
Figure 3.
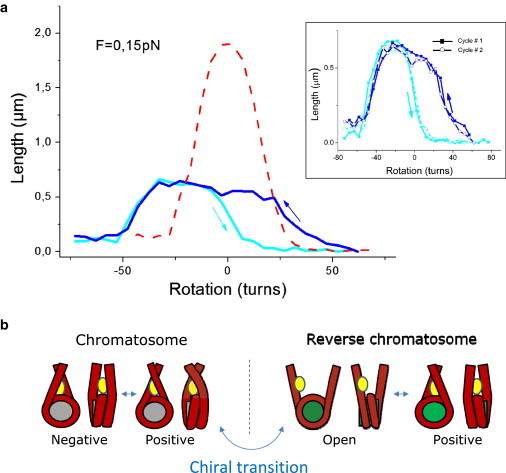
Response under topological deformation of a chromatosome fiber. (a) Extension-versus-rotation responses in buffer B0 at a constant force of 0.15 pN. (Light blue) Response of a chromatosome fiber when torsion is increased from negative to positive values. (Blue) Response of the same fiber when torsion is decreased from positive to negative values. This cycle displays the same hysteretic behavior as in the case of nucleosome fibers. It indicates that the chiral transition described previously is still possible when entry/exit DNAs are locked by linker histone. (Red) Response of the corresponding naked DNA after a treatment with heparin at a concentration of 500 μg/mL. The shift in topology after chromatosome removal is ∼−1.4 turns per particle. It reflects a different repartition of the two states of the chromatosome compared to the three states of the nucleosome and/or of their topologies. (Inset) Two successive hysteresis cycles are presented (light blue, forward curves; blue, backward curves). These two cycles are identical, thus proving that linker histones are not ejected from the fiber when submitted to an extensive positive supercoiling. (b) Models of the states accessible for the chromatosome under topological constraints. Under moderate deformation, chromatosomes can be found under the negatively or positively crossed conformation, explaining the large apex observable (light-blue curve in panel a). Extensive positive supercoiling induces the formation of reverse chromatosomes responsible for the trapping of one positive turn per particle. This value is comparable to the one measured for nucleosomes fibers. It could be a clue that reversome conformation is identical in the two cases and that linker histone does not participate in that structure. However, linker histone is not ejected during chiral transition, because the hysteresis is reproducible, this protein being still attached to the reversomes.