

Abstract
Expression of Mash1 is dysregulated in human neuroblastoma. We have also reported that LMO3 (LIM-only protein 3) has an oncogenic potential in collaboration with neuronal transcription factor HEN2 in neuroblastoma. However, the precise molecular mechanisms of its transcriptional regulation remain elusive. Here we found that LMO3 forms a complex with HEN2 and acts as an upstream mediator for transcription of Mash1 in neuroblastoma. The high levels of LMO3 or Mash1 mRNA expression were significantly associated with poor prognosis in 100 primary neuroblastomas. The up-regulation of Mash1 remarkably accelerated the proliferation of SH-SY5Y neuroblastoma cells, while siRNA-mediated knockdown of LMO3 induced inhibition of growth of SH-SY5Y cells in association with a significant down-regulation of Mash1. Additionally, overexpression of both LMO3 and HEN2 induced expression of Mash1, suggesting that they might function as a transcriptional activator for Mash1. Luciferase reporter assay demonstrated that the co-expression of LMO3 and HEN2 attenuates HES1 (a negative regulator for Mash1)-dependent reduction of luciferase activity driven by the Mash1 promoter. Chromatin immunoprecipitation assay revealed that LMO3 and HEN2 reduce the amount of HES1 recruited onto putative HES1-binding sites and E-box within the Mash1 promoter. Furthermore, both LMO3 and HEN2 are physically associated with HES1 by immunoprecipitation assay. Thus, our present results suggest that a transcriptional complex of LMO3 and HEN2 may contribute to the genesis and malignant phenotype of neuroblastoma by inhibiting HES1 which suppresses the transactivation of Mash1.
Introduction
Neuroblastoma is one of the typical childhood cancers and is originated from sympathetic cell lineage of the neural crest [1], [2]. Since the tumor never occurs from the other lineages of the neural crest, the oncogenic events to cause neuroblastoma might be strictly regulated in a lineage-specific manner [1], [2].
LIM-only protein (LMO) family is composed of four members, LMO1, LMO2, LMO3 and LMO4. Although LMO proteins lack a DNA-binding activity, accumulating evidence suggest that LMO proteins are involved in transcriptional regulation of specific target genes in collaboration with other transcription factors [3]. Genetic analyses demonstrated that LMO1 and LMO2 contribute to the genesis of immature and aggressive T-cell leukemia [4], whereas LMO4 was implicated in development of breast cancer [5], [6]. Previously, we reported that LMO3 is expressed at significantly high levels in human unfavorable neuroblastomas relative to favorable ones, and has an oncogenic potential in neuroblastoma [7]. LMO3 formed a complex with neuronal-specific basic helix-loop-helix (bHLH) transcription factor HEN2, which was also expressed at higher levels in unfavorable neuroblastoma than favorable one, raising a possibility that LMO3 may form a complex with HEN2 and play an important role in genesis and development of neuroblastoma through transcriptional regulation of as yet unidentified target gene(s).
A proneural bHLH transcription factor termed Mash1 plays a critical role in development of sympathetic neuron and is highly expressed in neuroblastoma [8], [9]. However, its possible contribution to development of neuroblastoma remains elusive. A bHLH protein termed HES1 acts as a negative regulator for Mash1 [10]. Intriguingly, studies in Drosophila demonstrated that expression levels of achaete-scute, a Drosophila homolog of Mash1, are remarkably induced by a transcriptional complex composed of Drosophila homolog of LMO (dLMO) and bHLH proteins [11], [12].
In this study, we examined whether there could exist functional relationship between LMO3/HEN2 and Mash1 in neuroblastoma, and found that LMO3/HEN2 attenuates HES1 function and enhances transactivation of Mash1, leading to aggressive phenotype of neuroblastoma.
Results
High levels of Mash1 expression is associated with poor outcome of neuroblastoma
Mash1 is constitutively expressed at high levels in neuroblastoma cell lines and primary neuroblastoma tumors [9], [13], however, its prognostic significance remained elusive. On the other hand, expression of LMO3 was significantly associated with poor outcome of the patients [7]. To verify whether a significant relationship could be observed between expression of LMO3 and that of Mash1 in primary neuroblastomas, we quantitatively measured the expression levels of LMO3 and Mash1 mRNA in 100 primary tumors by using a quantitative real-time RT-PCR. The student's t-test showed that high expression of LMO3 was significantly associated with ≥1 year of age (p = 0.036), low expression of TrkA (p = 0.003) and MYCN amplification (p = 0.04), but not with the tumor stage (p = 0.17), tumor origin (p = 0.083) and Shimada classification (p = 0.082). High expression of Mash1 was significantly associated with advanced tumor stage (p = 0.004) but not with age (p = 0.81), TrkA expression (p = 0.4), MYCN copy number (p = 0.11), tumor origin (p = 0.2) and Shimada classification (p = 0.45) (Table S1). No significant relationship was observed between LMO3 and Mash1 mRNA expression levels (the Pearson correlation coefficient was 0.27). Kaplan-Meier survival curves indicated that high expression of LMO3 as well as that of Mash1 were significantly associated with poor prognosis (log-rank test, p = 0.006 and p = 0.037, respectively; Figure 1). The univariate analysis according to the Cox proportional hazard model also indicated that the expression levels of Mash1 and those of LMO3 were significantly associated with poor outcome of the patients (p = 0.048 and p = 0.012, respectively; Table S2). The multivariate Cox proportional hazard model analysis showed that the expression of Mash1 was significantly independent prognostic factor from LMO3 expression and age, marginally from MYCN copy number and origin, but not from the disease stage, and that the expression of LMO3 was significantly independent prognostic factor from Mash1 expression, age, the disease stage and origin, but not from MYCN copy number (Table S2). Thus, the results obtained from the primary neuroblastomas suggested that both high mRNA expression of LMO3 and Mash1 were strongly associated with poor prognoses of the patients with neuroblastoma but the way of contribution of those seemed to be rather independent.
Figure 1. Kaplan-Meier survival curves of patients with neuroblastomas based on high or low expression of LMO3 (A) or Mash1 (B).

Kaplan-Meier survival curves (n = 100) in relation to the expression levels of LMO3 or Mash1 (average cutoff). The patients with high expression of LMO3 or Mash1 represented significantly poor prognosis than those with its low expression.
Mash1 mediates growth promotion of neuroblastoma cells
Since Mash1 is highly expressed in primary neuroblastoma [9] and its higher expression was significantly correlated with poor prognosis of the patient with neuroblastoma, we then investigated a possible contribution of Mash1 to neuroblastoma cell growth. For this purpose, we established three stable Mash1 infectants derived from the parental SH-SY5Y neuroblastoma cells expressing exogenous Mash1 (M-1, M-2 and M-3) and two control vector alone infectants (V-1 and V-2) by retrovirus-mediated gene transfer (Figure 2A). As shown in Figure 2B, constitutive expression of Mash1 in SH-SY5Y cells resulted in a remarkable increase in their growth rate as compared with the control infectants, suggesting that Mash1 is involved in regulation of neuroblastoma cell growth.
Figure 2. Mash1-mediated growth promotion of neuroblastoma cells.

(A) Enforced expression of Mash1. Neuroblastoma SH-SY5Y cells were infected with empty retrovirus or with retrovirus encoding Mash1 and established two control infectants (V-1 and V-2) and three infectants expressing Mash1 (M-1, M-2 and M-3). Total RNA was extracted from the indicated cell clones and subjected to RT-PCR to examine expression levels of Mash1. GAPDH was used as an internal control. (B) Mash1-mediated growth promotion. The indicated infectants were seeded at a density of 3×104/cell culture dish and allowed to attach overnight. At the indicated time periods, number of viable cells was measured. (C) siRNA-mediated knockdown of LMO3. SH-SY5Y cells were transfected with empty plasmid (4 µg) or with expression plasmid for siRNA targeting LMO3 (4 µg). Forty-eight hours after transfection, total RNA was prepared and analyzed for expression levels of LMO3 and Mash1 by RT-PCR. (D) Decreased growth rate in LMO3-knocked down cells. SH-SY5Y cells (3×105 cells/cell culture dish) were transfected as in (C). Forty-eight hours after transfection, cells were transferred into fresh medium. At the indicated time points, number of viable cells was measured.
As described previously [7], LMO3 has an oncogenic potential in collaboration with HEN2 in neuroblastoma cells. We then asked whether or not LMO3 is involved in the Mash1-mediated enhancement of cell growth. As shown in Figure 2C, siRNA-mediated knockdown of LMO3 in SH-SY5Y cells was significantly associated with a down-regulation of Mash1. Additionally, LMO3-knocked down SH-SY5Y cells showed a slower growth rate than the control SH-SY5Y cells (Figure 2D), which might be at least in part due to reduction of Mash1. We conducted the same experiments by using another cell line SK-N-BE and obtained the similar results (Figure S1A and B). We then hypothesized that Mash1 could be one of transcriptional targets of LMO3/HEN2 complex.
LMO3/HEN2 mediate transcriptional induction of Mash1
To address whether Mash1 transcription could be induced by LMO3/HEN2, SH-SY5Y cells were infected with the indicated combinations of recombinant adenoviruses encoding HA-LMO3 or FLAG-HEN2, and the expression levels of Mash1 were examined by semi-quantitative RT-PCR. Time course experiments demonstrated that Mash1 is readily detectable in cells expressing HA-LMO3 alone or in cells co-expressing with HA-LMO3 and FLAG-HEN2 at 48 h after infection (Figure 3A). Seventy-two hours after infection, co-expression of HA-LMO3 and FLAG-HEN2 led to a significant induction of Mash1. The induction of Mash1 was also observed in SK-N-BE cells transfected with expression vector HA-LMO3 alone or HA-LMO3 and FLAG-HEN2 at 72 h after transfection (Figure S1C). To further confirm these observations, we generated a luciferase reporter construct carrying human Mash1 promoter (pluc-Mash1). As shown in Figure 3B, the 5′-upstream region of Mash1 gene contains three putative HES1-binding sites and one E-box. In both SH-SY5Y cells and SK-N-BE cells, siRNA-mediated knockdown of human LMO3 reduced promoter activity of Mash1 in a dose-dependent manner (Figure 3C and Figure S1D). For luciferase reporter assay without siRNA for human LMO3, we used mouse neuroblastoma Neuro2a cells which displayed higher transfection efficiency than human neuroblastoma cells as examined by GFP staining (data not shown). Consistent with the above expression studies, LMO3 enhanced luciferase activity driven by Mash1 promoter (Figure 3D). Furthermore, we examined the effect of HEN2 on Mash1 promoter activity in Neuro2a cells, showing that HEN2 itself inhibited Mash1 promoter activity (Figure 3E). Intriguingly, however, LMO3 interfered with HEN2 function, resulting in up-regulation of Mash1 transcription (Figure 3F). Thus, it is likely that the LMO3 complex including HEN2 and HES1 regulates transcription of Mash1. The mRNA expression pattern of LMO3, HEN2, Mash1 and HES1, a negative regulator of Mash1 transcription, in neuroblastoma cell lines is shown in Figure S2.
Figure 3. LMO3/HEN2-mediated transcriptional induction of Mash1.

(A) RT-PCR. SH-SY5Y cells were infected with empty adenovirus or with the indicated combinations of recombinant adenovirus encoding HA-LMO3 or FLAG-HEN2. At the indicated time points after infection, total RNA was analyzed for expression levels of LMO3, HEN2 and Mash1 by RT-PCR. GAPDH was used as an internal control. (B) Schematic drawing of human Mash1 promoter. Nucleotide positions were indicated relative to transcriptional initiation site (+1). The putative HES1-binding sites and E-box were depicted by filled and open boxes, respectively. This genomic fragment was subcloned into appropriate restriction sites of pGL3-Basic Vector to give pluc-hMash1. (C) siRNA-mediated knockdown of LMO3 reduces the promoter activity of Mash1. SH-SY5Y cells were co-transfected with constant amount of pluc-Mash1 (100 ng) and pRL-CMV (0.2 ng) in the presence or absence of increasing amounts of expression plasmid for siRNA against human LMO3 (100 or 400 ng). Forty-eight hours after transfection, cells were lysed and their luciferase activities were measured. (D) LMO3 transactivates Mash1 promoter. Mouse neuroblastoma Neuro2a cells (1×105 cells/24-well plate) were co-transfected with constant amount of pluc-hMash1 (100 ng) and pRL-CMV (0.2 ng) together with or without expression plasmid for HA-LMO3 (150 ng). Forty-eight hours after transfection, cells were lysed and their luciferase activities were measured. (E) HEN2 inhibits Mash1 promoter activity. Luciferase activities were measured in Neuro2a cells with or without FLAG-HEN2 (100 ng). (F) LMO3 interferes with negative effect of HEN2 on Mash1 transcription in Neuro2a cells. Luciferase activities were measured in Neuro2a cells transfected with HA-LMO3 (150 ng), FLAG-HEN2 (100 ng) or both of them.
LMO3/HEN2 attenuates HES1-dependent down-regulation of Mash1
As reported previously [10], HES1 is one of the negative regulators for Mash1. In accordance with the previous observations, enforced expression of HES1 dramatically reduced luciferase activity driven by Mash1 promoter (Figure 4A). The inhibitory effect of HES1 on Mash1 promoter was stronger than that of HEN2. To investigate the relationships between HES1 and LMO3/HEN2 in transcriptional regulation of Mash1, we examined effects of HEN2 and LMO3 on HES1-dependent down-regulation of Mash1 (Figure 4B). The HES1-dependent inhibition of Mash1 promoter activity was attenuated by co-expression with FLAG-HEN2 alone or with co-expression with FLAG-HEN2 plus HA-LMO3. Inhibitory effects of FLAG-HEN2 plus HA-LMO3 on HES1 were larger than that of FLAG-HEN2 alone, suggesting that LMO3/HEN2 complex plays a critical role in regulation of Mash1 transcription by neutralizing the inhibitory effect of HES1.
Figure 4. LMO3/HEN2 attenuates HES1-dependent down-regulation of Mash1.

(A) Luciferase reporter assay. Neuro2a cells were co-transfected with constant amount of pluc-hMash1 (100 ng), pRL-CMV (0.2 ng) and expression plasmid for HES1 (50 ng) or HEN2 (50 ng). Forty-eight hours after transfection, cells were lysed and their luciferase activities were examined. (B) Luciferase reporter assay. Neuro2a cells were co-transfected with constant amount of pluc-hMash1 (100 ng), pRL-CMV (0.2 ng) and expression plasmid for HES1 (5 ng) in the presence or absence of expression plasmid for HA-LMO3 (150 ng) together with or without increasing amounts of FLAG-HEN2 expression plasmid (100, 200 or 300 ng). Forty-eight hours after transfection, cells were lysed and their luciferase activities were examined. (C) Schematic representation of mouse Mash1 promoter. The canonical HES1-binding sites and E-box were indicated by filled and open boxes, respectively. The positions of primer sets used for chromatin immunoprecipitation (ChIP) assays were also indicated. (D) ChIP assay. Cross-linked chromatin prepared from Neuro2a cells transfected with the indicated combinations of expression plasmids was sonicated and immunoprecipitated with normal rabbit serum (NRS), polyclonal anti-Myc tag or with polyclonal anti-FLAG antibody. The genomic DNA was purified from the immunoprecipitates and amplified by PCR.
To ask about mechanistic insights into understanding how LMO3 and/or HEN2 could attenuate the inhibitory effects of HES1 on Mash1 expression, we performed chromatin immunoprecipitation (ChIP) assay. Similar to human Mash1 promoter, mouse Mash1 promoter also contains three putative HES1-binding sites and one E-box (Figure 4C). Neuro2a cells were transfected with constant amount of empty plasmid or with expression plasmid for Myc-HES1 together with or without increasing amounts of FLAG-HEN2 expression plasmid. Forty-eight hours after transfection, cross-linked chromatin was prepared and subjected to ChIP assay. As shown in Figure 4D, the anti-Myc tag immunoprecipitates contained genomic fragments including putative HES1-binding sites as well as E-box. The amounts of Myc-HES1 recruited onto HES1-binding sites and E-box significantly decreased in the presence of FLAG-HEN2 in a dose-dependent manner. Additionally, the anti-FLAG immunoprecipitates contained genomic fragments including putative HES1-binding sites and E-box in the absence of exogenous HES1. Co-expression of FLAG-HEN2 and Myc-HES1 inhibited recruitment of FLAG-HEN2 onto putative HES1-binding sites and E-box, however, its inhibition was efficiently abrogated by increasing amounts of FLAG-HEN2. These results suggest that HEN2 might compete with HES1 in binding to putative HES1-binding sites and E-box, and thereby inducing the expression of Mash1.
HEN2 Interacts with HES1 in cells
To examine whether HEN2 could interact with HES1 in cells, we performed immunoprecipitation experiments. Cell lysates prepared from Neuro2a cells co-transfected with the indicated combinations of expression plasmids were subjected to immunoprecipitation. As clearly shown in Figure 5A, HES1 was co-immunoprecipitated with FLAG-HEN2. Consistent with these results, reciprocal experiments showed that the anti-Myc tag immunoprecipitates contain FLAG-HEN2. In vitro pull-down assay demonstrated that radio-labeled FLAG-HEN2 is co-immunoprecipitated with Myc-HES1 (Figure 5B). Additional immunoprecipitation experiments demonstrated that LMO3 also forms a stable complex with HES1 (Figure 6A). We have previously showed that LMO3 forms a stable complex with HEN2 [7]. To investigate the effect of LMO3 on binding of HEN2 and HES1 to putative HES1-binding sites and E-box, Neuro2a cells were transfected with Myc-HES1 or FLAG-HEN2 together with or without HA-LMO3 expression plasmid and subjected to ChIP assay. As shown in Figure 6B, the immunoprecipitates using anti-Myc tag or anti-FLAG tag antibody contained genomic fragments including putative HES1-binding sites as well as E-box. The amount of Myc-HES1 recruited onto HES1-binding sites and E-box decreased in the presence of HA-LMO3. On the other hand, the amount of FLAG-HEN2 recruited onto HES1-binding sites and E-box increased in the presence of HA-LMO3. As shown in Figure 3F, LMO3 interferes with inhibitory effect of HEN2 on Mash1 expression. These suggest that LMO3 may additively interfere with the inhibitory effect of HES1 on Mash1 expression by promoting binding of HEN2 to HES1-binding sites and E-box. Collectively, it is conceivable that LMO3/HEN2 reduces the inhibitory effect of HES1 on Mash1 expression through binding to HES1 and thereby blocking its recruitment onto putative HES1-binding sites and E-box (Figure 7 and Figure S3).
Figure 5. Interaction between HEN2 and HES1 in cells.

(A) Neuro2a cells were co-transfected with the indicated combinations of expression plasmids. Forty-eight hours after transfection, cells were lysed and immunoprecipitated with anti-FLAG (left panel) or with anti-Myc tag antibody (right panel) and the immunoprecipitates were analyzed by immunoblotting with anti-HES1 or with anti-FLAG antibody, respectively. Aliquots of cell lysates were subjected to immunoblotting with anti-HES1, anti-FLAG or with anti-Myc tag antibody. (B) In vitro pull-down assay. Radio-labeled FLAG-HEN2 was incubated with cell lysates prepared from Neuro2a cells transfected with Myc-HES1 expression plasmid. The reaction mixture was immunoprecipitated with normal rabbit serum (NRS) or with polyclonal anti-Myc tag antibody and separated by SDS-PAGE followed by autoradiography. 1/5 inputs were also shown.
Figure 6. LMO3 attenuates binding of HES1 to Mash1 promoter and promotes that of HEN2.

(A) Complex formation between LMO3 and HES1 in cells. Neuro2a cells were transiently transfected with the indicated combinations of the expression plasmids. Forty-eight hours after transfection, cell lysates were immunoprecipitated with anti-FLAG antibody followed by immunoblotting with anti-Myc tag antibody (top panel). Expressions of FLAG-LMO3 and Myc-HES1 are also shown (lower panels). (B) ChIP assay. Cross-linked chromatin prepared from Neuro2a cells transfected with the indicated combinations of expression plasmids was sonicated and immunoprecipitated with normal rabbit serum (NRS), polyclonal anti-Myc tag or with polyclonal anti-FLAG antibody. The genomic DNA was purified from the immunoprecipitates and amplified by PCR.
Figure 7. Model for LMO3 and HEN2 cooperation in transcriptional regulation of Mash1 in Neuroblastoma.
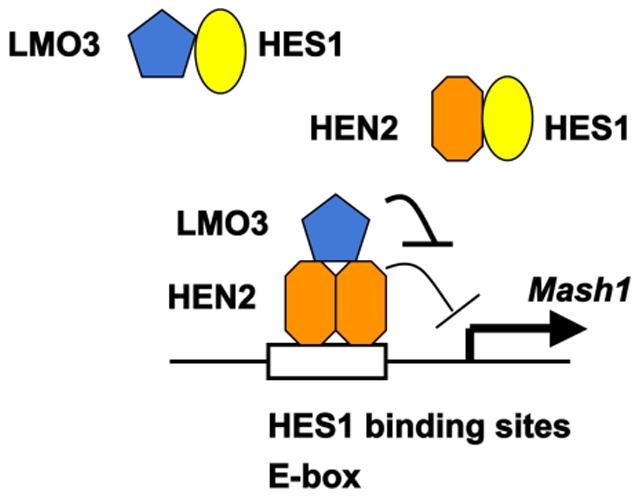
HES1 binds to HES1 binding sites and E-box on Mash1 promoter and represses Mash1 transcription. LMO3 inhibits recruitment of HES1 onto HES1-binding sites and E-box on Mash1 promoter by forming complex with HES1. HEN2 interferes with recruitment of HES1 onto HES1-binding sites and E-box on Mash1 promoter by forming complex with HES1 and competing with HES1 in binding to these sites. LMO3 promotes recruitment of HEN2 onto HES1-binding sites and E-box on Mash1 promoter by forming complex with HEN2 but inhibits negative effects of HEN2 on Mash1 promoter. Thereby expression of Mash1 is up-regulated.
Discussion
In this study, we found that Mash1 is one of transcriptional targets of LMO3/HEN2 transcriptional complex, and its protein product may play an important role in regulation of neuroblastoma cell growth. As described previously [14], HEN1 as well as its closely related gene HEN2 encodes bHLH-type transcription factor, which might recognize E-box (5′-CACGTG-3′). On the other hand, HES1 has an intrinsic transcriptional repressor activity [10]. Based on our present results, adenovirus-mediated expression of LMO3/HEN2 significantly induced Mash1, and HES1-mediated down-regulation of Mash1 promoter activity was recovered by co-expression of LMO3 and HEN2. Our ChIP analyses indicated that HES1 binds to HES1-recognition sites and E-box within Mash1 promoter in the absence of HEN2, whereas HEN2 efficiently inhibits the recruitment of HES1 onto HES1-binding sites and E-box within Mash1 promoter, suggesting that HES1 occupies HES1-binding sites and E-box to inhibit the promoter activity of Mash1. On the other hand, HEN2 formed a complex with HES1 and reduced the amounts of HES1 recruited onto HES1-binding sites as well as E-box to increase the promoter activity of Mash1 in collaboration with LMO3. Thus, it is likely that the balance between intracellular amounts of HES1 and LMO3/HEN2 might determine expression levels of Mash1, and thereby regulating neuroblastoma cell growth.
It was reported that de-repression of Mash1 might interfere with differentiation of sympatho-adrenal precursors of Insm1 mutant mice although Mash1 is expressed transiently in those cells during normal neural differentiation [15]. Furthermore, Watt et al. reported that N-myc positively regulates Mash1 transcription [16]. Therefore, it is possible that in the transcriptional regulation of Mash1, LMO3 and HEN2 may associate with other nuclear factors like Insm1 and N-myc besides HES1.
From the developmental point of view, it is known that the LMO/HEN complex plays an important role in regulating neuronal differentiation [11], [17]. As described [7], [18], expression of LMO3 was highly restricted in adult and fetal brains, and HEN2 was expressed in developing nervous system. Genetic studies demonstrated that HEN2 participates in proper neural crest-derived neuroendocrine development and that Mash1 has a critical role in maintaining neuroendocrine cell phenotype [19], [20]. Although LMO3-knockout mice did not exhibit any significant developmental defects, mice lacking both LMO1 and LMO3 died after birth, which might be due to neural defects [21]. Since neuroblastoma is one of the most common childhood solid tumors of peripheral nervous system arising from as yet unidentified population of neural crest cells [22] and Mash1 regulates proliferation of the sympathetic nervous system [23], it is likely that deregulated expression of Mash1 could contribute to genesis and development of neuroblastoma, which might be regulated by LMO3/HEN2 transcriptional complex both in vitro and in vivo. This LMO3/HEN2-HES1-Mash1 pathway could be the new future target for developing the anti-neuroblastoma treatment.
Materials and Methods
Ethics Statement
A hundred human neuroblastoma specimens used in the present study were kindly provided from various institutions and hospitals in Japan to the Chiba Cancer Center Neuroblastoma Tissue Bank. Written informed consent was obtained at each institution or hospital. This study was approved by the Chiba Cancer Center Institutional Review Board and were conducted according to the principles expressed in the Declaration of Helsinki.
Tumor Specimens
Tumors were classified according to the International Neuroblastoma Staging System (INSS); 25 Stage 1, 13 Stage 2, 33 Stage 3, 23 Stage 4, and 6 Stage4s. Clinical information including age at diagnosis, tumor origin, Shimada's histology, prognosis, and survival months of each patient were obtained. The median follow-up time for survivors was 35 months (range 3 to 91 months). Each tumor specimen was assayed for TRKA expression by Northern blot analysis and for MYCN amplification status by both fluorescence in situ hybridization (FISH) and real-time quantitative polymerase chain reaction (PCR).
Quantitative Real-time PCR
Total RNA prepared from primary neuroblastomas was reverse transcribed into cDNA (SuperScript II kit) and subjected to the real-time PCR. The expression level of GAPDH was measured in all samples to normalize LMO3 and Mash1 expression according to the manufacturer's instructions (Applied Biosystems, Foster City, CA, USA). Oligonucleotide primers and TaqMan probes, which were labeled at the 5′ end with the reporter dye 6-carboxyfluorescein (FAM) and at the 3′ end with the quencher dye 6-carboxytetramethylrhodamine (TAMRA), were as follows: LMO3: forward 5′-TCTGAGGCTCTTTGGTGTAACG-3′, reverse 5′-CCAGGTGGTAAACATTGTCCTTG-3′ and probe 5′-FAM-AAACTGCGCTGCCTGTAGTAAGCTCATCC-TAMRA-3′. Taqman(R) Gene Expression Assay (Applied Biosystems) was purchased for Mash1 with Assay ID Hs00269932-m1. Amplification and detection were done using the ABI Prism 7700 Sequence Detection System (Applied Biosystems).
Statistical Analysis
Student's t tests were used to explore possible associations between LMO3 expression and other factors. The distinction between high and low levels of LMO3 and Mash1 expression was based on the mean value. Kaplan-Meier survival curves were calculated, and survival distributions were compared using the log-rank test. Cox regression models were used to explore associations among LMO3 expression, Mash1 expression, age, MYCN amplification, tumor origin, Shimada classification and survival. Statistical significance was declared if P<0.05.
Cell Culture and Transfection
SH-SY5Y (human neuroblastoma, ATCC number CRL-2266), SK-N-BE (human neuroblastoma, ATCC number CRL-2271) and Neuro2a (mouse neuroblastoma, ATCC number CCL-131) cells were maintained in RPMI 1640 supplemented with 10% heat-inactivated fetal bovine serum at 37°C in an atmosphere of 5% CO2 in the air. Cells were transfected with the indicated expression plasmids using Lipofectamine 2000 transfection reagent (Invitrogen, Carlsbad, CA, USA) as recommended by the manufacturer.
Generation of Recombinant Retroviral Vector and Retrovirus-mediated Gene Transfer
Human Mash1 cDNA was subcloned into the HpaI restriction site of the pLXSN vector. pLXSN or pLXSN- Mash1 was transfected into the ϕ2 packaging cells, and SH-SY5Y cells (1×106 cells) infected with virus-containing culture medium were cultured in the medium containing 500 µg/ml G418 (Sigma Chemical Co., St. Louis, MO, USA). Two weeks after the selection in G418, drug-resistant clones were isolated and allowed to proliferate in medium containing G418.
Reverse Transcription-PCR Analysis
Total RNA was prepared from cultured cells by using the RNeasy Mini Kit (Qiagen, Valencia, CA, USA). Reverse transcription was carried out using random primers and SuperScript II (Invitrogen). Following the reverse transcription, the resultant cDNA was subjected to PCR-based amplification. PCR primers used were as follows: human LMO3, forward 5′-ATGCTCTCAGTCCAGCCAGA-3′ and reverse 5′-TCAGCGAACCTGGGGTGCAT-3′; human HEN2, forward 5′-AAGCAGCAGATTCGGACCAT-3′ and reverse 5′-CTTCTCCTCGCGGCTCAG-3′; human Mash1, forward 5′-GCGTTCAGCACTGACTTTTG-3′ and reverse 5′-CCCCGGGAGACTTCTTAGAG-3′; human HES1, forward 5′-TGAGCCAGCTGAAAACACTG-3′ and reverse 5′-GTCACCTCGTTCATGCACTC-3; human glyceraldehydes-3-phosphate dehydrogenase (GAPDH), forward 5′-ACCTGACCTGCCGTCTAGAA-3′ and reverse 5′-TCCACCACCCTGTTGCTGTA-3′.
RNA Interference Experiments
Human LMO3 RNAi vector was made using the original plasmid that is gift from A.K. Munirajan (Chiba Cancer Center Research Institute). The targeted sequence is 5′- GTAGTAAGCTCATCCCTGC -3′. RNAi construct was transiently transfected into SH-SY5Y cells using Lipofectamine 2000 transfection reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instruction.
Generation of Recombinant Adenoviral Vector
For construction of the adenovirus expression vector, an HA-tagged human LMO3 cDNA or a FLAG-tagged human HEN2 cDNA were inserted into the shuttle vector pHMCMV6 [24]. Efficient construction of a recombinant adenovirus vector by an improved in vitro ligation method [25]. The resultant shuttle vector was digested with I-CeuI and PI-SceI and subcloned into the identical restriction sites of the adenovirus expression vector pAdHM4. The recombinant adenovirus construct was digested with PacI and transfected into 293 cells to generate recombinant adenovirus.
Luciferase Reporter Assay
The reporter plasmid contains a 1.2-kb fragment of the human Mash1 promoter that was subcloned into the pGL3-Basic Vector (Promega Corp., Madison, WI, USA) upstream of the luciferase reporter gene. Cells were seeded in triplicates into 24-well plates (1×105 cells/well) 24 h prior to transfection. Cells were cotransfected with 100 ng of the reporter plasmid, 0.2 ng of pRL-CMV encoding Renilla luciferase cDNA, 5 ng of rat HES1expression vector, 150 ng of HA-LMO3, 100 to 300 ng of FLAG-HEN2 expression vectors. Total amount of plasmid DNA per transfection was kept constant with pcDNA3 (Invitrogen). At 48 h after transfection, luciferase activity was measured by a Dual-Luciferase Reporter Assay System (Promega), and the transfection efficiency was standardized against Renilla luciferase activity.
Chromatin Immunoprecipitation (ChIP) Assays
ChIP assay was performed according to the protocol recommended by Upstate (Lake Placid, NY, USA). Cross-linked chromatin prepared from Neuro2a cells transfected with expression plasmids was sonicated and immunoprecipitated with normal rabbit serum (NRS), polyclonal anti-Myc tag (Medical & Biological Laboratories, Nagoya, Japan) or with polyclonal anti-FLAG (Sigma, St. Louis, MO, USA) antibody. The genomic DNA was purified from the immunoprecipitates and amplified by PCR.
The primers used to amplify the mouse Mash1 promoters were as follows: HES1 binding site (PCR-1), forward 5′-ATTTCTAGAGCCACCCCCTG-3′ and reverse 5′-TTGTTGCAGTGCGTGCGCC-3′; HES1 binding site (PCR-2), forward 5′-AGTGCGCTCGGCACTGACTT-3′ and reverse 5′-CGCGGTTGGCTTCGGGAGCC-3′; E-box (PCR-3), forward 5′-ATGGAGAGTTTGCAAGGAGC-3′ and reverse 5′-CAGCCCCACGCGCAGCCCTG-3′.
Western Blot Analysis and Immunoprecipitation
After transfection, Neuro2a cells were placed on ice, washed twice with phosphate-buffered saline, and lysed in lysis buffer containing 25 mM Tris-HCl (pH 8.0), 137 mM NaCl, 2.7 mM KCl, 1% TritonX-100, 1 mM phenylmethylsulfonyl fluoride and protease inhibitor mixture (Sigma). Lysates were placed on ice for 30 min, sonicated briefly, and clarified by centrifugation at 15,000× g for 5 min at 4°C. Protein concentrations of the supernatants were determined by using a Bio-Rad protein assay. For immunoblot analysis, proteins were resolved by sodium dodecyl sulfatepolyacrylamide gel electrophoresis (SDS-PAGE) and electrotransferred onto a nitrocellulose membrane. The membrane filter was blocked with 2% gelatin in Tris-buffered saline (TBS) for 3 h at room temperature and then incubated with a primary antibody including monoclonal anti-rat HES1 (Medical & Biological Laboratories, Nagoya, Japan), monoclonal anti-FLAG (M2; Sigma) or monoclonal anti-Myc (9B11; Cell Signaling Technology, Danvers, MA, USA) antibody over night at 4°C. The membrane filter was then incubated with a goat anti-mouse secondary antibody conjugated to horseradish peroxidase (Cell Signaling Technology, Danvers, MA, USA) or goat anti-rat secondary antibody conjugated to horseradish peroxidase (Beckman Coulter, Marseille, France) for 1 h at room temperature and bound secondary antibody was detected by enhanced chemiluminescence (Amersham Pharmacia Biotech) according to the manufacturer's protocol. For Immunoprecipitation, equal amounts of cell lysates (2 mg) were precleared with 25 µl of protein G-Sepharose (Amersham Bioscience, Uppsala, Sweden). After brief centrifugation, immunoprecipitation was carried out by incubating the supernatant with anti-FLAG polyclonal (Sigma, St. Louis, MO, USA) or anti-Myc tag polyclonal antibody (Medical & Biological Laboratories, Nagoya, Japan) over night at 4°C. Immunocomplexes were precipitated with protein G-Sepharose beads (Amersham Biosciences) for 3 hours at 4°C. The immunoprecipitated proteins were resolved by SDS-PAGE and analyzed by Western blotting.
In vitro Pull-down Assay
Radio-labeled FLAG-HEN2 was generated by using in vitro transcription/translation system (Promega) and incubated with cell lysates prepared from Neuro2a cells transfected with Myc-HES1 expression plasmid. The reaction mixture was immunoprecipitated with normal rabbit serum (NRS) or with polyclonal anti-Myc tag antibody (Medical & Biological Laboratories, Nagoya, Japan) and separated by SDS-PAGE followed by autoradiography.
Data Analysis and Statistics
All values for statistical significance represent mean ± SD. We carried out comparisons between means using the Student's t-test. Statistical significance implies P<0.05.
Supporting Information
Mash1-mediated growth promotion and LMO3/HEN2-mediated transcriptional induction of Mash1 in SK-N-BE cells. (A) siRNA-mediated knockdown of LMO3. SK-N-BE cells were transfected with empty plasmid (4 µg) or with expression plasmid for siRNA targeting LMO3 (4 µg). Forty-eight hours after transfection, total RNA was prepared and analyzed for expression levels of LMO3 and Mash1 by RT-PCR. (B) Decreased growth rate in LMO3-knocked down cells. SK-N-BE cells (4.5×103 cells/well, 96 well culture plate) were transfected with empty plasmid (0.2 µg) or with expression plasmid for siRNA targeting LMO3 (0.2 µg). Forty-eight hours after transfection, cells were transferred into fresh medium. At the indicated time points, cell growth was measured by MTT assay (Cell Counting Kit-8, DOJINDO). (C) RT-PCR. SK-N-BE cells were transfected with pcDNA3 empty plasmid or with the indicated combinations of expression plasmid HA-LMO3 or FLAG-HEN2. At 72 hours after transfection, total RNA was analyzed for expression levels of LMO3, HEN2 and Mash1 by RT-PCR. GAPDH was used as an internal control. (D) siRNA-mediated knockdown of LMO3 reduces the promoter activity of Mash1. SK-N-BE cells were co-transfected with constant amount of pluc-Mash1 (100 ng) and pRL-CMV (0.2 ng) in the presence or absence of increasing amounts of expression plasmid for siRNA against human LMO3 (100 or 400 ng). Forty-eight hours after transfection, cells were lysed and their luciferase activities were measured.
(TIF)
Expression of LMO3 , HEN2 , Mash1 or HES1 in neuroblastoma cell lines. Semiquantitative RT-PCR analysis for expression of LMO3, HEN3, Mash1 or HES1 in neuroblastoma cell lines is performed under linear amplification conditions. Expression of GAPDH is shown as a control.
(TIF)
Model for LMO3 and HEN2 cooperation in transcriptional regulation of Mash1 in Neuroblastoma. (A) HES1 binds to HES1 binding sites and E- box on Mash1 promoter and represses Mash1 transcription. (B) LMO3 inhibits recruitment of HES1 onto HES1-binding sites and E-box on Mash1 promoter by forming complex with HES1, and thereby inducing the expression of Mash1. (C) HEN2 interferes with recruitment of HES1 onto HES1-binding sites and E-box on Mash1 promoter by forming complex with HES1 and competing with HES1 in binding to these sites. HEN2 also represses Mash1 transcription but the inhibitory effects are weaker than that of HES1, and so up-regulating transcription of Mash1. (D) LMO3 promotes recruitment of HEN2 onto HES1-binding sites and E-box on Mash1 promoter by forming complex with HEN2 but inhibits negative effects of HEN2 on Mash1 promoter. Furthermore, LMO3 inhibits recruitment of HES1 onto HES1-binding sites and E-box on Mash1 promoter, and so Mash1 may be more highly expressed.
(TIF)
Correlation between expression of LMO3 or Mash1 and other prognostic factors (Student's t-test).
(PDF)
Univariate and multivariate analyses of Mash1 and LMO3 mRNA expression as well as other prognostic factors in primary neuroblastomas.
(PDF)
Acknowledgments
We are grateful to Dr. Ryuichiro Kageyama for providing the expression plasmid encoding HES1, to Drs. Nobutaka Hattori and Kaori Shiba for their valuable discussions. We also thank Yuki Nakamura for her technical assistance.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: This work was supported in part by a Grant-in-Aid from the Ministry of Health, Labour and Welfare for the Third Term Comprehensive Control Research for Cancer; a Grant-in-Aid for Scientific Research on Priority Areas from the Ministry of Education, Culture, Sports, Science and Technology, Japan; and a Grant-in-Aid for Scientific Research from Japan Society for the Promotion of Science. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Nakagawara A. Aloe L, Calzà L, editors. Neural crest development and neuroblastoma: the genetic and biological link. NGF and related molecules in health and disease. 2004. pp. 33–242. Progress in brain research. ELSEVIER. [DOI] [PubMed]
- 2.Nakagawara A, Ohira M. Comprehensive genomics linking between neural development and cancer: neuroblastoma as a model. Cancer Letters. 2004;204:213–224. doi: 10.1016/S0304-3835(03)00457-9. [DOI] [PubMed] [Google Scholar]
- 3.Bach I. The LIM domain: regulation by association. Mech Dev. 2000;91:5–17. doi: 10.1016/s0925-4773(99)00314-7. [DOI] [PubMed] [Google Scholar]
- 4.Rabbitts TH. LMO T-cell translocation oncogenes typify genes activated by chromosomal translocations that alter transcription and developmental processes. Genes Dev. 1998;12:2651–2657. doi: 10.1101/gad.12.17.2651. [DOI] [PubMed] [Google Scholar]
- 5.Visvader JE, Venter D, Hahm K, Santamaria M, Sum EY, et al. The LIM domain gene LMO4 inhibits differentiation of mammary epithelial cells in vitro and is overexpressed in breast cancer. Proc Natl Acad Sci USA. 2001;98:14452–14457. doi: 10.1073/pnas.251547698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sum EY, Segara D, Duscio B, Bath ML, Field AS, et al. Overexpression of LMO4 induces mammary hyperplasia, promotes cell invasion, and is a predictor of poor outcome in breast cancer. Proc Natl Acad Sci USA. 2005;102:7659–7664. doi: 10.1073/pnas.0502990102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Aoyama M, Ozaki T, Inuzuka H, Tomotsune D, Hirato J, et al. LMO3 interacts with neuronal transcription factor, HEN2, and acts as an oncogene in neuroblastoma. Cancer Res. 2005;65:4587–4597. doi: 10.1158/0008-5472.CAN-04-4630. [DOI] [PubMed] [Google Scholar]
- 8.Gestblom C, Grynfeld A, Ora I, Ortoft E, Larsson C, et al. The basic helix-loop-helix transcription factor dHAND, a marker gene for the developing human sympathetic nervous system, is expressed in both high- and low-stage neuroblastomas. Lab Invest. 1999;79:67–79. [PubMed] [Google Scholar]
- 9.Ichimiya S, Nimura Y, Seki N, Ozaki T, Nagase T, et al. Downregulation of hASH1 is associated with the retinoic acid-induced differentiation of human neuroblastoma cell lines. Med Pediatr Oncol. 2001;36:132–134. doi: 10.1002/1096-911X(20010101)36:1<132::AID-MPO1031>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- 10.Kageyama R, Ohtsuka T, Kobayashi T. The Hes family: repressors and oscillators that orchestrate embryogenesis. Development. 2007;134:1243–1251. doi: 10.1242/dev.000786. [DOI] [PubMed] [Google Scholar]
- 11.Ramain P, Khechumian R, Khechumian K, Arbogast N, Ackermann C, et al. Interactions between chip and the achaete/scute-daughterless heterodimers are required for pannier-driven proneural patterning. Mol Cell. 2000;6:781–790. doi: 10.1016/s1097-2765(05)00079-1. [DOI] [PubMed] [Google Scholar]
- 12.Asmar J, Biryukova I, Heitzler P. Drosophila dLMO-PA isoform acts as an early activator of achaete/scute proneural expression. Developmental biology. 2008;316:487–497. doi: 10.1016/j.ydbio.2008.01.040. [DOI] [PubMed] [Google Scholar]
- 13.Axelson H. The Notch signaling cascade in neuroblastoma: role of the basic helix-loop-helix proteins HASH-1 and HES-1. Cancer Letters. 2004;204:171–178. doi: 10.1016/S0304-3835(03)00453-1. [DOI] [PubMed] [Google Scholar]
- 14.Brown L, Baer R. HEN1 encodes a 20-kilodalton phosphoprotein that binds an extended E-box motif as a homodimer. Mol Cell Biol. 1994;14:1245–1255. doi: 10.1128/mcb.14.2.1245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wildner H, Gierl MS, Strehle M, Pla P, Birchmeier C. Insm1 (IA-1) is a crucial component of the transcriptional network that controls differentiation of the sympatho-adrenal lineage. Development. 2008;135:473–481. doi: 10.1242/dev.011783. [DOI] [PubMed] [Google Scholar]
- 16.Watt F, Watanabe R, Yang W, Agren N, Arvidsson Y, et al. A novel MASH1 enhancer with N-myc and CREB-binding sites is active in neuroblastoma. Cancer Gene Therapy. 2007;14:287–296. doi: 10.1038/sj.cgt.7701012. [DOI] [PubMed] [Google Scholar]
- 17.Bao J, Talmage DA, Role LW, Gautier J. Regulation of neurogenesis by interactions between HEN1 and neuronal LMO proteins. Development. 2000;127:425–435. doi: 10.1242/dev.127.2.425. [DOI] [PubMed] [Google Scholar]
- 18.Bagley CG, Lipkowitz S, Gobel V, Mahon KA, Bertness V, et al. Molecular characterization of NSCL, a gene encoding a helix-loop-helix protein expressed in the developing nervous system. Proc Natl Acad Sci USA. 1992;89:38–42. doi: 10.1073/pnas.89.1.38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Good DJ, Porter FD, Mahon KA, Parlow AF, Westphal H, et al. Hypogonadism and obesity in mice with a targeted deletion of the Nhlh2 gene. Nat Genet. 1997;15:397–401. doi: 10.1038/ng0497-397. [DOI] [PubMed] [Google Scholar]
- 20.Lanigan TM, DeRaad SK, Russo AF. Requirement of the MASH-1 transcription factor for neuroendocrine differentiation of thyroid C cells. J Neurobiol. 1998;34:126–134. [PubMed] [Google Scholar]
- 21.Tse E, Smith AJ, Hunt S, Lavenir I, Forster A, et al. Null mutation of the Lmo4 gene or a combined null mutation of the Lmo1/Lmo3 genes causes perinatal lethality, and Lmo4 controls neural tube development in mice. Mol Cell Biol. 2004;24:2063–2073. doi: 10.1128/MCB.24.5.2063-2073.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Brodeur GM. Neuroblastoma: biological insights into a clinical enigma. Nat Rev Cancer. 2003;3:203–216. doi: 10.1038/nrc1014. [DOI] [PubMed] [Google Scholar]
- 23.Morikawa Y, Zehir A, Maska E, Deng C, Schneider MD, et al. BMP signaling regulates sympathetic nervous system development through Smad 4-dependent and -independent pathways. Development. 2009;136:3575–3584. doi: 10.1242/dev.038133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mizuguchi H, Kay MA. Efficient construction of a recombinant adenovirus vector by an improved in vitro ligation method. Hum Gene Ther. 1998;9:2577–2583. doi: 10.1089/hum.1998.9.17-2577. [DOI] [PubMed] [Google Scholar]
- 25.Mizuguchi H, Kay MA. A simple method for constructing E1- and E1/E4-deleted recombinant adenoviral vectors. Hum Gene Ther. 1999;10:2013–2017. doi: 10.1089/10430349950017374. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Mash1-mediated growth promotion and LMO3/HEN2-mediated transcriptional induction of Mash1 in SK-N-BE cells. (A) siRNA-mediated knockdown of LMO3. SK-N-BE cells were transfected with empty plasmid (4 µg) or with expression plasmid for siRNA targeting LMO3 (4 µg). Forty-eight hours after transfection, total RNA was prepared and analyzed for expression levels of LMO3 and Mash1 by RT-PCR. (B) Decreased growth rate in LMO3-knocked down cells. SK-N-BE cells (4.5×103 cells/well, 96 well culture plate) were transfected with empty plasmid (0.2 µg) or with expression plasmid for siRNA targeting LMO3 (0.2 µg). Forty-eight hours after transfection, cells were transferred into fresh medium. At the indicated time points, cell growth was measured by MTT assay (Cell Counting Kit-8, DOJINDO). (C) RT-PCR. SK-N-BE cells were transfected with pcDNA3 empty plasmid or with the indicated combinations of expression plasmid HA-LMO3 or FLAG-HEN2. At 72 hours after transfection, total RNA was analyzed for expression levels of LMO3, HEN2 and Mash1 by RT-PCR. GAPDH was used as an internal control. (D) siRNA-mediated knockdown of LMO3 reduces the promoter activity of Mash1. SK-N-BE cells were co-transfected with constant amount of pluc-Mash1 (100 ng) and pRL-CMV (0.2 ng) in the presence or absence of increasing amounts of expression plasmid for siRNA against human LMO3 (100 or 400 ng). Forty-eight hours after transfection, cells were lysed and their luciferase activities were measured.
(TIF)
Expression of LMO3 , HEN2 , Mash1 or HES1 in neuroblastoma cell lines. Semiquantitative RT-PCR analysis for expression of LMO3, HEN3, Mash1 or HES1 in neuroblastoma cell lines is performed under linear amplification conditions. Expression of GAPDH is shown as a control.
(TIF)
Model for LMO3 and HEN2 cooperation in transcriptional regulation of Mash1 in Neuroblastoma. (A) HES1 binds to HES1 binding sites and E- box on Mash1 promoter and represses Mash1 transcription. (B) LMO3 inhibits recruitment of HES1 onto HES1-binding sites and E-box on Mash1 promoter by forming complex with HES1, and thereby inducing the expression of Mash1. (C) HEN2 interferes with recruitment of HES1 onto HES1-binding sites and E-box on Mash1 promoter by forming complex with HES1 and competing with HES1 in binding to these sites. HEN2 also represses Mash1 transcription but the inhibitory effects are weaker than that of HES1, and so up-regulating transcription of Mash1. (D) LMO3 promotes recruitment of HEN2 onto HES1-binding sites and E-box on Mash1 promoter by forming complex with HEN2 but inhibits negative effects of HEN2 on Mash1 promoter. Furthermore, LMO3 inhibits recruitment of HES1 onto HES1-binding sites and E-box on Mash1 promoter, and so Mash1 may be more highly expressed.
(TIF)
Correlation between expression of LMO3 or Mash1 and other prognostic factors (Student's t-test).
(PDF)
Univariate and multivariate analyses of Mash1 and LMO3 mRNA expression as well as other prognostic factors in primary neuroblastomas.
(PDF)