

Abstract
Most higher plants develop severe toxicity symptoms when grown on
ammonium (NH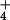 ) as the sole nitrogen source. Recently,
NH
) as the sole nitrogen source. Recently,
NH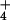 toxicity has been implicated as a cause of forest
decline and even species extinction. Although mechanisms underlying
NH
toxicity has been implicated as a cause of forest
decline and even species extinction. Although mechanisms underlying
NH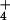 toxicity have been extensively sought, the primary
events conferring it at the cellular level are not understood. Using a
high-precision positron tracing technique, we here present a
cell-physiological characterization of NH
toxicity have been extensively sought, the primary
events conferring it at the cellular level are not understood. Using a
high-precision positron tracing technique, we here present a
cell-physiological characterization of NH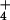 acquisition
in two major cereals, barley (Hordeum vulgare), known to
be susceptible to toxicity, and rice (Oryza sativa),
known for its exceptional tolerance to even high levels of
NH
acquisition
in two major cereals, barley (Hordeum vulgare), known to
be susceptible to toxicity, and rice (Oryza sativa),
known for its exceptional tolerance to even high levels of
NH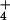 . We show that, at high external
NH
. We show that, at high external
NH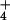 concentration
([NH
concentration
([NH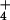 ]o), barley root cells experience a
breakdown in the regulation of NH
]o), barley root cells experience a
breakdown in the regulation of NH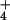 influx, leading to
the accumulation of excessive amounts of NH
influx, leading to
the accumulation of excessive amounts of NH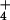 in the
cytosol. Measurements of NH
in the
cytosol. Measurements of NH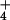 efflux, combined with a
thermodynamic analysis of the transmembrane electrochemical potential
for NH
efflux, combined with a
thermodynamic analysis of the transmembrane electrochemical potential
for NH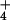 , reveal that, at elevated
[NH
, reveal that, at elevated
[NH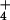 ]o, barley cells engage a
high-capacity NH
]o, barley cells engage a
high-capacity NH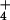 -efflux system that supports outward
NH
-efflux system that supports outward
NH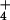 fluxes against a sizable gradient. Ammonium efflux
is shown to constitute as much as 80% of primary influx, resulting in
a never-before-documented futile cycling of nitrogen across the plasma
membrane of root cells. This futile cycling carries a high energetic
cost (we record a 40% increase in root respiration) that is
independent of N metabolism and is accompanied by a decline in growth.
In rice, by contrast, a cellular defense strategy has evolved that is
characterized by an energetically neutral, near-Nernstian,
equilibration of NH
fluxes against a sizable gradient. Ammonium efflux
is shown to constitute as much as 80% of primary influx, resulting in
a never-before-documented futile cycling of nitrogen across the plasma
membrane of root cells. This futile cycling carries a high energetic
cost (we record a 40% increase in root respiration) that is
independent of N metabolism and is accompanied by a decline in growth.
In rice, by contrast, a cellular defense strategy has evolved that is
characterized by an energetically neutral, near-Nernstian,
equilibration of NH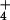 at high
[NH
at high
[NH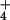 ]o. Thus our study has characterized
the primary events in NH
]o. Thus our study has characterized
the primary events in NH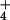 nutrition at the cellular
level that may constitute the fundamental cause of NH
nutrition at the cellular
level that may constitute the fundamental cause of NH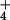 toxicity in plants.
toxicity in plants.
Plants can extract and use
various forms of nitrogen (N) from soils, most importantly the
inorganic ions ammonium (NH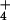 ) and nitrate
(NO
) and nitrate
(NO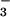 ). While one might expect NH
). While one might expect NH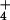 to
be preferred by plants, as its assimilation requires less energy than
that of NO
to
be preferred by plants, as its assimilation requires less energy than
that of NO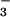 (1), only a few species perform well when
NH
(1), only a few species perform well when
NH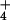 is the only, or predominant, source of N (2). By
contrast, most species develop toxicity symptoms when grown on moderate
to high levels of NH
is the only, or predominant, source of N (2). By
contrast, most species develop toxicity symptoms when grown on moderate
to high levels of NH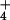 (1–6), whereas normal growth in
these species is seen on NO
(1–6), whereas normal growth in
these species is seen on NO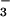 . Ammonium toxicity is
especially problematic in areas with intensive agriculture and
cultivation of livestock, where high levels of
NH3 emission, and subsequent NH
. Ammonium toxicity is
especially problematic in areas with intensive agriculture and
cultivation of livestock, where high levels of
NH3 emission, and subsequent NH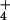 deposition, are observed (6, 7). It is estimated that N deposition from
atmospheric NH3 can reach levels as high as 50
kg⋅ha−1⋅yr−1 in
some parts of Europe, in extreme cases constituting 50–80% of total N
deposited from all possible sources (6). As a result, N saturation in
many natural ecosystems is exceeded by as much as 10-fold, and damage
to forest and agricultural crops alike has been attributed directly to
this phenomenon (8, 9). This is a problem of serious concern in both
Europe and North America (6–10).
deposition, are observed (6, 7). It is estimated that N deposition from
atmospheric NH3 can reach levels as high as 50
kg⋅ha−1⋅yr−1 in
some parts of Europe, in extreme cases constituting 50–80% of total N
deposited from all possible sources (6). As a result, N saturation in
many natural ecosystems is exceeded by as much as 10-fold, and damage
to forest and agricultural crops alike has been attributed directly to
this phenomenon (8, 9). This is a problem of serious concern in both
Europe and North America (6–10).
Several hypotheses have been advanced to explain why
NH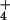 is toxic to plants, but none is considered
satisfactory (3, 5). As NH
is toxic to plants, but none is considered
satisfactory (3, 5). As NH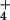 uptake mechanisms are
coupled to H+ extrusion into the rooting medium
and H+ release is also associated with
NH
uptake mechanisms are
coupled to H+ extrusion into the rooting medium
and H+ release is also associated with
NH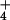 incorporation into protein (6), it has been
repeatedly suggested that root medium acidification (11) and/or
intracellular pH disturbance (3, 12) may explain the observed symptoms.
However, in many cases, toxicity is equally observed in pH-buffered
media (13), and a recent study on pea (Pisum sativum), known
to be ammonium sensitive, has discounted the occurrence of
ammonium-induced cytosolic pH disturbance (12). Others have suggested
that carbohydrate limitation may contribute to the toxicity syndrome,
based on the finding that NH
incorporation into protein (6), it has been
repeatedly suggested that root medium acidification (11) and/or
intracellular pH disturbance (3, 12) may explain the observed symptoms.
However, in many cases, toxicity is equally observed in pH-buffered
media (13), and a recent study on pea (Pisum sativum), known
to be ammonium sensitive, has discounted the occurrence of
ammonium-induced cytosolic pH disturbance (12). Others have suggested
that carbohydrate limitation may contribute to the toxicity syndrome,
based on the finding that NH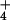 per se is not
translocated to the shoot in most plants (14), and, thus, all C
skeletons for N assimilation must be provided in roots, causing local C
deprivation (15). In some cases, external provision of
α-ketoglutarate, a key carbon source for N assimilation, alleviated
toxicity symptoms (4), but in other cases it failed to enhance
NH
per se is not
translocated to the shoot in most plants (14), and, thus, all C
skeletons for N assimilation must be provided in roots, causing local C
deprivation (15). In some cases, external provision of
α-ketoglutarate, a key carbon source for N assimilation, alleviated
toxicity symptoms (4), but in other cases it failed to enhance
NH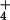 metabolism (16), suggesting that other factors may
limit NH
metabolism (16), suggesting that other factors may
limit NH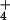 assimilation. The hypothesis that
NH
assimilation. The hypothesis that
NH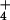 toxicity results from the uncoupling of
photophosphorylation in chloroplasts (6) has long been shown to be
incorrect, as even very large NH
toxicity results from the uncoupling of
photophosphorylation in chloroplasts (6) has long been shown to be
incorrect, as even very large NH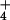 concentrations do
not affect this process in intact chloroplasts (17). A diminution of
essential cations such as K+,
Mg2+, and Ca2+ in tissues
of plants grown under prolonged exposure to NH
concentrations do
not affect this process in intact chloroplasts (17). A diminution of
essential cations such as K+,
Mg2+, and Ca2+ in tissues
of plants grown under prolonged exposure to NH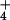 has
been widely documented (18–20), and while this may be a contributor to
the toxicity syndrome, it is clearly a longer-term effect, resulting
from nutrient imbalance.
has
been widely documented (18–20), and while this may be a contributor to
the toxicity syndrome, it is clearly a longer-term effect, resulting
from nutrient imbalance.
What these hypotheses have in common is that they are all contingent
upon the permeation of NH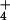 into the cell. Therefore,
the study of transmembrane NH
into the cell. Therefore,
the study of transmembrane NH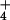 fluxes is essential to
the establishment of a proper context for the discussion of the issue
of toxicity. Surprisingly, however, no study of the primary events of
NH
fluxes is essential to
the establishment of a proper context for the discussion of the issue
of toxicity. Surprisingly, however, no study of the primary events of
NH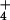 acquisition, including the modes of entry,
metabolism, and sequestration, has been undertaken in this context.
Here we have used positron-emission tracing with the short-lived
nitrogen isotope 13N to characterize these
primary events at the cellular level, in the model grass species rice
(Oryza sativa) and barley (Hordeum vulgare).
acquisition, including the modes of entry,
metabolism, and sequestration, has been undertaken in this context.
Here we have used positron-emission tracing with the short-lived
nitrogen isotope 13N to characterize these
primary events at the cellular level, in the model grass species rice
(Oryza sativa) and barley (Hordeum vulgare).
Methods
Plant Culture and Radiotracer Experiments.
Seedlings of barley (cv. CM-72) and of two major tropical-wetland
varieties of rice (cv. IR-72 and cv. M-202) were cultivated
hydroponically for 7 d and 21 d, respectively, in
controlled-environment chambers. Hydroponic tanks contained aerated and
N-free modified Johnson's solution (2, 14, 16). NH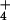 was supplied as
(NH4)2SO4
at steady-state concentrations of 0.1 mM or 10 mM during growth and for
radiolabeling experiments, which were conducted in the growth chambers.
The short-lived radiotracer 13N (half-life =
9.98 min) was used to trace NH
was supplied as
(NH4)2SO4
at steady-state concentrations of 0.1 mM or 10 mM during growth and for
radiolabeling experiments, which were conducted in the growth chambers.
The short-lived radiotracer 13N (half-life =
9.98 min) was used to trace NH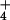 fluxes and to estimate
cytosolic concentrations of NH
fluxes and to estimate
cytosolic concentrations of NH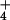 by compartmental
analysis, as described previously (2, 16, 21). In brief, roots of
intact seedlings were exposed to growth solution spiked with
13NH
by compartmental
analysis, as described previously (2, 16, 21). In brief, roots of
intact seedlings were exposed to growth solution spiked with
13NH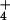 for 1 h, to maximize
the specific activity of tracer in the cytosolic compartment of root
cells (21). Seedlings were then transferred to efflux funnels (16, 21),
and tracer from the roots was eluted successively with 20-ml aliquots
of nonlabeled solution for various washout periods. With
t = 0 as the time of transfer from loading to washing
solution and tfinal = 24 min for the
final elution, the time periods for the 25 successive washes were:
10 s (three times), 15 s (six times), 30 s (four times),
1 min (four times), and 2 min (eight times). γ-Ray emissions in the
eluates were then measured (see Fig. 1). After the final elution, roots
were excised from shoots, spin-dried for 30 s, and weighed, and
the γ-ray emissions of roots and shoots were counted. Influx, efflux,
net flux, and cytosolic concentrations of NH
for 1 h, to maximize
the specific activity of tracer in the cytosolic compartment of root
cells (21). Seedlings were then transferred to efflux funnels (16, 21),
and tracer from the roots was eluted successively with 20-ml aliquots
of nonlabeled solution for various washout periods. With
t = 0 as the time of transfer from loading to washing
solution and tfinal = 24 min for the
final elution, the time periods for the 25 successive washes were:
10 s (three times), 15 s (six times), 30 s (four times),
1 min (four times), and 2 min (eight times). γ-Ray emissions in the
eluates were then measured (see Fig. 1). After the final elution, roots
were excised from shoots, spin-dried for 30 s, and weighed, and
the γ-ray emissions of roots and shoots were counted. Influx, efflux,
net flux, and cytosolic concentrations of NH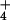 were
estimated from analysis of the wash-out kinetics as described in detail
elsewhere (2, 16, 21). All fluxes are expressed in μmol per g (fresh
weight) per h. Symbols, and basic calculation methods, for fluxes are
as follows:
were
estimated from analysis of the wash-out kinetics as described in detail
elsewhere (2, 16, 21). All fluxes are expressed in μmol per g (fresh
weight) per h. Symbols, and basic calculation methods, for fluxes are
as follows:
Figure 1.

Semilogarithmic plots of 13NH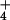 efflux from
the cytosolic compartments of barley and rice roots. Plants were
prelabeled under steady-state conditions, with NH
efflux from
the cytosolic compartments of barley and rice roots. Plants were
prelabeled under steady-state conditions, with NH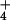 provided externally at 10 mM. Plots have been corrected for specific
activities of 13N tracer (2, 21), allowing a direct
comparison of initial efflux rates by inspection of the
y-intercepts of the regression lines.
provided externally at 10 mM. Plots have been corrected for specific
activities of 13N tracer (2, 21), allowing a direct
comparison of initial efflux rates by inspection of the
y-intercepts of the regression lines.
Φco = efflux from the cytosol, obtained from the rate of 13N release from the cytosol at time 0.
Φnet = net flux, obtained directly from the accumulation of 13N in the plants at the end of the elution period.
Φoc = unidirectional influx, calculated from Φnet + Φco.
Φxylem = xylem flux of 13N, obtained directly from accumulation in the shoot, at the end of the elution period.
Φmet/vac = combined flux to assimilation and vacuole, resulting from Φnet − Φxylem.
Cytosolic NH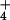 concentrations were calculated from the
quotient of the rate of 13NH
concentrations were calculated from the
quotient of the rate of 13NH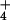 release integrated over 5 times the half-life of cytosolic
NH
release integrated over 5 times the half-life of cytosolic
NH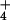 exchange, and the ratio of efflux to all fluxes
removing 13NH
exchange, and the ratio of efflux to all fluxes
removing 13NH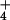 from the cytosol,
and were based on the assumption that the cytosol occupies 5% of cell
volume (2, 16, 21). Experiments were repeated four to five times.
Standard errors (SE) for fluxes were less than 20% of the means.
from the cytosol,
and were based on the assumption that the cytosol occupies 5% of cell
volume (2, 16, 21). Experiments were repeated four to five times.
Standard errors (SE) for fluxes were less than 20% of the means.
Electrophysiological Measurements.
Membrane potential differences were measured as described elsewhere
(22). Intact barley or rice roots were held between steel pins and
pieces of silicone tubing in a Plexiglas chamber affixed to a light
microscope, and they were impaled with glass microelectrodes until a
stable electrical potential difference between root and external
solution was maintained for at least 2 min. Sixty-seven measurements in
total were made on different plants for the 0.1 mM NH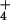 condition in barley, and 20 for the 10 mM NH
condition in barley, and 20 for the 10 mM NH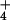 condition. SE was <3% of the means. For rice, 17 measurements were
undertaken at 0.1 mM external NH
condition. SE was <3% of the means. For rice, 17 measurements were
undertaken at 0.1 mM external NH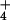 concentration
([NH
concentration
([NH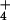 ]o), and 26 at 10 mM
[NH
]o), and 26 at 10 mM
[NH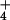 ]o (SE < 5% of the
means).
]o (SE < 5% of the
means).
Respiration Measurements.
O2 depletion was followed potentiometrically in intact roots of barley and rice with a Hansatech cuvette/electrode system attached to a chart recorder. Cuvettes were filled with air-saturated growth solutions. Calculations were based on slopes of steady O2 depletion lines generated over 3–10 min, after which plants were weighed and discarded. The O2 content of the cuvette was not reduced by more than 25% of the initial value. Measurements were repeated 20–25 times.
Relative Growth Rates.
Individual barley plants were monitored over days 1, 3, and 7 after
germination. Surface water was removed from roots before weighing,
after which the plants were immediately returned to solution. Eight
replicates were used for each of the two steady-state
NH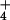 -growth concentrations.
-growth concentrations.
Results and Discussion
In the present study, we assayed NH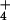 transport
properties in the root cells of two major cereal species known to
differ markedly in their abilities to tolerate NH
transport
properties in the root cells of two major cereal species known to
differ markedly in their abilities to tolerate NH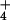 as
the sole N source. 13NH
as
the sole N source. 13NH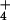 efflux
from roots of intact plants of the two species, prelabeled with this
tracer, showed substantial differences in the rates of
NH
efflux
from roots of intact plants of the two species, prelabeled with this
tracer, showed substantial differences in the rates of
NH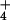 extrusion from root cells to the external medium
(Fig. 1). Interestingly, barley
(Hordeum vulgare), which suffers from NH
extrusion from root cells to the external medium
(Fig. 1). Interestingly, barley
(Hordeum vulgare), which suffers from NH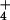 toxicity (23), excretes NH
toxicity (23), excretes NH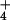 at a significantly higher
rate than rice (Oryza sativa), a species whose tolerance to
NH
at a significantly higher
rate than rice (Oryza sativa), a species whose tolerance to
NH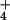 is considered exceptional (24). Fig.
2, depicting bidirectional chemical
fluxes of NH
is considered exceptional (24). Fig.
2, depicting bidirectional chemical
fluxes of NH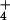 across the plasma membranes of these two
species, documents in both cases a large increase in total flux
(equivalent to unidirectional influx) as the steady-state external
NH
across the plasma membranes of these two
species, documents in both cases a large increase in total flux
(equivalent to unidirectional influx) as the steady-state external
NH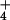 concentration is stepped up from a moderate level
of 0.1 mM to a level of 10 mM, which represents the high end of
NH
concentration is stepped up from a moderate level
of 0.1 mM to a level of 10 mM, which represents the high end of
NH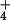 concentrations found in agricultural soils
(1–11), and which is potentially toxic (24). Importantly, however, the
increase in barley is much higher (12-fold) than in rice (7-fold). In
fact, the magnitudes of the fluxes in barley are the highest ever
reported for NH
concentrations found in agricultural soils
(1–11), and which is potentially toxic (24). Importantly, however, the
increase in barley is much higher (12-fold) than in rice (7-fold). In
fact, the magnitudes of the fluxes in barley are the highest ever
reported for NH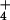 in a plant root system, and strongly
suggest the operation of channel-based NH
in a plant root system, and strongly
suggest the operation of channel-based NH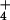 -transport
systems, whose existence has been demonstrated by several recent
studies (25, 26). As the steady-state net fluxes are almost identical
in the two species, a substantially larger fraction of incoming
nitrogen is subsequently excreted in the ammonium-sensitive species.
This pronounced excretion of N is inextricably linked to an apparent
inability of barley to exclude NH
-transport
systems, whose existence has been demonstrated by several recent
studies (25, 26). As the steady-state net fluxes are almost identical
in the two species, a substantially larger fraction of incoming
nitrogen is subsequently excreted in the ammonium-sensitive species.
This pronounced excretion of N is inextricably linked to an apparent
inability of barley to exclude NH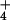 at the more
primary, intake, step. Our preliminary analyses on other species known
to suffer NH
at the more
primary, intake, step. Our preliminary analyses on other species known
to suffer NH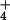 toxicity, including wheat, tomato, and
the tree species Douglas-fir and trembling aspen (27), suggest that
this phenomenon is by no means isolated, but may well occur universally
among susceptible higher plants.
toxicity, including wheat, tomato, and
the tree species Douglas-fir and trembling aspen (27), suggest that
this phenomenon is by no means isolated, but may well occur universally
among susceptible higher plants.
Figure 2.

Comparison of steady-state bidirectional plasma-membrane
NH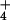 fluxes in barley and rice roots at 0.1 mM and 10
mM [NH
fluxes in barley and rice roots at 0.1 mM and 10
mM [NH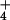 ]o. Column height represents
total influx from the external medium (Φoc), while the
filled areas depict the portion of influx returned to the environment
by efflux transport (Φco). The net flux
(Φnet) is the difference between these two fluxes.
Vertical bars indicate standard errors of influx means.
]o. Column height represents
total influx from the external medium (Φoc), while the
filled areas depict the portion of influx returned to the environment
by efflux transport (Φco). The net flux
(Φnet) is the difference between these two fluxes.
Vertical bars indicate standard errors of influx means.
Knowledge of the kinetic constants of NH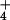 exchange for
the cytosolic component in the experimental root system, and of the
tracer content in this pool at the onset of the elution protocol,
allowed us to compare the concentrations of NH
exchange for
the cytosolic component in the experimental root system, and of the
tracer content in this pool at the onset of the elution protocol,
allowed us to compare the concentrations of NH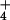 on
either side of the plasma membrane (2), and hence to investigate the
energetics of fluxes across this membrane. Our analysis shows that, at
10 mM [NH
on
either side of the plasma membrane (2), and hence to investigate the
energetics of fluxes across this membrane. Our analysis shows that, at
10 mM [NH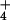 ]o, the cytosolic
concentrations of NH
]o, the cytosolic
concentrations of NH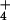 in both barley and rice are of a
level higher than ever reported in plant cells (Table
1), and indeed are irreconcilable with
the widely held dogma that no appreciable quantities of free
NH
in both barley and rice are of a
level higher than ever reported in plant cells (Table
1), and indeed are irreconcilable with
the widely held dogma that no appreciable quantities of free
NH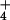 can exist in the cytosol (1, 5, 6). Nevertheless,
a thermodynamic analysis, using measurements of electrical potential
(ΔΨ) across the plasma membrane, reveals that, in barley, the
cytosolic levels of NH
can exist in the cytosol (1, 5, 6). Nevertheless,
a thermodynamic analysis, using measurements of electrical potential
(ΔΨ) across the plasma membrane, reveals that, in barley, the
cytosolic levels of NH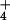 at 10 mM
[NH
at 10 mM
[NH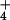 ]o are substantially lower
than predicted by the Nernst equation, which applies when
NH
]o are substantially lower
than predicted by the Nernst equation, which applies when
NH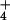 is passively distributed across a highly permeable
membrane (Table 1). Rice, by contrast, maintains a lower internal
concentration than barley under this condition, but because of the
relatively depolarized state of its plasma membrane under high
NH
is passively distributed across a highly permeable
membrane (Table 1). Rice, by contrast, maintains a lower internal
concentration than barley under this condition, but because of the
relatively depolarized state of its plasma membrane under high
NH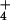 provision (22), the transmembrane distribution
ratio of NH
provision (22), the transmembrane distribution
ratio of NH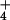 very closely approximates that predicted
by Nernstian analysis (Table 1). This lowering of membrane polarization
with increasing NH
very closely approximates that predicted
by Nernstian analysis (Table 1). This lowering of membrane polarization
with increasing NH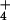 provision, which is not followed
by a restoration of that polarization in the steady state, is confirmed
by previously published work on rice (ref. 22; see Table 1) and the
aquatic NH
provision, which is not followed
by a restoration of that polarization in the steady state, is confirmed
by previously published work on rice (ref. 22; see Table 1) and the
aquatic NH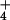 -specialist plant Lemna gibba
(28). Table 1 shows that, in rice, a depolarized membrane potential is
already achieved at 1 mM
[NH
-specialist plant Lemna gibba
(28). Table 1 shows that, in rice, a depolarized membrane potential is
already achieved at 1 mM
[NH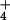 ]o, a more modest
NH
]o, a more modest
NH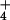 concentration. The reduction of ΔΨ with
increasing [NH
concentration. The reduction of ΔΨ with
increasing [NH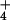 ]o apparent in
rice has the important biophysical consequence of lowering the ceiling
for NH
]o apparent in
rice has the important biophysical consequence of lowering the ceiling
for NH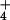 accumulation in the cytosol, thus eliminating
the gradient against which efflux transporters must work to remove
excess cytosolic NH
accumulation in the cytosol, thus eliminating
the gradient against which efflux transporters must work to remove
excess cytosolic NH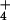 . The failure of barley to
down-regulate ΔΨ, on the other hand, sustains a gradient for very
large inward NH
. The failure of barley to
down-regulate ΔΨ, on the other hand, sustains a gradient for very
large inward NH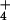 fluxes, which we in fact observe
(Fig. 2). Homeostatic restoration of ΔΨ to preset values has been
previously reported for barley exposed to various concentrations of
NO
fluxes, which we in fact observe
(Fig. 2). Homeostatic restoration of ΔΨ to preset values has been
previously reported for barley exposed to various concentrations of
NO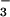 (29), the second major N source used by plants.
Under NH
(29), the second major N source used by plants.
Under NH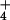 nutrition, an undiminished ΔΨ in barley
yields a high potential for NH
nutrition, an undiminished ΔΨ in barley
yields a high potential for NH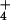 accumulation, and a
significantly larger demand on the efflux process (Table 1). Our data
show that, whereas the net flux of NH
accumulation, and a
significantly larger demand on the efflux process (Table 1). Our data
show that, whereas the net flux of NH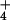 is nearly
identical in both species at 10 mM, the efflux process in barley
mediates a 2- to 3-fold higher removal of NH
is nearly
identical in both species at 10 mM, the efflux process in barley
mediates a 2- to 3-fold higher removal of NH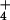 from the
cytosol, effectively reducing a Nernstian NH
from the
cytosol, effectively reducing a Nernstian NH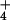 concentration of 1.32 M by the equivalent of 962 mM (Table 1).
concentration of 1.32 M by the equivalent of 962 mM (Table 1).
Table 1.
Thermodynamic analysis of cytosolic NH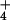 pool sizes in barley and rice
pool sizes in barley and rice
| Plant | [NH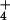 ]o, mM ]o, mM |
Influx, μmol⋅g−1⋅h−1 | Efflux, μmol⋅g−1⋅h−1 | Flux ratio | ΔΨ, mV | [NH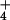 ]c, mM ]c, mM
|
|
|---|---|---|---|---|---|---|---|
| Predicted | Measured | ||||||
| Barley | 0.1 | 5.46 | 1.53 | 0.28 | −121 | 12 | 28 |
| 10 | 65.1 | 49.4 | 0.76 | −123 | 1,320 | 358 | |
| Rice | 0.1 | 4.85 | 0.67 | 0.14 | −132 | 15.6 | 33.4 |
| 5.97 (ref. 34) | 1.17 (ref. 34) | 0.19 | −122 (ref. 22) | 10.7 | 20.6 (ref. 34) | ||
| 1 | 10.51 (ref. 34) | 3.09 (ref. 34) | 0.29 | −88 (ref. 22) | 28.78 | 38.1 (ref. 34) | |
| 10 | 35.0 | 18.6 | 0.53 | −87.9 | 289 | 232 | |
| −82 (ref. 22) | 233 | ND | |||||
The Nernst equation {ΨN =
RT/(nF)
ln([NH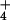 ]o/[NH
]o/[NH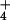 ]c),
where ΨN = the Nernst potential; R =
8.3144 J⋅K−1⋅mol−1; F = 96,485
C⋅mol−1; n = 1; and T
= 293.15 K (for barley) or 303.15 K (for rice)} was used to predict
equilibrium cytosolic concentrations
([NH
]c),
where ΨN = the Nernst potential; R =
8.3144 J⋅K−1⋅mol−1; F = 96,485
C⋅mol−1; n = 1; and T
= 293.15 K (for barley) or 303.15 K (for rice)} was used to predict
equilibrium cytosolic concentrations
([NH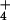 ]c) on the basis of external
concentration, membrane potential (ΔΨ), and an ambient temperature
of 20°C for barley and 30°C for rice. Cytosolic concentrations were
measured by using compartmental analysis (see text). Additional
membrane potential readings and estimates of cytosolic
NH
]c) on the basis of external
concentration, membrane potential (ΔΨ), and an ambient temperature
of 20°C for barley and 30°C for rice. Cytosolic concentrations were
measured by using compartmental analysis (see text). Additional
membrane potential readings and estimates of cytosolic
NH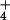 concentrations for rice (cv. M-202) are included
on the basis of analysis of previously published work (22, 34). ND, not
determined.
concentrations for rice (cv. M-202) are included
on the basis of analysis of previously published work (22, 34). ND, not
determined.
Such a process must carry a substantial energetic burden, and this
prediction was consistent with measurements of oxygen consumption in
roots of intact barley plants under the steady-state conditions of
NH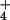 supply corresponding to those of flux
measurements. As indicated in Fig. 3,
upon transition from low steady-state NH
supply corresponding to those of flux
measurements. As indicated in Fig. 3,
upon transition from low steady-state NH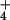 provision to
high, respiration in barley increased by a startling 41%, whereas no
significant difference was found in rice between growth conditions (not
shown). To isolate, in barley, the flux processes at the plasma
membrane level from other potentially energy-requiring pathways of
NH
provision to
high, respiration in barley increased by a startling 41%, whereas no
significant difference was found in rice between growth conditions (not
shown). To isolate, in barley, the flux processes at the plasma
membrane level from other potentially energy-requiring pathways of
NH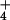 processing, we applied methionine sulfoximine
(MSX), a compound known to completely block NH
processing, we applied methionine sulfoximine
(MSX), a compound known to completely block NH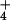 assimilation in the species under investigation (14, 16, 30).
Importantly, MSX did not diminish respiratory activity associated with
elevated external NH
assimilation in the species under investigation (14, 16, 30).
Importantly, MSX did not diminish respiratory activity associated with
elevated external NH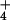 (Fig. 3), emphasizing that the
respiratory stimulation was attributable to processes upstream of
NH
(Fig. 3), emphasizing that the
respiratory stimulation was attributable to processes upstream of
NH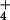 metabolism—i.e., residing at the level of
membrane transport—and independent of activities of glutamine
synthetase and other processes related to N metabolism. Other work on
barley has shown that the proportion of total respiration assigned to
processes associated with NH
metabolism—i.e., residing at the level of
membrane transport—and independent of activities of glutamine
synthetase and other processes related to N metabolism. Other work on
barley has shown that the proportion of total respiration assigned to
processes associated with NH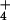 uptake and assimilation
constitutes as much as 14% of the plant's total respiratory
expenditure at 0.1 mM external NH
uptake and assimilation
constitutes as much as 14% of the plant's total respiratory
expenditure at 0.1 mM external NH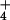 provision (31).
Most of this expenditure appears to be due to plasma-membrane transport
activity, hence we conclude that the 41% increase in total root
respiration observed in the present study for barley at 10 mM
NH
provision (31).
Most of this expenditure appears to be due to plasma-membrane transport
activity, hence we conclude that the 41% increase in total root
respiration observed in the present study for barley at 10 mM
NH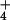 can be attributed directly to the approximately
30-fold increase in NH
can be attributed directly to the approximately
30-fold increase in NH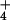 efflux from the cytosol under
this condition. Work with the NH
efflux from the cytosol under
this condition. Work with the NH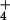 analogue
methylammonium shows that a respiratory increase is also observed in
barley (32), as well as other species (33), on addition of this
nonmetabolized compound. In the absence of metabolic sinks, it becomes
clear that membrane processes per se can impose a
substantial energetic burden on the plant root system. Indeed, in
barley the increase in respiration at 10 mM external
NH
analogue
methylammonium shows that a respiratory increase is also observed in
barley (32), as well as other species (33), on addition of this
nonmetabolized compound. In the absence of metabolic sinks, it becomes
clear that membrane processes per se can impose a
substantial energetic burden on the plant root system. Indeed, in
barley the increase in respiration at 10 mM external
NH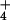 is not correlated with an increase in
growth-related energy demands; on the contrary, it accompanies a
depression in relative growth rate at the whole plant level (Fig. 3
Inset) not seen in ammonium-tolerant species such as rice
(2, 14, 21, 22, 34, 35). It is intriguing that in rice, where no
respiratory increase was found with increasing NH
is not correlated with an increase in
growth-related energy demands; on the contrary, it accompanies a
depression in relative growth rate at the whole plant level (Fig. 3
Inset) not seen in ammonium-tolerant species such as rice
(2, 14, 21, 22, 34, 35). It is intriguing that in rice, where no
respiratory increase was found with increasing NH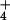 provision (data not shown), a thermodynamic equilibrium is achieved
across the plasma membrane (Table 1), suggesting that passive inwardly
and outwardly directed NH
provision (data not shown), a thermodynamic equilibrium is achieved
across the plasma membrane (Table 1), suggesting that passive inwardly
and outwardly directed NH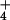 channel activities may
mediate NH
channel activities may
mediate NH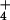 distribution across this membrane without
energetic cost. These findings underscore the cellular adaptations rice
has evolved to use NH
distribution across this membrane without
energetic cost. These findings underscore the cellular adaptations rice
has evolved to use NH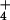 as an N source (34, 35), which
allow it to thrive in flooded soil environments where hypoxic to anoxic
conditions typically render NH
as an N source (34, 35), which
allow it to thrive in flooded soil environments where hypoxic to anoxic
conditions typically render NH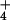 the only major N
source available to plant growth (35).
the only major N
source available to plant growth (35).
Figure 3.

Respiration rates of intact barley roots at 0.1 mM and 10 mM
[NH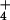 ]o.
In one experiment, 1 mM methionine sulfoximine (MSX) was applied to
block NH
]o.
In one experiment, 1 mM methionine sulfoximine (MSX) was applied to
block NH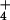 metabolism (see text). Relative growth rates
under the two conditions are shown in the Inset. Rice
experienced no significant difference in respiration under the two
NH
metabolism (see text). Relative growth rates
under the two conditions are shown in the Inset. Rice
experienced no significant difference in respiration under the two
NH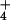 regimes (see text).
regimes (see text).
We propose that the inability of barley, and that of other species
susceptible to ammonium toxicity, to exclude NH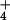 by
regulation of plasma-membrane influx systems constitutes a fundamental
breakdown in plant cell function and must precede any intracellular
toxicity-associated events such as cation displacement or carbohydrate
depletion (see above). The operation of the energy-intensive
NH
by
regulation of plasma-membrane influx systems constitutes a fundamental
breakdown in plant cell function and must precede any intracellular
toxicity-associated events such as cation displacement or carbohydrate
depletion (see above). The operation of the energy-intensive
NH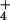 extrusion mechanism we describe appears to be
central to the ammonium toxicity syndrome, and is similar in principle
to mechanisms evolved by bacteria and carcinomas to actively excrete
cytotoxins such as antibiotics and chemotherapeutic agents (36, 37).
The ecological significance of this syndrome is substantial, not only
because soil nitrogen profiles profoundly determine the spatial and
temporal dynamics of ecosystems (2, 38), but especially in the light of
recent interventions by humans in the functioning of the global
nitrogen cycle (5–7, 39).
extrusion mechanism we describe appears to be
central to the ammonium toxicity syndrome, and is similar in principle
to mechanisms evolved by bacteria and carcinomas to actively excrete
cytotoxins such as antibiotics and chemotherapeutic agents (36, 37).
The ecological significance of this syndrome is substantial, not only
because soil nitrogen profiles profoundly determine the spatial and
temporal dynamics of ecosystems (2, 38), but especially in the light of
recent interventions by humans in the functioning of the global
nitrogen cycle (5–7, 39).
Acknowledgments
We thank M. Adam, T. Hurtado, and T. Ruth at the particle accelerator facility TRIUMF on the University of British Columbia campus for providing 13N, R. C. Huffaker for providing barley seeds, the International Rice Research Institute (IRRI) for providing rice seeds, and G. Kirk, M. Sheriff, and S. Silim for assistance with experiments and for helpful discussion. The work was supported by grants from the Natural Sciences and Engineering Research Council of Canada (NSERC).
Abbreviation
- [NH
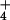 ]o
]o
external NH
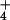 concentration
concentration
References
- 1.Reisenauer H M. In: Nitrogen in the Environment. Nielsen D R, Macdonald J G, editors. New York: Academic; 1978. pp. 157–189. [Google Scholar]
- 2.Kronzucker H J, Siddiqi M Y, Glass A D M. Nature (London) 1997;385:59–61. [Google Scholar]
- 3.Gerendas J, Zhu Z, Bendixen R, Ratcliffe R G, Sattelmacher B. Z Pflanzenernähr Bodenkd. 1997;160:239–251. [Google Scholar]
- 4.Magalhaes J R, Huber D M, Tsai C Y. Plant Sci. 1992;85:135–141. [Google Scholar]
- 5.De Graaf M C C, Bobbink R, Roelofs J G M, Verbeek P J M. Plant Ecol. 1998;135:185–196. [Google Scholar]
- 6.Pearson J, Stewart G R. New Phytol. 1993;125:283–305. doi: 10.1111/j.1469-8137.1993.tb03882.x. [DOI] [PubMed] [Google Scholar]
- 7.Fangmeier A, Hadwiger-Fangmeier A, Van der Eerden L, Jäger H-J. Environ Poll. 1994;86:43–82. doi: 10.1016/0269-7491(94)90008-6. [DOI] [PubMed] [Google Scholar]
- 8.Van Breeman N, Burrough P A, Velthorst E J, van Dobben H F, de Wit T, Ridder T B, Reijnders H F R. Nature (London) 1982;299:548–550. [Google Scholar]
- 9.Van Breeman N, Van Dijk H F G. Environ Poll. 1988;54:249–274. doi: 10.1016/0269-7491(88)90115-7. [DOI] [PubMed] [Google Scholar]
- 10.Padgett P E, Allen E B, Bytnerowicz A, Minich R A. Environ Poll. 1999;104:179–187. [Google Scholar]
- 11.Barker A V, Volk R J, Jackson W A. Plant Physiol. 1966;41:1193–1199. doi: 10.1104/pp.41.7.1193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bligny R, Gout E, Kaiser W, Heber U, Walker D, Douce R. Biochim Biophys Acta. 1997;1320:142–152. [Google Scholar]
- 13.Blacquière T, Voortman E, Stulen I. Plant Soil. 1998;106:23–34. [Google Scholar]
- 14.Kronzucker H J, Schjoerring J K, Erner Y, Kirk G J D, Siddiqi M Y, Glass A D M. Plant Cell Physiol. 1998;39:1287–1293. [Google Scholar]
- 15.Lewis O A M, James D M, Hewitt E J. Ann Bot. 1982;49:39–49. [Google Scholar]
- 16.Kronzucker H J, Siddiqi M Y, Glass A D M. Plant Physiol. 1995;109:481–490. doi: 10.1104/pp.109.2.481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Heber U. In: in Advances in Photosynthesis Research. Sybesma C, editor. The Hague, The Netherlands: Nijhoff/Junk; 1984. pp. 381–389. [Google Scholar]
- 18.Holldampf B, Barker A V. Commun Soil Sci Plant Anal. 1993;24:1945–1957. [Google Scholar]
- 19.Cao W, Tibbits T W. J Plant Nutr. 1993;16:1691–1704. doi: 10.1080/01904169309364643. [DOI] [PubMed] [Google Scholar]
- 20.Speer M, Kaiser W M. Plant Cell Environ. 1994;17:1223–1231. [Google Scholar]
- 21.Kronzucker H J, Glass A D M, Siddiqi M Y, Kirk G J D. New Phytol. 2000;145:471–476. doi: 10.1046/j.1469-8137.2000.00606.x. [DOI] [PubMed] [Google Scholar]
- 22.Wang M Y, Glass A D M, Shaff J E, Kochian L V. Plant Physiol. 1994;104:899–906. doi: 10.1104/pp.104.3.899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lewis O A M, Soares M I M, Lips S H. In: Fundamental, Ecological and Agricultural Aspects of Nitrogen Metabolism in Higher Plants, Developments in Plant and Soil Sciences. Lambers H, Neeteson J J, Stulen I, editors. Vol. 19. Dordrecht, The Netherlands: Nijhoff; 1986. pp. 295–300. [Google Scholar]
- 24.Magalhaes J R, Huber D M. Fert Res. 1989;21:1–6. [Google Scholar]
- 25.Tyerman S D, Whitehead L F, Day D A. Nature (London) 1995;378:629–632. [Google Scholar]
- 26.Nielsen K H, Schjoerring J K. Plant Physiol. 1998;118:1361–1368. doi: 10.1104/pp.118.4.1361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Min X J, Siddiqi M Y, Guy R D, Glass A D M, Kronzucker H J. Plant Cell Environ. 1999;22:821–830. [Google Scholar]
- 28.Ullrich W R, Larsson M, Larsson C-M, Lesch S, Novacky A. Physiol Plant. 1984;61:369–376. [Google Scholar]
- 29.Glass A D M, Shaff J, Kochian L V. Plant Physiol. 1992;99:456–463. doi: 10.1104/pp.99.2.456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wieneke J, Roeb G W. Z Pflanzenernähr Bodenkd. 1998;161:1–7. [Google Scholar]
- 31.Bloom A J, Sukrapanna S S, Warner R L. Plant Physiol. 1992;99:1294–1301. doi: 10.1104/pp.99.4.1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Rigano C, Di Martino Rigano V, Vona V, Carfagna S, Carillo P, Esposito S. New Phytol. 1996;132:375–382. doi: 10.1111/j.1469-8137.1996.tb01857.x. [DOI] [PubMed] [Google Scholar]
- 33.Larson T R, Rees T A V. Planta. 1994;195:195–200. [Google Scholar]
- 34.Wang M Y, Siddiqi M Y, Ruth T J, Glass A D M. Plant Physiol. 1993;103:1249–1258. doi: 10.1104/pp.103.4.1249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kronzucker H J, Kirk G J D, Siddiqi M Y, Glass A D M. Plant Physiol. 1998;116:581–587. doi: 10.1104/pp.116.2.581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.McMurry L, Petrucci R E, Jr, Levy S B. Proc Natl Acad Sci USA. 1980;77:3974–3977. doi: 10.1073/pnas.77.7.3974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nikaido H. Science. 1994;264:382–388. doi: 10.1126/science.8153625. [DOI] [PubMed] [Google Scholar]
- 38.Tilman D. Ecology. 1986;67:555–563. [Google Scholar]
- 39.Vitousek P M. Ecology. 1994;75:1861–1876. [Google Scholar]