
Figure 1.
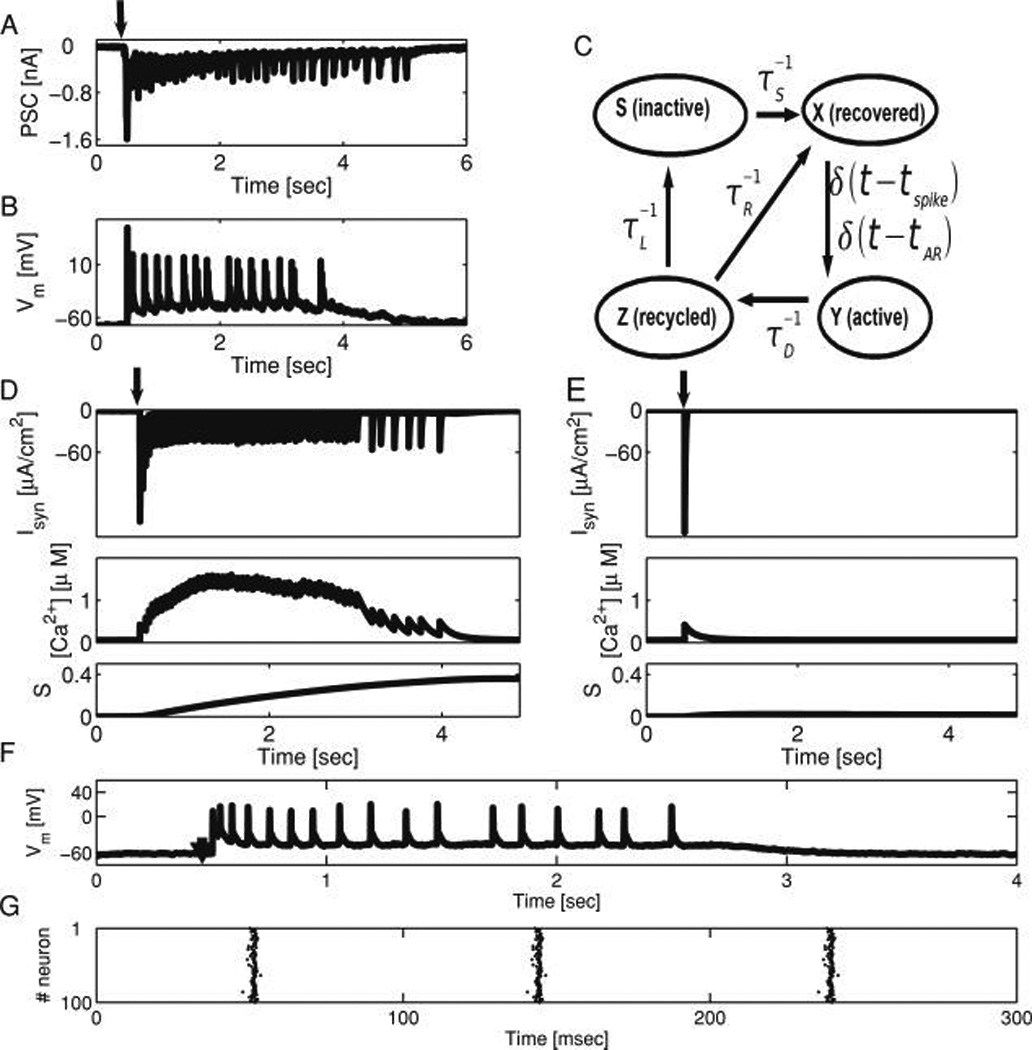
Presynaptic calcium-driven asynchronous neurotransmitter release and short-term depression underlie network reverberations. (A) Postsynaptic current recorded from a cultured hippocampal neuron in a small (≈100 neurons) network in response to a transient (2 msec) suprathreshold input delivered to another neuron (stimulation time marked with arrow). Recurrent synaptic activity can be observed in the recorded neuron at increasing intervals, lasting for seconds. (B) Membrane voltage dynamics of simultaneously recorded neuron in the same network. (C) Schematic presentation of the synaptic dynamics model used in this work, elaborated from Tsodyks et al. (2000) and described in Volman et al. (2007). Synaptic resource (excitatory neurotransmitter) traffics between four states in an activity-dependent manner: X (recovered, ready for release), Y (active, bound to postsynaptic receptors), Z (recycled, unbound or uptaken), and S (inactive, a longer route to recovery). Postsynaptic currents are generated at times tSP and tAR that describe the occurrence of action potential–driven (phasic) and asynchronous release (AR) events, correspondingly. (D, E) The rate of AR depends on presynaptic calcium concentration, which is a function of previous action potential times. The calcium dependence of the rate of AR determines whether a model network will respond to a brief stimulation with persistent reverberatory activity lasting for seconds, composed of dozens of approximately 50 ms polysynaptic current (PSC) clusters (D, upper panel, ηmax = 0.3), or a single PSC cluster with no further network activity, as in E, upper panel, ηmax = 0. Stimulation times marked with an arrow. Compare to Lau and Bi (2005) for experimental analogs of each scenario. The dynamics of presynaptic calcium (middle panels) and the slow synaptic depression variable S (lower panels) at model synaptic terminals are shown. In the case of a reverberatory response (D), recurrent network activity is terminated when a critical fraction of synaptic resource has accumulated in the inactive state (S), acting as a sink to deprive synapses of sufficient efficacy to drive the next PSC cluster. (F) In the course of model network reverberation, the neuronal membrane remains depolarized, and eachmodel neuron generates 0–1 spikes per PSC cluster, as in the experimental analog (B; Lau and Bi, 2005). (G) Raster plot of model network activity, showing that during the PSC clusters, most of the model neurons are engaged in collective, highly correlated activity, ruling out the possibility that the reverberatory response is due to the circulation of activity in the network with a return time ≈ 1/rate. Only AR, not spike-locked phasic release, can bridge the gap between PSC clusters. In all model cases shown, the probability of establishing a unidirectional connection between a pair of model neurons was p = 0.1.