 6 38 10 | −10.01 | −1.23 | 4.76 | .043 | |
| Lateral Visual | BA 23 | right | 59 | 2 −26 32 | 3.83 | −3.21 | 5.26 | .018 |
| BA 4 | right | 88 | 10 −46 70 | −1.77 | 5.99 | 5.20 | .002 | |
| BA 5 | right | 47 | 6 −42 56 | −4.57 | 2.35 | 4.44 | .05 | |
| Sensori-motor | BA 31 | bilateral | 47 | 0 −74 22 | 4.49 | −2.26 | 4.46 | .025 |
| BA 30 | right | 63 | 20 −50 6 | 4.59 | −2.20 | 4.98 | .006 | |
| Executive Control | BA 40 | left | 76 | −50 −50 32 | −2.76 | −4.09 | 4.35 | .003 |
Note: OP = parietal operculum; TMBSR = T-value on MBSR single group map; TWLC = T-value on wait list control single group map; Tdiff and pdiff = T-value and corrected p-value for the two-group comparison
3.3.1 Auditory/salience ICN
The identified auditory/salience ICN (Figure 1a) included primary auditory (BA41/42), superior temporal gyrus (BA22/38), and posterior insula, (p<.05, corrected), similar to an ICN identified in a previous study as corresponding to task co-activation maps in the auditory domain (Smith et al., 2009). In addition, the ICN included regions of the anterior cingulate (BA24/32) and lateral frontal cortex (BA46), which are typically associated with salience and executive control networks (Seeley et al., 2007). Therefore, the ICN was labeled as “auditory/salience”. It is unclear why the components split differently in our sample, perhaps the instructions that brought attention to auditory input altered the covariance structure somewhat. Chang & Glover (2010) demonstrated high variability during resting scans in the coherence of salience/executive control regions with a DMN seed in the posterior cingulate. In addition, visual inspection of the supporting information from Smith et al (2009) shows sACC to be present in multiple components, either in the resting state or in the BrainMap-derived components, including the executive control, right frontoparietal, somatosensory, and cerebellar. We might therefore speculate that the connectivity of salience/executive control regions with other networks shows variability and sensitivity to state.
Results from the single group analyses and two-group comparison are presented in Figure 2. MBSR subjects, compared to wait list controls demonstrated significantly increased functional connectivity with the auditory/salience ICN in two regions: (1) right dorsomedial prefrontal cortex (dmPFC) and (2); the left medial parietal operculum (extending into posterior insula; Table 3). The parietal operculum cluster may be localized to OP 2 or OP 3 (Eickhoff et al., 2006). The left parietal operculum had a positive correlation with auditory/salience ICN intrinsic activity in both groups, but was significantly more correlated in the MBSR subjects. The right dmPFC had a negative correlation with auditory/salience ICN intrinsic activity in wait list controls and a positive correlation in MBSR subjects (see Table 3). Although intrinsic connectivity networks are largely nonoverlapping, some areas of overlap can be found (Seeley et al., 2007). The right dmPFC cluster overlapped with the DMN and demonstrated a significant positive association with the DMN in both subject groups.
Figure 2.

Single group maps of the auditory/salience ICN for MBSR subjects (left panel) and wait list controls subjects (middle panel) are presented. In the right panel, significant group differences are shown; red indicates areas of significantly increased functional connectivity while blue indicates areas of greater anticorrelation with the auditory/salience ICN in MBSR subjects compared to wait list controls.
In addition, visual regions (bilateral BA 18/19) demonstrated significantly decreased functional connectivity with the auditory/salience ICN in MBSR subjects relative to controls. These visual regions overlapped the medial visual ICN and had a negative correlation with auditory/salience ICN intrinsic activity in MBSR subjects and a positive/zero correlation in wait list controls (see Table 3). Thus, significantly greater anticorrelation between medial visual cortex (BA 18/19) and the auditory/salience ICN was seen in the MBSR subjects compared to controls.
Ad hoc analyses were performed to aid interpretation of the results. The four regions displaying group differences in functional connectivity with the auditory/salience ICN (the left BA 19, bilateral BA 18, right dmPFC, and left OP) were used as seed ROIs in seed-based correlation analyses. The primary purpose of the ad hoc analyses was to determine which regions within the auditory/salience ICN showed the greatest change in functional connectivity with each of the seed ROIs. Although the visual regions (BA 18, BA 19) showed significantly greater anti-correlation with the auditory/salience ICN in MBSR subjects, the difference was most robust in auditory region left BA 22 for both BA 18 and BA 19 seed ROIs (coordinates: −64 −38 8; −62 −46 18, respectively), right inferior parietal BA 40 for the BA 19 seed ROI (coordinates: 60 −36 28) and right ACC (BA 24) for the BA 18 seed ROI (coordinates: 6 36 10; located within the identified DMN). Although OP and dmPFC showed significantly greater correlation with the auditory/salience ICN in MBSR subjects, the difference was most robust in right BA 22 and dorsal ACC (dACC; BA 32) for the OP seed ROI (coordinates: 58 8 −6; 8 12 48, respectively); and right BA 22/39 and left dACC (BA 24) for the dmPFC seed ROI (coordinates: 46 −52 8; −4 2 40, respectively). The dACC clusters did not overlap with the DMN.
3.3.2 Medial Visual ICN
The identified medial visual ICN (Figure 1b) included visual cortex (BA 17/18/19), retrosplenial cortex (BA 30; p<.05, corrected) and was well-matched to an ICN identified in a previous study as corresponding to task co-activation maps in the visual domain (Smith et al., 2009). Results from the single group and two-group comparison are presented in Figure 3. MBSR subjects, compared to wait list controls demonstrated significantly increased functional connectivity with the medial visual ICN in bilateral calcarine retrosplenial cortex (BA 30; Table 3). The retrosplenial cortex demonstrated positive correlation with medial visual ICN intrinsic activity in both groups, but was significantly more correlated in the MBSR subjects. This portion of the retrosplenial cortex was more posterior to retrosplenial cortex in the DMN and did not overlap with the DMN (p>.05). In addition, the right supragenual anterior cingulate cortex (sACC; BA 24/32) demonstrated significantly decreased functional connectivity with the medial visual ICN in MBSR subjects compared to controls. The right sACC was negatively correlated with medial visual ICN intrinsic activity in MBSR subjects and was not correlated in control subjects. Thus, significantly greater anticorrelation between the right sACC and medial visual ICN intrinsic activity was seen in the MBSR subjects compared to controls. This portion of the ACC was positively correlated with DMN intrinsic activity in both groups (p<.05, corrected).
Figure 3.

Single group maps of the medial visual ICN for MBSR subjects (left panel) and wait list controls subjects (middle panel) are presented. In the right panel, significant group differences are shown; red indicates areas of significantly increased functional connectivity while blue indicates areas of greater anticorrelation with the medial visual ICN in MBSR subjects compared to wait list controls.
An ad hoc analysis was performed to evaluate group differences in the overall level of within-network functional connectivity of the medial visual ICN. The average ICA score within a spatial template containing the positively weighted voxels common to the MBSR and control ICN maps was computed across runs using Marsbar (Brett et al., 2002). A two-sample t-test demonstrated a significant increase in the overall network functional connectivity in the MBSR subjects compared to wait list controls (t(30)=2.298, p=.029).
3.3.3 Additional ICNs
Table 3 includes information about significant differences in functional connectivity between MBSR trained subjects and wait list controls for executive control (Figure 1f), sensorimotor (Figure 1d) and lateral visual networks (Figure 1c). These differences are not as extensive as those seen in the auditory/salience and medial visual networks. The default mode network (Figure 1e) and two frontoparietal networks (Figure 1g, h) did not show any significant training-related differences in functional connectivity.
4. Discussion
The results indicate that a course of MBSR training in healthy subjects impacts regional functional connectivity within auditory-related and medial visual intrinsic connectivity networks during instructed mindful awareness to auditory stimuli. MBSR subjects, relative to wait list controls, displayed increased functional connectivity of the parietal operculum and the dmPFC (BA9/32) in the auditory/salience ICN and increased functional connectivity of retrosplenial cortex in the medial visual ICN. In addition, MBSR subjects displayed stronger differentiation (greater anticorrelation) between the auditory/salience ICN and portions of medial visual ICN and between the medial visual ICN and the sACC. The results are discussed below in terms of four main hypotheses.
4.1 Increased connectivity among auditory and salience/control regions in MBSR subjects during mindful awareness of sounds
The parietal operculum was functionally correlated with an auditory/salience network in both MBSR and control groups. However, MBSR subjects demonstrated significantly increased functional connectivity of this region with the auditory/salience network, especially with BA 22 (auditory association cortex) and dorsal ACC (involved in salience). The parietal operculum contains auditory-related cortex and plays a role in auditory function (Celesia, 1976; Galaburda and Sanides, 1980). The parietal operculum is strongly activated by nonvocal sounds and may mediate processing of fine-grained, temporal acoustic features (Binder et al., 1996; Meyer et al., 2005). Our results demonstrate that MBSR-trained subjects during mindful awareness of sounds have greater synergy/positive coherence between a region involved in fine-grained perceptual processing of auditory information and other auditory-related regions as well as salience/control regions.
Although ICNs are mostly distinct, they are not completely independent (Seeley et al., 2007). Nodes that are present in two networks may be involved in information transfer, facilitate switching between the networks or underlie another key interaction process (Seeley et al., 2007). The dmPFC was functionally correlated with the DMN network in both MBSR and control groups. However, MBSR subjects demonstrated significantly increased functional connectivity of this anterior DMN region with the auditory/salience network, especially with BA 22/39 (associated with auditory processing) and dorsal ACC (involved in salience). Anterior cortical midline structures such as dmPFC are considered crucial in transforming simple sensory processing into more complex self-referential processing (Northoff et al., 2006). Dorsomedial PFC is involved in the process of attending to mental states (Walter et al., 2009) and has been previously implicated in studies of mindfulness. Trait mindfulness has been associated with increased dmPFC activation during reappraisal of negative emotion (Modinos et al., 2010) and during affect labeling (Creswell et al., 2007). The increased functional connectivity between dmPFC and auditory/salience ICN suggests greater synergy/positive coherence between self-referential, attention, and auditory sensory processing and may underlie a greater attention and reflective awareness of auditory experience in MBSR trained subjects.
4.2 Increased connectivity of the medial visual network in MBSR subjects during mindful awareness of sounds
Retrosplenial cortex (BA 30) was functionally correlated with a visual-related network in both MBSR and control groups, yet MBSR subjects demonstrated significantly increased functional connectivity of this region. Spontaneous BOLD oscillations of the retrosplenial cortex are sensitive to whether the subject is resting with eyes closed or eyes open, with increased oscillations and functional connectivity during eyes closed (McAvoy et al., 2008). The retrosplenial cortex is activated by scene viewing and imagery and is involved in placing a scene within a broader spatial environment by integrating egocentric spatial information into an allocentric environmental representation (Burgess, 2008; Epstein, 2008; O’Craven and Kanwisher, 2000). Our results show that MBSR training affects the functional connectivity between the retrosplenial cortex and the medial visual system even when the subject is mindfully meditating with eyes closed.
Spontaneous BOLD oscillations in the visual cortex are increased during light sleep and are correlated with an inverse measure of wakefulness (Horovitz et al., 2008). Also, the within network functional connectivity of the visual cortex is increased during sedation and alcohol consumption (Esposito et al., 2010) (Kiviniemi et al., 2005). Thus, arousal/vigilance level can affect the overall intrinsic functional connectivity of the visual cortex. An ad hoc analysis revealed a significant global increase in medial visual ICN functional connectivity in MBSR subjects compared to wait list controls, perhaps indicating a greater degree of relaxation in MBSR subjects.
4.3 Greater differentiation between auditory and visual intrinsic activity in MBSR subjects during mindful awareness of sounds
Reciprocal modulation/attenuation between cortical regions is a common phenomenon and has been observed between visual and auditory cortex (Laurienti et al., 2002; Northoff et al., 2004). Laurienti et al (2002) observed simultaneous activations in auditory cortex and deactivations in visual cortex during auditory stimulation and an opposite pattern during visual stimulation. MBSR subjects had significantly greater anticorrelation between visual areas and the auditory/salience ICN, especially in auditory region BA 22 (and sACC, discussed in section 4.4) suggesting greater cross-modal inhibition and less intrusion of the two forms of processing on each other.
A stronger anticorrelation between intrinsic connectivity networks has been associated with improved, less variable performance on cognitive tasks, improved perceptual learning and may reflect processing efficiency (Deco et al., 2009; Kelly et al., 2008; Lewis et al., 2009). Thus, stronger reciprocal modulation between auditory and visual areas may reflect a more consistent focus in MBSR subjects and be part of previously shown increase in attentional ability following meditation practice (Chambers et al., 2008; Jha et al., 2007; Valentine and Sweet, 1999). Alternatively, auditory information may have become increasingly dominant in the condition of poor visual stimulation (eyes closed) in the MBSR subjects compared to controls, reducing attentional resources devoted to the visual domain. Improved allocation of attentional resources both within and across sensory modalites has been demonstrated in mindfulness meditators (Slagter et al., 2007; van den Hurk et al., 2010a; van den Hurk et al., 2010b). Previous studies have shown that during eyes closed compared to eyes open, increased functional connectivity has been found in auditory, somatosensory, and visual cortex and decreased functional connectivity has been shown in default mode and attention networks (McAvoy et al., 2008; Nir et al., 2006; Yan et al., 2009). Similarly, activation studies have shown increased activity in auditory, somatosensory, lateral visual and frontopolar areas during eyes closed compared to eyes open and increased activity during eyes open compared to eyes closed in attention and ocular motor regions (Marx et al., 2003). Since eyes closed was used throughout this study it is not clear if the group differences found would also hold for eyes open.
4.4 Increased anticorrelation between visual cortex and anterior DMN in MBSR subjects during mindful awareness to sounds
In addition to increased connectivity in MBSR subjects between a region within the anterior DMN (dmPFC) and the auditory/salience network (discussed in section 4.1), a neighboring cluster (sACC) demonstrated increased anticorrelation with visual cortex (see Figure 4). This increased anticorrelation with the aDMN in MBSR subjects was reflected in both the medial visual ICN (main ICA analysis) and in the auditory/salience ICN (ad hoc seed-based correlation analysis). The same portion of the sACC was significant in both analyses. The sACC is involved in monitoring and control functions (Northoff and Bermpohl, 2004). Thus, the stronger anticorrelation may reflect greater inhibitory control in MBSR subjects necessary to maintain mindful awareness of sounds and may be related to the previously demonstrated increase in attentional ability as mentioned in section 4.3. Alternatively, reduced attentional resources may be devoted to the visual domain in conditions of poor visual stimulation (eyes closed) in the MBSR subjects compared to controls as mentioned in section 4.3.
Figure 4.
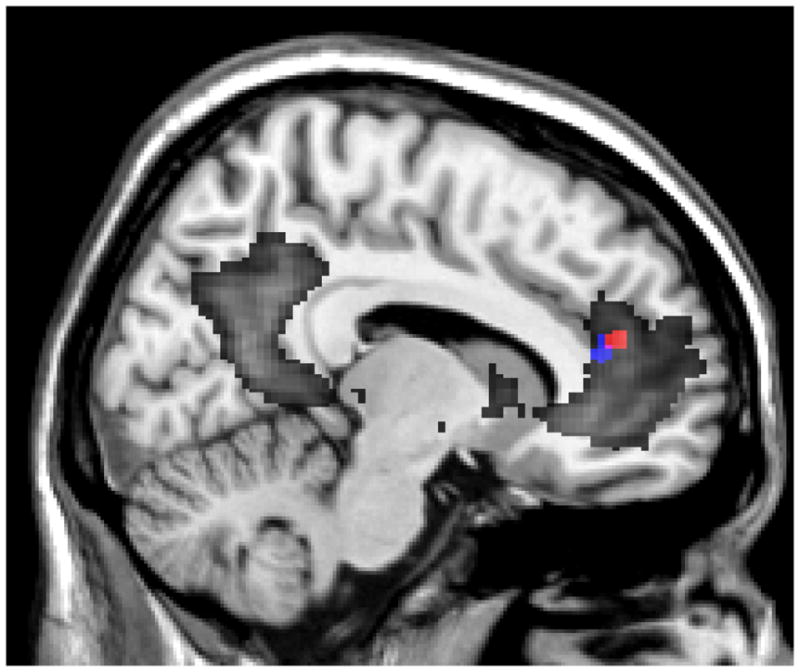
Two regions in the anterior DMN displayed altered functional connectivity with sensory-related ICNs in MBSR subjects compared to controls. The red cluster (dmPFC) had increased functional connectivity with the auditory/salience ICN. The blue cluster (sACC) had greater anticorrelation with the medial visual ICN. The DMN across all subjects is outlined in dark grey.
4.5 Comparison to training-related connectivity studies
Previous studies of training-related changes in intrinsic connectivity have demonstrated that, in general, brain networks recruited during training show robust changes in intrinsic connectivity (Albert et al., 2009; Lewis et al., 2009). MBSR training is thought to recruit attention and sensory-related networks; thus the current results demonstrating training-related changes in functional connectivity among regions associated with sensory and attentional processes are consistent with previous studies in terms of this general supposition. Similar to a previous study investigating changes in intrinsic connectivity following training on a difficult visual perceptual task (Lewis et al., 2009), the current study demonstrated training-related changes in connectivity between sensory cortex, DMN regions and regions involved in attention/salience. However, the direction of training-related changes was very different. Following training, Lewis et al (2009) found greater anticorrelation between trained regions of the visual cortex and attention areas; possibly reflecting an active decoupling of the two systems allowing for more automatic processing and less top-down control as training progressed. In the current study, MBSR subjects demonstrated increased positive connectivity between auditory and salience/attention areas; possibly reflecting less automatic, mindless processing and more top-down control, consistent with the goals of MBSR training. In addition, Lewis et al (2009) found less anticorrelation following training between untrained regions of the visual cortex and DMN regions; possibly reflecting an attenuation of a filter for unattended sensory information that became less important as training progressed. In the current study, MBSR subjects demonstrated greater anticorrelation between visual and DMN regions; possibly reflecting increased filtering of the unattended visual information, consistent with the goals of the focused attention instruction.
4.6 Limitations and future directions
In this study, subjects were specifically asked to be mindful of sounds. Open monitoring, defined as monitoring present moment experience without any particular emphasis or explicit focus, is another form of attention-focusing meditation central to MBSR. It would be useful to examine intrinsic connectivity networks without mindful instruction or without a particular emphasis (open monitoring instruction) as well as with mindful emphasis on interoception (such as awareness of breath) to see if findings are similar but more related to the emphasized domain. For example, we hypothesize that mindful awareness of breath in MBSR trained subjects may increase intrinsic functional connectivity within a frontoparietal ICN or between anterior DMN regions and a frontoparietal ICN. Futhermore, future studies should examine the relationship between MBSR-related changes in functional connectivity and task-related activations.
5. Conclusions
Intrinsic connectivity fMRI allows for an examination of the functional connectivity of basic brain processing systems during meditation states. The current findings extend the results of prior studies that showed meditation-related changes in specific brain regions active during attention and sensory processing by providing evidence that MBSR trained compared to untrained subjects, during a focused attention instruction, have increased connectivity within sensory networks and between regions associated with attentional processes and those in the attended sensory cortex. In addition they show greater differentiation between regions associated with attentional processes and the unattended sensory cortex as well as greater differentiation between attended and unattended sensory networks. If greater differentiation between networks reflects processing efficiency as previously hypothesized (Deco et al., 2009; Kelly et al., 2008; Lewis et al., 2009), our data could indicate enhanced sensory processing, better attentional resource allocation and a more consistent attentional focus in MBSR trained subjects. In addition, MBSR trained subjects displayed altered connectivity between anterior DMN regions involved in self-referential processing and sensory cortex that may indicate greater reflective awareness for sensory experience in MBSR trained subjects.
Acknowledgments
Supported by funds from the G. Oppenheimer Family Foundation; National Institutes of Health grants AT 00268, and DK64531; and the UCLA Ahmanson-Lovelace Brain Mapping Center.
Abbreviations
- MBSR
Mindfulness-Based Stress Reduction
- DMN
Default Mode Network
- ICN
Intrinsic Connectivity Network
- MAAS
Mindfulness Attention Awareness Scale
- ICA
Independent Component Analysis
- OP
parietal operculum
- dmPFC
dorsomedial prefrontal cortex
- sACC
supragenual anterior cingulate cortex
- fcMRI
functional connectivity magnetic resonance imaging
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Albert NB, Robertson EM, Miall RC. The resting human brain and motor learning. Curr Biol. 2009;19:1023–1027. doi: 10.1016/j.cub.2009.04.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell AJ, Sejnowski TJ. An information-maximization approach to blind separation and blind deconvolution. Neural Comput. 1995;7:1129–1159. doi: 10.1162/neco.1995.7.6.1129. [DOI] [PubMed] [Google Scholar]
- Bianciardi M, Fukunaga M, van Gelderen P, Horovitz SG, de Zwart JA, Duyn JH. Modulation of spontaneous fMRI activity in human visual cortex by behavioral state. Neuroimage. 2009;45:160–168. doi: 10.1016/j.neuroimage.2008.10.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Binder JR, Frost JA, Hammeke TA, Rao SM, Cox RW. Function of the left planum temporale in auditory and linguistic processing. Brain. 1996;119 (Pt 4):1239–1247. doi: 10.1093/brain/119.4.1239. [DOI] [PubMed] [Google Scholar]
- Brett M, Anton JL, Valabregue R, Poline JB. Region of interest analysis using an SPM toolbox. Eighth International Conference on Functional Mapping of the Human Brain; Sendai, Japan. 2002. [Google Scholar]
- Brown KW, Ryan RM. The benefits of being present: mindfulness and its role in psychological well-being. J Pers Soc Psychol. 2003;84:822–848. doi: 10.1037/0022-3514.84.4.822. [DOI] [PubMed] [Google Scholar]
- Brown KW, Ryan RM, Creswell JD. Mindfulness: Theoretical foundations and evidence for its salutary effects. Psychological Inquiry. 2007;18:1–27. [Google Scholar]
- Buckner RL, Andrews-Hanna JR, Schacter DL. The brain’s default network: anatomy, function, and relevance to disease. Ann N Y Acad Sci. 2008;1124:1–38. doi: 10.1196/annals.1440.011. [DOI] [PubMed] [Google Scholar]
- Burgess N. Spatial cognition and the brain. Ann N Y Acad Sci. 2008;1124:77–97. doi: 10.1196/annals.1440.002. [DOI] [PubMed] [Google Scholar]
- Celesia GG. Organization of auditory cortical areas in man. Brain. 1976;99:403–414. doi: 10.1093/brain/99.3.403. [DOI] [PubMed] [Google Scholar]
- Chambers R, Lo BCY, Allen NB. The impact of intensive mindfulness training on attentional control, cognitive style, and affect. Cognitive Therapy and Research. 2008;32:303–322. [Google Scholar]
- Chang C, Glover GH. Time-frequency dynamics of resting-state brain connectivity measured with fMRI. Neuroimage. 2010;50:81–98. doi: 10.1016/j.neuroimage.2009.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiesa A, Serretti A. Mindfulness-based stress reduction for stress management in healthy people: a review and meta-analysis. J Altern Complement Med. 2009;15:593–600. doi: 10.1089/acm.2008.0495. [DOI] [PubMed] [Google Scholar]
- Chiesa A, Serretti A. A systematic review of neurobiological and clinical features of mindfulness meditations. Psychol Med. 2010;40:1239–1252. doi: 10.1017/S0033291709991747. [DOI] [PubMed] [Google Scholar]
- Creswell JD, Myers HF, Cole SW, Irwin MR. Mindfulness meditation training effects on CD4+ T lymphocytes in HIV-1 infected adults: A small randomized controlled trial. Brain, Behavior, and Immunity. 2009;23:184–188. doi: 10.1016/j.bbi.2008.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Creswell JD, Way BM, Eisenberger NI, Lieberman MD. Neural correlates of dispositional mindfulness during affect labeling. Psychosom Med. 2007;69:560–565. doi: 10.1097/PSY.0b013e3180f6171f. [DOI] [PubMed] [Google Scholar]
- Damoiseaux JS, Rombouts SA, Barkhof F, Scheltens P, Stam CJ, Smith SM, Beckmann CF. Consistent resting-state networks across healthy subjects. Proc Natl Acad Sci U S A. 2006;103:13848–13853. doi: 10.1073/pnas.0601417103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deco G, Jirsa V, McIntosh AR, Sporns O, Kotter R. Key role of coupling, delay, and noise in resting brain fluctuations. Proc Natl Acad Sci U S A. 2009;106:10302–10307. doi: 10.1073/pnas.0901831106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff SB, Amunts K, Mohlberg H, Zilles K. The human parietal operculum. II. Stereotaxic maps and correlation with functional imaging results. Cereb Cortex. 2006;16:268–279. doi: 10.1093/cercor/bhi106. [DOI] [PubMed] [Google Scholar]
- Epstein RA. Parahippocampal and retrosplenial contributions to human spatial navigation. Trends Cogn Sci. 2008;12:388–396. doi: 10.1016/j.tics.2008.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esposito F, Pignataro G, Di Renzo G, Spinali A, Paccone A, Tedeschi G, Annunziato L. Alcohol increases spontaneous BOLD signal fluctuations in the visual network. Neuroimage. 2010 doi: 10.1016/j.neuroimage.2010.06.061. [DOI] [PubMed] [Google Scholar]
- Farb NA, Anderson AK, Mayberg H, Bean J, McKeon D, Segal ZV. Minding one’s emotions: mindfulness training alters the neural expression of sadness. Emotion. 2010;10:25–33. doi: 10.1037/a0017151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farb NA, Segal ZV, Mayberg H, Bean J, McKeon D, Fatima Z, Anderson AK. Attending to the present: mindfulness meditation reveals distinct neural modes of self-reference. Soc Cogn Affect Neurosci. 2007;2:313–322. doi: 10.1093/scan/nsm030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franco AR, Pritchard A, Calhoun VD, Mayer AR. Interrater and intermethod reliability of default mode network selection. Hum Brain Mapp. 2009;30:2293–2303. doi: 10.1002/hbm.20668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galaburda A, Sanides F. Cytoarchitectonic organization of the human auditory cortex. J Comp Neurol. 1980;190:597–610. doi: 10.1002/cne.901900312. [DOI] [PubMed] [Google Scholar]
- Goldin PR, Gross JJ. Effects of mindfulness-based stress reduction (MBSR) on emotion regulation in social anxiety disorder. Emotion. 2010;10:83–91. doi: 10.1037/a0018441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gopinath K, Ringe W, Goyal A, Carter K, Dinse HR, Haley R, Briggs R. Striatal functional connectivity networks are modulated by fMRI resting state conditions. Neuroimage. 2010 doi: 10.1016/j.neuroimage.2010.07.021. [DOI] [PubMed] [Google Scholar]
- Greicius M. Resting-state functional connectivity in neuropsychiatric disorders. Curr Opin Neurol. 2008;21:424–430. doi: 10.1097/WCO.0b013e328306f2c5. [DOI] [PubMed] [Google Scholar]
- Greicius MD, Flores BH, Menon V, Glover GH, Solvason HB, Kenna H, Reiss AL, Schatzberg AF. Resting-state functional connectivity in major depression: abnormally increased contributions from subgenual cingulate cortex and thalamus. Biol Psychiatry. 2007;62:429–437. doi: 10.1016/j.biopsych.2006.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Himberg J, Hyvarinen A, Esposito F. Validating the independent components of neuroimaging time series via clustering and visualization. Neuroimage. 2004;22:1214–1222. doi: 10.1016/j.neuroimage.2004.03.027. [DOI] [PubMed] [Google Scholar]
- Holzel BK, Ott U, Hempel H, Hackl A, Wolf K, Stark R, Vaitl D. Differential engagement of anterior cingulate and adjacent medial frontal cortex in adept meditators and non-meditators. Neurosci Lett. 2007;421:16–21. doi: 10.1016/j.neulet.2007.04.074. [DOI] [PubMed] [Google Scholar]
- Horovitz SG, Fukunaga M, de Zwart JA, van Gelderen P, Fulton SC, Balkin TJ, Duyn JH. Low frequency BOLD fluctuations during resting wakefulness and light sleep: a simultaneous EEG-fMRI study. Hum Brain Mapp. 2008;29:671–682. doi: 10.1002/hbm.20428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jha AP, Krompinger J, Baime MJ. Mindfulness training modifies subsystems of attention. Cogn Affect Behav Neurosci. 2007;7:109–119. doi: 10.3758/cabn.7.2.109. [DOI] [PubMed] [Google Scholar]
- Kabat-Zinn J. Full catastrophe living: using the wisdom of your body and mind to face stress, pain, and illness. Dell Publishing; New York, NY: 1990. [Google Scholar]
- Kelly AM, Uddin LQ, Biswal BB, Castellanos FX, Milham MP. Competition between functional brain networks mediates behavioral variability. Neuroimage. 2008;39:527–537. doi: 10.1016/j.neuroimage.2007.08.008. [DOI] [PubMed] [Google Scholar]
- Kiviniemi VJ, Haanpaa H, Kantola JH, Jauhiainen J, Vainionpaa V, Alahuhta S, Tervonen O. Midazolam sedation increases fluctuation and synchrony of the resting brain BOLD signal. Magn Reson Imaging. 2005;23:531–537. doi: 10.1016/j.mri.2005.02.009. [DOI] [PubMed] [Google Scholar]
- Laurienti PJ, Burdette JH, Wallace MT, Yen YF, Field AS, Stein BE. Deactivation of sensory-specific cortex by cross-modal stimuli. J Cogn Neurosci. 2002;14:420–429. doi: 10.1162/089892902317361930. [DOI] [PubMed] [Google Scholar]
- Lazar SW, Kerr CE, Wasserman RH, Gray JR, Greve DN, Treadway MT, McGarvey M, Quinn BT, Dusek JA, Benson H, Rauch SL, Moore CI, Fischl B. Meditation experience is associated with increased cortical thickness. Neuroreport. 2005;16:1893–1897. doi: 10.1097/01.wnr.0000186598.66243.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis CM, Baldassarre A, Committeri G, Romani GL, Corbetta M. Learning sculpts the spontaneous activity of the resting human brain. Proc Natl Acad Sci U S A. 2009;106:17558–17563. doi: 10.1073/pnas.0902455106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y, Adali T, Calhoun VD. Sample Dependence correction for order selection in fMRI analysis. Proc ISBI; Washington, D.C. 2006. [Google Scholar]
- Ludwig DS, Kabat-Zinn J. Mindfulness in medicine. Jama. 2008;300:1350–1352. doi: 10.1001/jama.300.11.1350. [DOI] [PubMed] [Google Scholar]
- Lutz A, Brefczynski-Lewis J, Johnstone T, Davidson RJ. Regulation of the neural circuitry of emotion by compassion meditation: effects of meditative expertise. PLoS One. 2008a;3:e1897. doi: 10.1371/journal.pone.0001897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lutz A, Slagter HA, Dunne JD, Davidson RJ. Attention regulation and monitoring in meditation. Trends Cogn Sci. 2008b;12:163–169. doi: 10.1016/j.tics.2008.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marx E, Stephan T, Nolte A, Deutschlander A, Seelos KC, Dieterich M, Brandt T. Eye closure in darkness animates sensory systems. Neuroimage. 2003;19:924–934. doi: 10.1016/s1053-8119(03)00150-2. [DOI] [PubMed] [Google Scholar]
- McAvoy M, Larson-Prior L, Nolan TS, Vaishnavi SN, Raichle ME, d’Avossa G. Resting states affect spontaneous BOLD oscillations in sensory and paralimbic cortex. J Neurophysiol. 2008;100:922–931. doi: 10.1152/jn.90426.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer M, Zysset S, von Cramon DY, Alter K. Distinct fMRI responses to laughter, speech, and sounds along the human peri-sylvian cortex. Brain Res Cogn Brain Res. 2005;24:291–306. doi: 10.1016/j.cogbrainres.2005.02.008. [DOI] [PubMed] [Google Scholar]
- Modinos G, Ormel J, Aleman A. Individual differences in dispositional mindfulness and brain activity involved in reappraisal of emotion. Soc Cogn Affect Neurosci. 2010 doi: 10.1093/scan/nsq006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nir Y, Hasson U, Levy I, Yeshurun Y, Malach R. Widespread functional connectivity and fMRI fluctuations in human visual cortex in the absence of visual stimulation. Neuroimage. 2006;30:1313–1324. doi: 10.1016/j.neuroimage.2005.11.018. [DOI] [PubMed] [Google Scholar]
- Northoff G, Bermpohl F. Cortical midline structures and the self. Trends Cogn Sci. 2004;8:102–107. doi: 10.1016/j.tics.2004.01.004. [DOI] [PubMed] [Google Scholar]
- Northoff G, Heinzel A, Bermpohl F, Niese R, Pfennig A, Pascual-Leone A, Schlaug G. Reciprocal modulation and attenuation in the prefrontal cortex: an fMRI study on emotional-cognitive interaction. Hum Brain Mapp. 2004;21:202–212. doi: 10.1002/hbm.20002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Northoff G, Heinzel A, de Greck M, Bermpohl F, Dobrowolny H, Panksepp J. Self-referential processing in our brain--a meta-analysis of imaging studies on the self. Neuroimage. 2006;31:440–457. doi: 10.1016/j.neuroimage.2005.12.002. [DOI] [PubMed] [Google Scholar]
- O’Craven KM, Kanwisher N. Mental imagery of faces and places activates corresponding stiimulus-specific brain regions. J Cogn Neurosci. 2000;12:1013–1023. doi: 10.1162/08989290051137549. [DOI] [PubMed] [Google Scholar]
- Peltier SJ, LaConte SM, Niyazov DM, Liu JZ, Sahgal V, Yue GH, Hu XP. Reductions in interhemispheric motor cortex functional connectivity after muscle fatigue. Brain Res. 2005;1057:10–16. doi: 10.1016/j.brainres.2005.06.078. [DOI] [PubMed] [Google Scholar]
- Rack-Gomer AL, Liau J, Liu TT. Caffeine reduces resting-state BOLD functional connectivity in the motor cortex. Neuroimage. 2009;46:56–63. doi: 10.1016/j.neuroimage.2009.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seeley WW, Menon V, Schatzberg AF, Keller J, Glover GH, Kenna H, Reiss AL, Greicius MD. Dissociable intrinsic connectivity networks for salience processing and executive control. J Neurosci. 2007;27:2349–2356. doi: 10.1523/JNEUROSCI.5587-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slagter HA, Lutz A, Greischar LL, Francis AD, Nieuwenhuis S, Davis JM, Davidson RJ. Mental training affects distribution of limited brain resources. PLoS Biol. 2007;5:e138. doi: 10.1371/journal.pbio.0050138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith SM, Fox PT, Miller KL, Glahn DC, Fox PM, Mackay CE, Filippini N, Watkins KE, Toro R, Laird AR, Beckmann CF. Correspondence of the brain’s functional architecture during activation and rest. Proc Natl Acad Sci U S A. 2009;106:13040–13045. doi: 10.1073/pnas.0905267106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spielberger CD, Gorsuch RL, Lushene RE. Manual for the State-Trait Anxiety Inventory. Consulting Psychologists Press; Palo Alto, CA: 1970. [Google Scholar]
- Valentine ER, Sweet PLG. Meditation and attention: A comparison of the effects of concentrative and mindfulness meditation on sustained attention. Mental Health, Religion, and Culture. 1999;2:59–70. [Google Scholar]
- van den Hurk PAM, Giommi F, Gielen SC, Speckens AEM, Barendregt HP. Greater efficiency in attentional processing related to mindfulness meditation. Quarterly Journal of Experimental Psychology. 2010a;63:1168–1180. doi: 10.1080/17470210903249365. [DOI] [PubMed] [Google Scholar]
- van den Hurk PAM, Janssen BH, Giommi F, Barendregt HP, Gielen SC. Mindfulness meditation associated with alterations in bottom-up processing: Psychophysiological evidence for reduced reactivity. International Journal of Psychophysiology. 2010b;78:151–157. doi: 10.1016/j.ijpsycho.2010.07.002. [DOI] [PubMed] [Google Scholar]
- Walter M, Matthia C, Wiebking C, Rotte M, Tempelmann C, Bogerts B, Heinze HJ, Northoff G. Preceding attention and the dorsomedial prefrontal cortex: process specificity versus domain dependence. Hum Brain Mapp. 2009;30:312–326. doi: 10.1002/hbm.20506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan C, Liu D, He Y, Zou Q, Zhu C, Zuo X, Long X, Zang Y. Spontaneous brain activity in the default mode network is sensitive to different resting-state conditions with limited cognitive load. PLoS One. 2009;4:e5743. doi: 10.1371/journal.pone.0005743. [DOI] [PMC free article] [PubMed] [Google Scholar]