


Abstract
Tcf/Lef family transcription factors are the downstream effectors of the Wingless/Wnt signal transduction pathway. Upon Wingless/Wnt signalling, β-catenin translocates to the nucleus, interacts with Tcf (1–3) and thus activates transcription of target genes (4,5). Tcf factors also interact with members of the Groucho (Grg/TLE) family of transcriptional co-repressors (6). We have now tested all known mammalian Groucho family members for their ability to interact specifically with individual Tcf/Lef family members. Transcriptional activation by any Tcf could be repressed by Grg-1, Grg-2/TLE-2, Grg-3 and Grg-4 in a reporter assay. Specific interactions between Tcf and Grg proteins may be achieved in vivo by tissue- or cell type-limited expression. To address this, we determined the expression of all Tcf and Grg/TLE family members in a panel of cell lines. Within any cell line, several Tcfs and TLEs are co-expressed. Thus, redundancy in Tcf/Grg interactions appears to be the rule. The ‘long’ Groucho family members containing five domains are repressors of Tcf-mediated transactivation, whereas Grg-5, which only contains the first two domains, acts as a de-repressor. As previously shown for Drosophila Groucho, we show that long Grg proteins interact with histone deacetylase-1. Although Grg-5 contains the GP homology domain that mediates HDAC binding in long Grg proteins, Grg-5 fails to bind this co-repressor, explaining how it can de-repress transcription.
INTRODUCTION
Recent studies have demonstrated that members of the Tcf/Lef family of HMG box transcription factors are important downstream effectors of the Wnt/Wingless signalling cascade in mammalian, Xenopus, Drosophila and Caenorhabditis elegans development (7). In the absence of a Wnt signal, β-catenin associates with Axin, APC and GSK-3β in the cytoplasm (8–10). In this complex, β-catenin is phosphorylated by GSK-3β, resulting in ubiquitous degradation of β-catenin by the proteasome pathway (11,12). Wnt signalling results in inhibition of GSK-3β, leading to the accumulation of β-catenin in the cytoplasm and its translocation to the nucleus. Association of β-catenin with Tcf in the nucleus leads to the formation of a bipartite transcription factor activating target gene expression, such as the genes encoding cyclin D1 (5), Tcf-1 (13) and PPARδ (14).
Experiments in Drosophila and Xenopus indicated that Tcf molecules could also function as transcriptional repressors in the absence of a Wnt/Wingless signal (15–18). Several recent studies have proposed a molecular basis for this phenomenon. Tcf can associate with a number of different transcriptional co-repressors. Binding of Tcf with CBP (19) or CtBP (20) can lead to repression of the Wnt/Wingless response. We and others have shown that the co-repressor Groucho and its vertebrate homologues can bind to Tcf. The association of Tcf and Groucho results in repression of Tcf targets both in the context of synthetic promoters and of endogenous genes (6,21,22).
The vertebrate Tcf family of transcription factors consists of four members: Tcf-1, Lef-1, Tcf-3 and Tcf-4 (Fig. 1A). Studies have indicated that Tcf-1 is preferentially expressed in cells of the T cell lineage (23). Inactivation of the Tcf-1 gene by homologous recombination results in mice with a block in T cell development (24). Lef-1 is expressed in pre-B and T cells in adult mice, and mainly in the neural crest, mesencephalon, tooth germs and whisker follicles during embryogenesis. Lef-1-deficient mice die shortly after birth and lack teeth, mammary glands, whiskers and hair, but show no defects in lymphoid cell populations (25). T cell development in mice lacking both Tcf-1 and Lef-1 is completely arrested and impaired at an earlier stage than in mice lacking only Tcf-1, revealing that Tcf-1 and Lef-1 are partially redundant in the regulation of T cell development (26). Tcf-3 is expressed in stomach epithelium, hair follicles and keratinocytes of the skin (28). Tcf-4 expression occurs much later in embryogenesis than Tcf-1, Lef-1 or Tcf-3 and is most highly expressed in the midbrain and in intestinal and mammary epithelium. Tcf-4 exhibits a highly restricted expression pattern in the epithelium of the developing gut (27,28). Continued Tcf-4 expression at this site is essential for the maintenance of the progenitor compartment of gut epithelium, as indicated in Tcf-4-deficient mice by the abnormal development of their small intestines (29). Constitutively active Tcf-4–β-catenin complexes are found in the nuclei of colon carcinoma cells with mutations in APC or β-catenin (30). Presumably this results in the uncontrolled activation of Tcf target genes, which by implication transforms colon epithelial cells and initiates polyp formation (31).
Figure 1.

Domain structure of (A) Tcf and (B) Grg constructs. Tcf proteins contain a centrally located DNA-binding HMG box and the N-terminal β-catenin interaction domain. As previously reported (6), the Grg interaction domain is located between these two domains. The Grg proteins 1–4 contain five distinct domains: Q, GP, CCN, SP and WD40 regions (51), while the Grg-5 protein consists only of the Q and GP domains.
Drosophila Groucho is the founding member of a conserved family of transcriptional co-repressors. Other members of this family are found in diverse organisms, from C.elegans to vertebrates. The murine and human genomes harbour four full-length homologues of Groucho, as well as a gene that encodes a truncated Groucho protein. The human Groucho homologues are termed TLE-1, -2, -3 and -4 (for transducin-like enhancer of split) and the truncated variant hAES (for amino-terminal enhancer of split). The mouse Groucho family consists of the Groucho-related genes, Grg-1, -2, -3 and -4 and the Grg-5 gene encoding a shorter variant (32). The Grg/TLE proteins are highly similar to Drosophila Groucho in their domain structure. They contain five protein domains: an N-terminal Q (glutamine-rich) domain, followed by a GP (glycine/proline-rich) domain, a CCN domain (containing putative casein kinase II/cdc2 phosphorylation sites and nuclear localisation signal), an SP (serine/proline-rich) domain and four WD40 repeats (protein interaction domain) (Fig. 1B). Of these, the Q and WD40 domain sequences are most highly conserved. Grg-5 and hAES encode only the two N-terminal domains of these proteins, the Q and GP domains. Groucho-related proteins have been described to tetramerise through a leucine zipper-like structure in the N-terminal Q domain (33).
Groucho proteins function as co-repressors for specific subsets of DNA-binding transcription factors, including the Hairy-related proteins, Runt domain proteins (35), Engrailed (34), Dorsal (46), Pax-5 (36), NK-3 (37), NK-4 (38), NF-κB (38) and HNF3β (39). For some of these transcription factors, the domain that interacts with Groucho is mapped to a short peptide motif. Hairy-related bHLH transcription factors are involved in diverse developmental processes such as sex determination, segmentation, neurogenesis and myogenesis in the fly. The C-terminal WRPW motif of these bHLH transcription factors binds to the SP domain of Groucho, although the WD40 domain may also be involved (34,40,41). Runt domain proteins like Drosophila Runt and human AML1 associate with Groucho proteins through a related motif, VWPRY (35). Another conserved Groucho interaction motif is found in a number of transcription factors such as Engrailed and Goosecoid, which associate with Groucho through their eh1/GEH domain (34,42). Engrailed has been shown to interact with the WD40 domain of Groucho (43). Some of these transcription factors have been described only to repress transcription dependent on Groucho for their activity, such as Hairy (40), HES (44) and Blimp-1 (45). Others, like Tcf (6), Runt (35) and Dorsal (46), are activators and convert to repressors upon association with Groucho-related proteins.
The Drosophila histone deacetylase Rpd3 has been shown to interact with Groucho (47). Recruitment of Rpd3 to the promotor of target genes will result in modulation of the local chromatin structure. Histone deacetylases remove acetyl groups from lysine residues in the N-terminal tails of core histones. By mechanisms that are not yet fully understood, this appears to result in a more compact chromatin structure that is associated with a repressed transcriptional state (48).
The current study characterises the Tcf/Groucho interaction in more detail. We describe the (co-)expression of individual Tcf and Grg/TLE family members in a panel of cell lines. In addition, we have mapped the Tcf interaction domain of Grg proteins and analysed whether there is specificity in the interaction between individual Tcf and TLE family members. Deletion analysis of Xenopus Grg-4 (XGrg-4) showed that only the first two domains, Q and GP, are sufficient to repress Tcf-mediated transcriptional activation. However, Grg-5, which contains the same, highly conserved domains, functions as a de-repressor. To determine the basis of the opposing functions of the truncated XGrg-4-QGP and XGrg-5 we examined their relative ability to interact with the human homologue of RPD3 (HDAC-1).
MATERIALS AND METHODS
Transfections and luciferase assays
IIAI.6 B cells (2.5 × 106) were transfected by electroporation with various combinations of a luciferase reporter plasmid containing three optimal Tcf sites upstream of the minimal HSV-TK promoter (1 µg of pTKTOP) or its negative control vector containing mutated Tcf sites (pTKFOP) in combination with the following plasmids: an internal transfection control (50 ng of pRNL-TK; Promega); Tcf expression vectors (2 µg); Grg expression plasmids (0.5 or 5.0 µg); β-catenin expression plasmid (5.0 µg). For the repression and de-repression experiments (see Fig. 5), 250 µg XTcf-3, 100, 50 and 25 ng Armadillo and 5 µg of Grg-5 or XGrg-4-QGP (1-197) expression plasmid were used. Additional pCDNA3 plasmid was included where necessary to make the total amount of DNA equivalent. cDNAs encoding hTCF-1, hLEF-1, XTcf-3, hTCF-4 and myc-tagged versions of Grg-1, TLE-2 and Grg-3 were expressed in pCDNA3. The expression construct for Grg-4 in the vector pKW2T was kindly provided by Dr M.Busslinger (Research Institute of Molecular Pathology, Vienna, Austria). Luciferase activities were determined 24 h after transfection using the DUAL luciferase system, according to the manufacturer’s protocol (Promega). Luciferase activity was normalised relative to RNL-luciferase activity. For every experiment, transfections were performed in duplicate and several independent experiments were performed.
Figure 5.

(A) Structure of the truncated XGrg-4-QGP molecule compared to that of Grg-5. (B) Alignment of the GP domains of long Grg homologues with mGrg-5. The amino acid residues that are unique for mGrg-5 when compared to Gmgrg-1, Mgrg-3 and XGrg-4 are depicted in bold. (C) XGrg-4-QGP functions as a repressor, but mGrg-5 functions as a de-repressor for Tcf-mediated transcriptional activation. Mouse B cell line IIAI.6 was co-transfected with the Tcf reporter construct (TOP-TK) and XTcf-3 and Armadillo (the Drosophila β-catenin homologue). This results in transactivation and increase of luciferase activity. This transactivation is de-repressed when Grg-5 is co-transfected, whereas the transactivation is repressed upon co-transfection with a truncation of XGrg-4 that only contains the Q and GP domains. Transfections were performed in duplicate, and luciferase values were corrected for the efficiency of transfection using the internal Renilla transfection control pRNL-TK by determining the luciferase/Renilla ratio. This ratio is given on the y-axis.
Yeast two-hybrid assay
Two-hybrid experiments were carried out as described previously (27). Briefly, the Tcf (4-359) and Grg-5 (1-197) bait constructs were created by fusing the regions encoding the indicated amino acids to the GAL4 binding domain of pMD4. pVA3 encodes a murine P53-GAL4 binding domain hybrid in pGBT9 (Clontech). Multiple preys, mGrg-5 (1-197, 4-106, 82-197, 4-62, 46-106, 82-106) were inserted in-frame with the GAL4 activation domain in pGADGH (Clontech) or pGADRX (Stratagene). pTD1 encodes SV40 large T antigen in pGAD3F (Clontech). Baits and preys were transformed into the Saccharomyces cerevisiae strain HF7C (Clontech) and true interactions were scored by growth on selective nutrient agar plates. Equivalent results were obtained monitoring β-galactosidase activity (data not shown).
Cell lines
For the transfection experiments, the mouse B cell line IIAI.6 was used. For the reverse transcription-coupled PCR experiments, the following human cell lines were used as a source of RNA: Jurkat and CEM (T cell lines), Reh and Raji (B cell lines), 293T (embryonic kidney cell line), HT29, LS174, SW480, SW620, DLD-1, RKO (colon carcinoma cell lines) and SKBR3 and T47D (mammary carcinoma cell lines). Cells were cultured in RPMI-1640 supplemented with 10% foetal calf serum and antibiotics.
Reverse transcriptase–polymerase chain reaction (RT–PCR) analysis
Total RNA for detection of hTcf1, hLef-1, hTcf3, hTcf4, TLE-1, TLE-2, TLE-3 and TLE-4 was isolated from the different cell lines using RNAzolB solution (Tel Test Inc.) according to the manufacturer’s instructions. Random-primed cDNA from total RNA was prepared using standard techniques. The following primers were used for the different PCR reactions: for Tcf-1, Lef-1 and Tcf-4, specific primers were used to amplify part of the mRNA corresponding to the NH2-terminus upstream of the HMG box. For hTcf-3D, primers were developed to amplify the HMG box since this was the only sequence available at the time. Tcf1 primers: 5′-ACCAGCGGCATGTACAAAGAG-3′ (sense), 5′-TTCAGGTTGCGGTCGAAGGGC-3′ (antisense). Lef-1 primers: 5′-TTCTCCACCCATCCCGAGAAC-3′ (sense), 5′-CTGAGGCTTCACGTGCATTTAG-3′ (antisense). Tcf-4 primers: 5′-CCATCACCGGCACACATTGTC-3′ (sense), 5′-ACTATGGTGTGAGCCGACATC-3′(antisense). Tcf-3 primers: 5′-GAAATCACCAGTCACCGTGAAA-3′ (sense), 5′-ACCAGGTTGGGTAGAGCTGCG-3′ (antisense).
For TLE-1, TLE-2 and TLE-3, specific primers were used to amplify a fragment of GP domain, CCN domain and part of the SP domain. For hTLE-4, a part of the WD domain was amplified since this was the only sequence available for this gene. For hAESD, the entire gene, both the Q and GP domains were amplified. hTLE-1 primers: 5′-GGCAGTGCCGGCCTTCTTGCG-3′ (sense), 5′-AGGCTTGCCGAGACCTGGACG-3′ (antisense). hTLE-2 primers: 5′-AGTGCTACGGGGCTGCTTGCT-3′ (sense), 5′-CGTTGAGAGTGCTGTGGGAGC-3′ (antisense). hTLE-3 primers: 5′-AGCAGCTCCGGGCTGCTGGCA-3′ (sense), 5′-AGCCGAGGCCATTATACCTAT-3′ (antisense). hTLE-4 primers: 5′-TACACGGGTGGGAAGGGCGCG-3′ (sense), 5′-GGCTTGGTGACATGCAAAACT-3′ (antisense). hAES primers 5′-CTCAAATTCACCACCTCGGAG-3′ (sense), 5′-ATCCTCCTGGTGGGTGTCACC-3′ (antisense).
The primers for the GAPDH control PCR were: 5′-AAGGTGAAGGTCGGAGTCAAC-3′ (sense), 5′-TTACTCCTTGGAGGCCATGTG-3′ (antisense).
Immune precipitations
293T cells were co-transfected with pCDNA3-Flag-HDAC-1 (kindly provided by T.Kouzarides, Wellcome/CRC Institute, Cambridge, UK) and pCDNA3-MYC-XGrg4-QGP (amino acids 1–197) or pCDNA3-MYC-Grg5. Cells were harvested and whole cell lysate was prepared in Triton X-100 lysis buffer (20 mM Tris pH 8.0, 140 mM NaCl, 1% Triton X-100 and 10% glycerol) containing protease inhibitors (Protease inhibitor cocktail tablets; Roche Diagnostics). The extract was incubated with 1 µg anti-Myc monoclonal antibody and 5 µl protein A–G beads (Santa Cruz) or with 5 µl FLAG-coupled beads (Sigma) overnight at 4°C in Triton X-100 lysis buffer. The beads were washed twice in Triton X-100 buffer and twice in wash buffer (20 mM Tris pH 8.0, 150 mM NaCl and 0.5% NP-40) Supernatants were resolved by SDS–PAGE and transferred to nitrocellulose membrane (Immobilon-P; Millipore). The membrane was incubated with anti-FLAG (Sigma) or anti-Myc antibodies and immune reactive proteins were visualised by enhanced chemiluminescence (Amersham Pharmacia Biotech).
RESULTS AND DISCUSSION
A 100 amino acid region in Grg-5 is essential for interaction with Tcf
The interaction between Tcf and Grg was previously mapped to the QGP domains of Groucho, since Tcf interacts with Grg-5, consisting only of these two domains (49). Grg proteins can also tetramerise through their Q domain (33). We used the yeast two-hybrid assay to more precisely map the domain of Grg that interacts with Tcf versus the domain that is responsible for multimerization with other Grg molecules (Fig. 2). Constructs expressing different parts of Grg-5 were fused to the Gal-4 activation domain and their interaction with Tcf-1 (4-359) and Grg-5 (1-197) fused to the Gal-4 DNA binding domain was determined. The results shown in Figure 2 imply that the Tcf interaction domain resides in the first 106 amino acids of Grg-5. All other deletion constructs of Grg-5 failed to interact with Tcf. Our data indicate this N-terminal region also contains the Grg interaction domain, in accordance with the domain previously described to mediate tetramerisation (33). Further truncation of this region probably prevents proper folding of the tetramerisation domain and thereby results in loss of the interaction with both Grg and Tcf.
Figure 2.

Mapping of the Grg–Tcf interaction domain and Grg multimerisation domain. The different bait Grg-5 constructs used in the yeast two-hybrid assay are shown on the left. The ability of these constructs to interact with prey molecules Tcf-1 (4-359) or Grg-5 (1-197) and allow growth of yeast on selective plates is indicated. The minimal domain of Grg-5 required for interaction with Tcf resides in the first 106 amino acids. Further truncation of this domain results in loss of interaction.
Repression of TCF-mediated transcription by Groucho-related genes
To further investigate the issue of specificity in the interaction between different Tcf family members and Grg proteins, we used the previously established β-catenin–Tcf reporter gene assay (1). In this assay co-transfection of Tcf and β-catenin results in transactivation of a luciferase reporter gene, which we previously demonstrated could be repressed by introduction of Groucho (6). Here, the four different Tcf family members were co-transfected with β-catenin and increasing amounts of Grg-1, TLE-2, Grg3 or Grg-4. An expression construct of human TLE-2 was included instead of mGrg-2, since the latter had not been described at the time. Three human Tcf family members were used: hTcf-1, hLef-1 and hTcf-4 (Fig. 3A, B and D, respectively). For Tcf-3 we used the Xenopus homologue (Fig. 3C) because human Tcf-3 proved difficult to express (data not shown). Identical transfections were carried out using a reporter with mutated Tcf sites. A Renilla luciferase vector was also transfected to correct for transfection efficiency. [Note that the B cell line used for these assays expresses moderate levels of both Grg-1 and Grg-4 (6)]. These experiments show that all different Grg proteins can repress transcriptional activation mediated by all Tcf family members. Grg-1 and Grg-4 appear to be more potent repressors than Grg-3 and TLE-2. In this assay, the Tcf proteins show differences in transactivation capacity: hTcf-1 and Lef-1 transactivate more strongly than XTcf-3 and hTcf-4, but for all Tcf family members repression is observed upon co-transfection with the different Groucho homologues.
Figure 3.




All ‘long’ Grg proteins can repress transactivation mediated by all Tcf family members. Mouse B cell line IIAI.6 was co-transfected with the Tcf reporter construct (containing either wild-type Tcf binding sites, TOP-TK, or mutated Tcf binding sites, FOP-TK, as a negative control) and a specific Tcf family member and β-catenin. This results in transactivation and increase of luciferase activity. Upon addition of increasing amounts of any of the Grg family members, the luciferase activity decreases proportionally. (A) Repression of Tcf-1-mediated transcriptional activation by Grg-1, TLE-2, Grg-3 and Grg-4. (B) Repression of Lef-1-mediated transcriptional activation by Grg-1, TLE-2, Grg-3 and Grg-4. (C) Repression of Tcf-3-mediated transcriptional activation by Grg-1, TLE-2, Grg-3 and Grg-4. (D) Repression of Tcf-4-mediated transcriptional activation by Grg-1, TLE-2, Grg-3 and Grg-4. Transfections were performed in duplicate, and results from one representative independent experiment are depicted in each case. Luciferase values were corrected for the efficiency of transfection using the internal Renilla transfection control pRNL-TK by determining the luciferase/Renilla ratio. This ratio is given on the y-axis, and was arbitrarily set at 1 for the sample in which the TOP-TK reporter construct alone was transfected.
Expression of the different Tcf and TLE family members in a panel of cell lines
We examined whether there was any specificity in the interaction between Tcf and TLE proteins based on their expression pattern in a panel of cell lines. Two B cell lines (Reh and Raji) and two T cell lines (Cem and Jurkat) were tested, as well as a number of colon carcinoma cell lines (HCT116, LS174, HT29, DLD-1, SW480, SW620 and RKO) and two mammary carcinoma cell lines (T47D and SKBR3). In colon carcinoma cell lines the Wnt signalling cascade is constitutively activated. In all but one (RKO) of these colon carcinoma cell lines, mutations in either APC or β-catenin have been identified. Absence of TLE expression in the RKO cell line could potentially account for activation of the pathway in this cell line.
RNA was isolated from the above cell lines and first strand cDNA was prepared by RT. This cDNA was used as a template in PCR to assay for the expression of the different Tcf and TLE family members (Fig. 4). As a negative control, reverse transcriptase was omitted from the RT reaction. PCR for the different Tcf and Grg family members was carried out. A GAPDH PCR served as an internal control for the quality of the prepared cDNA. The PCR products were designed to be of different sizes to distinguish between the different family members. A product of each PCR was sequenced to confirm the identity of the amplified product and to exclude any cross-reactivity between family members (data not shown).
Figure 4.

Expression of (A) hTCF-1, hLEF-1, hTCF-3, hTCF-4; and (B) TLE-1, TLE-2, TLE-3, TLE-4 and hAES was determined by RT–PCR using cDNA generated from a panel of cell lines. Reh (lane 1), Raji (lane 2), CEM (lane 3), Jurkat (lane 4), 293T (lane 5), HCT116 (lane 6), LS174T (lane 7), HT29 (lane 8), DLD-1 (lane 9), SW480 (lane 10), SW620 (lane 11), RKO (lane 12), T47D (lane 13), SKBR3 (lane 14) and H2O control for the PCR (lane 15). The lane marked λ depicts the DNA marker (bacteriophage λ DNA digested with EcoRI and HindIII). Expression is tabulated for Tcfs and TLEs for the given cell lines. +, visible RT–PCR product; –, no visible RT–PCR product.
Expression of hTCF-1 was detected in all cell lines tested except in the B cell lines. In the cell lines where hTCF-1 is expressed, a very faint band was also observed running more slowly in the gel. This corresponds to a splice variant of Tcf-1 containing an extra exon, as described earlier (41). hLEF-1 expression was found in Reh, CEM, Jurkat, 293T, LS174T, DLD-1, SW480, SW620, RKO and T47D. It was known previously to be expressed in pre-B and T cells and several other cell types (25), but apparently it is not expressed in all colon or mammary carcinoma cell lines. The two different products present in most hLEF-1-expressing cell lines correspond to the presence of two different splice variants (50), as was confirmed by sequencing (data not shown). hTCF-3 expression was restricted to Jurkat, 293T, some colon carcinoma cell lines (HCT116, DLD-1, SW480 and SW620) and both mammary carcinoma cell lines T47D and SKBR3. For this family member, a more restricted expression pattern was expected since hTCF-3 is mainly expressed during early embryogenesis (28). hTCF-4 is expressed ubiquitously in all cell lines tested.
The expression of the Groucho family members also showed a relatively broad pattern. Within the panel of cell lines, only the expression pattern of hTLE-2 was restricted, being expressed only in Reh, Jurkat, HCT-116, RKO, T47D and SKBR-3. For hAES, hTLE-1, hTLE-3 and hTLE-4, a broad expression pattern was observed. hAES is expressed in all cell lines tested but at a low level in Raji, 293T and T47D. hTLE-1 is only absent in Raji and SW620 cell lines. hTLE-4 is present in all cell lines of this panel with the exception of HCT116 and LS174T. hTLE-3 is present in all cell lines in this panel. Corresponding with this broad TLE expression in cell lines, it has been shown that the TLE proteins are also broadly expressed in adult tissues (51).
These data show that a number of diverse cell lines all express multiple Tcf and TLE family members. This indicates that the pattern of expression does not contribute to a possible specificity of interaction between Tcf and TLE molecules. It does imply, however, that two or more Groucho repressor proteins are normally available for a tight control over the Tcf transcriptional activation pathway, since deregulation of this pathway can result in tumourigenesis, as has been shown for colon carcinoma (31) and melanoma (52).
Domains of Grg required for repression of Tcf-mediated transactivation and dependency on HDAC interaction
Deletion constructs of XGrg-4 were tested for their ability to repress Tcf-mediated transcriptional activation (data not shown). The minimal construct of Xenopus Grg-4 (XGrg-4) required for repression consists of only the first two N-terminal domains. This truncation of XGrg-4, consisting of amino acids 1–197, encodes the Q domain, involved in tetramerisation and interaction with Tcf, and the GP domain, essential for interaction with the histone deacetylase-1, HDAC (53). This construct is highly homologous to Grg-5, which is comprised of only the Q and GP domains, but functions as a de-repressor (6) (Fig. 5A). In Figure 5B, an alignment of the GP domains of mGrg-1, mGrg-3, XGrg-4 and mGrg-5 is depicted, showing the high homology between these proteins. The amino acids unique for Grg-5 are indicated in bold.
Cells were transfected with the Tcf reporter construct (TK-TOP), XTcf-3 and different amounts of Armadillo (the Drosophila homologue of β-catenin). XGrg-4-QGP consistently repressed transcription, while Grg-5 enhanced the Armadillo-mediated transactivation (Fig. 5C).
The GP domains of proteins of the Groucho family are highly conserved, and this domain is involved in interaction with HDAC (53). The subtle differences between the GP domain of the long Groucho homologues and the GP domain of the shorter variants like Grg-5 are possibly responsible for these antagonistic functions (Fig. 5B). To address this, the interaction of HDAC with XGrg-4-QGP and mGrg-5 was studied in immunoprecipitation experiments (Fig. 6). Flag-HDAC-1 construct was co-transfected with the Myc-tagged Grg constructs of mGrg-5 and the truncated XGrg-4-QGP. Cell lysates were immunoprecipitated with anti-Myc antibody and protein A–G beads. The presence of HDAC was assayed on an immunoblot using the Flag antibody. Figure 6A shows that precipitation of Myc-tagged Grg-5 and XGrg-4-QGP results in co-precipitation of HDAC with XGrg-4-QGP (lane 3) but not with Grg-5 (lane 4), although HDAC expression in the lysate of the cells co-transfected with mGrg-5 and HDAC-1 is apparent (lane 5). Lanes 6 and 7 show that the two Myc-tagged Grg constructs were immunoprecipitated with similar affinity. Similar results are obtained when Flag-tagged HDAC is immunoprecipitated and the presence of mGrg-5 and XGrg-4 QGP is assayed (Fig. 6B). Precipitation of Flag-tagged HDAC co-precipitates XGrg-4-QGP but not Grg-5 (Fig. 6B, lanes 3 and 4, respectively). Association of HDAC with full-length Grg-3 was also observed (data not shown).
Figure 6.
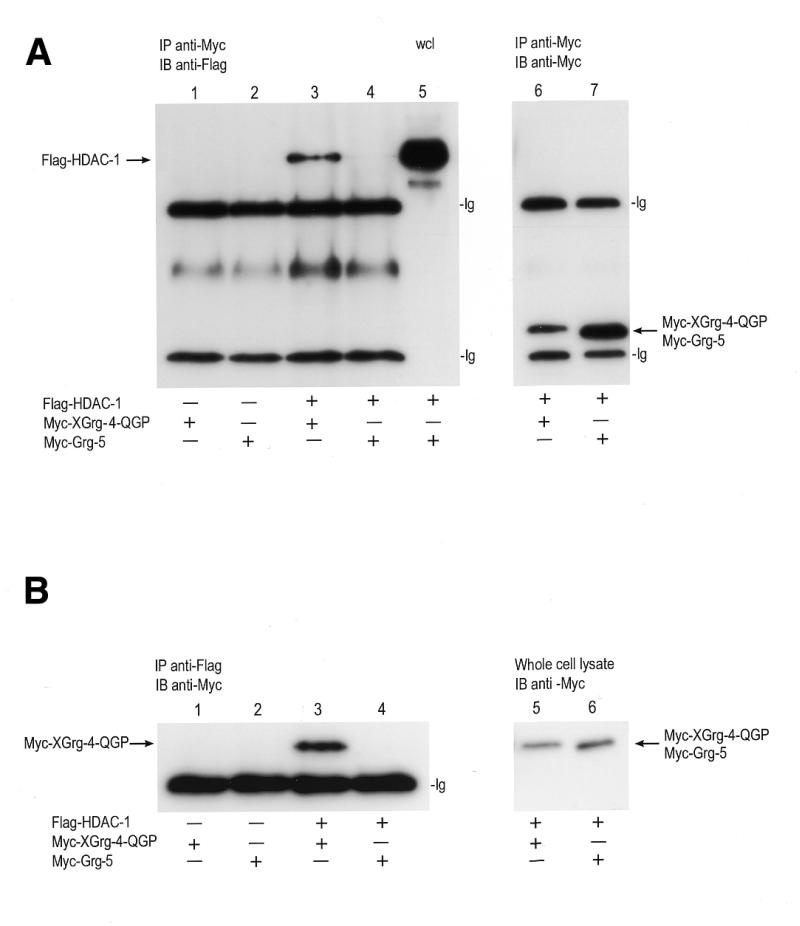
HDAC-1 associates with XGrg-4-QGP, but not with mGrg-5. 293T cells were transfected with either Myc-tagged XGrg-4 QGP or Myc-tagged Grg-5 alone (lanes 1 and 2), or with a Flag-tagged HDAC-1 (lanes 3 and 4). Cell lysates were prepared 24 h after transfection. (A) Extracts from transfected cells were immunoprecipitated (IP) with an anti-Myc antibody against Myc-XGrg-4-QGP and Myc-Grg-5, and the presence of Flag-HDAC-1 in the immunoprecipitate was assayed on immunoblot (IB) with anti-Flag antibody. In lanes 1 and 2, only Myc-tagged XGrg-4-QGP or Myc-Grg-5 were transfected. In lanes 3 and 4, Myc-tagged XGrg-4-QGP or Myc-Grg-5 were co-transfected with Flag-tagged HDAC-1. Lane 5 shows the presence of HDAC-1 in the co-transfection of Myc-Grg-5 and Flag-HDAC-1. Lanes 6 and 7 show an immunoblot with anti-Myc antibody, indicating the expression of the Myc-Grg-4-QGP and Myc-Grg-5 when co-transfected with Flag-HDAC-1. (B) Extracts from transfected cells were immunoprecipitated (IP) with an anti-Flag antibody against Flag-HDAC-1. The presence of Myc-XGrg-4-QGP and Myc-Grg-5 was assayed on immunoblot (IB) with anti-Myc antibody. In lanes 1 and 2 only Myc-tagged XGrg-4-QGP or Myc-Grg-5 were transfected. In lanes 3 and 4, Myc-tagged XGrg-4-QGP or Myc-Grg-5 were co-transfected with Flag-tagged HDAC-1. Lanes 5 and 6 show an immunoblot with anti-Myc antibody, indicating the expression of Myc-Grg-4-QGP and Myc-Grg-5 when co-transfected with Flag-HDAC-1. Bands corresponding to HDAC, Grg-5 and XGrg-4 QGP are indicated with arrows. Bands labelled ‘Ig’ indicate the heavy and light chains of the antibodies used for the immune precipitation.
These data show that the long, repressive vertebrate Groucho homologues readily associate with HDAC-1, as was described previously in the fly (53). The de-repressor Grg-5, however, does not bind HDAC, explaining why Grg-5 does not function as a repressor.
Grg-5 may function to fine-tune the repression mediated by Groucho family members. In one possible model, Grg-5 would associate with long repressive Grg proteins and thereby decrease the amount of HDAC that is tethered to the template, reducing the level of repression. Alternatively, Grg-5 could bind directly to a subclass of transcription factors interacting with the first two domains of Groucho and prevent long Grg family members associating with such factors, abrogating repression entirely (Fig. 7). The precise activation status of a given transcription factor interacting with Groucho proteins in vivo would be partially determined by the mutual antagonism between long and short forms, the balance of which could be influenced by protein expression levels, subcellular distribution or post-translational modifications of Groucho proteins.
Figure 7.

Model for Tcf, long Grg and Grg-5 functions. Expression of Tcf target genes is activated when Tcf is associated with β-catenin and repressed when Tcf binds Groucho. Here we show that the function of Grg-5 provides another level of regulation. When in a complex, both the long repressor Grg and Grg-5 are bound to Tcf, a less competent repressor is formed, since less HDAC activity is tethered to the promotor. When Grg-5 replaces the longer repressor entirely from the complex with Tcf, repression is absent because HDAC activity is lacking.
We find no specificity in the interactions between the different members of the Tcf and Groucho families. Every cell line tested expresses several TLE and Tcf family members. Furthermore, reporter assays reveal that any long Grg can repress any given Tcf, suggesting that all partners are capable of interacting in vivo. The abundance of Groucho molecules and the redundancy with which they interact with Tcf proteins probably highlights the requirement to silence target gene expression adequately, in order to prevent unrestrained cell growth and/or aberrant differentiation, hallmarks of cancer cells. It still remains a possibility that some specificity is achieved by other means not identified in this study. For example, TLE phosphorylation may influence their repressive activity, since Cdc2 and casein kinase II consensus sites are found in the Grg/TLE sequences and it has previously been shown that the TLE phosphorylation pattern changes during differentiation (54).
The mechanism of Groucho-mediated repression remained unclear until recently, when it was shown that Groucho functionally interacts with the histone deacetylase Rpd3 (53). Recruitment of Rpd3 to a target promoter would result in modification of local chromatin structure and, consequently, the formation of a more compact chromatin structure, associated with a repressed transcriptional state. This is likely the main mechanism through which Groucho-related proteins exert their function, although other data indicate that an additional mechanism of repression may exist. When different Grg constructs are tethered to a Gal4 DNA binding domain, domains capable of mediating repression are also found outside the GP domain (40) Moreover, when Groucho-mediated repression is assayed in the presence of the HDAC inhibitor trichostatin A, some residual repression activity remains (53). When the histone deacetylase RPD3 in Drosophila is mutated, only mild defects are observed (55). All these studies suggest that HDAC is an important, but not the sole, mediator of transcriptional repression. Further study of the mechanism of Groucho-mediated repression will address these subjects in the future.
References
- 1.van de Wetering M., Cavallo,R., Dooijes,D., van Beest,M., van Es,J., Loureiro,J., Ypma,A., Hursh,D., Jones,T., Bejsovec,A., Peifer,M., Mortin,M. and Clevers,H. (1997) Armadillo coactivates transcription driven by the product of the Drosophila segment polarity gene dTCF. Cell, 88, 789–799. [DOI] [PubMed] [Google Scholar]
- 2.Molenaar M., van de Wetering,M., Oosterwegel,M., Peterson Maduro,J., Godsave,S., Korinek,V., Roose,J., Destree,O. and Clevers,H. (1996) XTcf-3 transcription factor mediates β-catenin-induced axis formation in Xenopus embryos. Cell, 86, 391–399. [DOI] [PubMed] [Google Scholar]
- 3.Behrens J., von Kries,J.P., Kuhl,M., Bruhn,L., Wedlich,D., Grosschedl,R. and Birchmeier,W. (1996) Functional interaction of β-catenin with the transcription factor LEF-1. Nature, 382, 638–642. [DOI] [PubMed] [Google Scholar]
- 4.He T.C., Sparks,A.B., Rago,C., Hermeking,H., Zawel,L., da Costa,L.T., Morin,P.J., Vogelstein,B. and Kinzler,K.W. (1998) Identification of c-MYC as a target of the APC pathway. Science, 281, 1509–1512. [DOI] [PubMed] [Google Scholar]
- 5.Tetsu O. and McCormick,F. (1999) β-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature, 398, 422–426. [DOI] [PubMed] [Google Scholar]
- 6.Roose J., Molenaar,M., Peterson,J., Hurenkamp,J., Brantjes,H., Moerer,P., van de Wetering,M., Destree,O. and Clevers,H. (1998) The Xenopus Wnt effector XTcf-3 interacts with Groucho-related transcriptional repressors. Nature, 395, 608–612. [DOI] [PubMed] [Google Scholar]
- 7.Cadigan K.M. and Nusse,R. (1997) Wnt signaling: a common theme in animal development. Genes Dev., 11, 3286–3305. [DOI] [PubMed] [Google Scholar]
- 8.Behrens J., Jerchow,B.A., Wurtele,M., Grimm,J., Asbrand,C., Wirtz,R., Kuhl,M., Wedlich,D. and Birchmeier,W. (1998) Functional interaction of an axin homolog, conductin, with β-catenin, APC and GSK3β. Science, 280, 596–599. [DOI] [PubMed] [Google Scholar]
- 9.Nakamura T., Hamada,F., Ishidate,T., Anai,K., Kawahara,K., Toyoshima,K. and Akiyama,T. (1998) Axin, an inhibitor of the Wnt signalling pathway, interacts with β-catenin, GSK-3β and APC and reduces the β-catenin level. Genes Cells, 3, 395–403. [DOI] [PubMed] [Google Scholar]
- 10.Rubinfeld B., Albert,I., Porfiri,E., Fiol,C., Munemitsu,S. and Polakis,P. (1996) Binding of GSK3β to the APC-β-catenin complex and regulation of complex assembly. Science, 272, 1023–1026. [DOI] [PubMed] [Google Scholar]
- 11.Aberle H., Bauer,A., Stappert,J., Kispert,A. and Kemler,R. (1997) β-catenin is a target for the ubiquitin-proteasome pathway. EMBO J., 16, 3797–3804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kitagawa M., Hatakeyama,S., Shirane,M., Matsumoto,M., Ishida,N., Hattori,K., Nakamichi,I., Kikuchi,A. and Nakayama,K. (1999) An F-box protein, FWD1, mediates ubiquitin-dependent proteolysis of β-catenin. EMBO J., 18, 2401–2410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Roose J., Huls,G., van Beest,M., Moerer,P., van der Horn,K., Goldschmeding,R., Logtenberg,T. and Clevers,H. (1999) Synergy between tumor suppressor APC and the β-catenin-Tcf4 target Tcf1. Science, 285, 1923–1926. [DOI] [PubMed] [Google Scholar]
- 14.He T.C., Chan,T.A., Vogelstein,B. and Kinzler,K.W. (1999) PPARδ is an APC-regulated target of nonsteroidal anti-inflammatory drugs. Cell, 99, 335–345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bienz M. (1998) TCF: transcriptional activator or repressor? Curr. Opin. Cell Biol., 10, 366–372. [DOI] [PubMed] [Google Scholar]
- 16.Brannon M., Gomperts,M., Sumoy,L., Moon,R.T. and Kimelman,D. (1997) A β-catenin/XTcf-3 complex binds to the siamois promoter to regulate dorsal axis specification in Xenopus. Genes Dev., 11, 2359–2370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fan M.J., Gruning,W., Walz,G. and Sokol,S.Y. (1998) Wnt signaling and transcriptional control of Siamois in Xenopus embryos. Proc. Natl Acad. Sci. USA, 95, 5626–5631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Riese J., Yu,X., Munnerlyn,A., Eresh,S., Hsu,S.C., Grosschedl,R. and Bienz,M. (1997) LEF-1, a nuclear factor coordinating signaling inputs from wingless and decapentaplegic. Cell, 88, 777–787. [DOI] [PubMed] [Google Scholar]
- 19.Waltzer L. and Bienz,M. (1998) Drosophila CBP represses the transcription factor tcf to antagonize Wingless signaling. Nature, 395, 521–525. [DOI] [PubMed] [Google Scholar]
- 20.Brannon M., Brown,J.D., Bates,R., Kimelman,D. and Moon,R.T. (1999) XCtBP is a XTcf-3 co-repressor with roles throughout Xenopus development. Development, 126, 3159–3170. [DOI] [PubMed] [Google Scholar]
- 21.Cavallo R., Cox,R., Moline,M., Roose,J., Polevoy,G., Clevers,H., Peifer,M. and Bejsovec,A. (1998) Drosophila Tcf and Groucho interact to repress Wingless signalling activiy. Nature, 395, 604–608. [DOI] [PubMed] [Google Scholar]
- 22.Levanon D., Goldstein,R.E., Bernstein,Y., Tang,H., Goldenberg,D., Stifani,S., Paroush,Z. and Groner,Y. (1998) Transcriptional repression by AML1 and LEF-1 is mediated by the TLE/Groucho corepressors. Proc. Natl Acad. Sci. USA, 95, 11590–11595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Oosterwegel M., van de Wetering,M., Timmerman,J., Kruisbeek,A., Destree,O., Meijlink,F. and Clevers,H. (1993) Differential expression of the HMG box factors TCF-1 and LEF-1 during murine embryogenesis. Development, 118, 439–448. [DOI] [PubMed] [Google Scholar]
- 24.Verbeek S., Izon,D., Hofhuis,F., Robanus Maandag,E., te Riele,H., van de Wetering,M., Oosterwegel,M., Wilson,A., MacDonald,H.R. and Clevers,H. (1995) An HMG-box-containing T-cell factor required for thymocyte differentiation. Nature, 374, 70–74. [DOI] [PubMed] [Google Scholar]
- 25.van Genderen C., Okamura,R.M., Farinas,I., Quo,R.G., Parslow,T.G., Bruhn,L. and Grosschedl,R. (1994) Development of several organs that require inductive epithelial–mesenchymal interactions is impaired in LEF-1-deficient mice. Genes Dev., 8, 2691–2703. [DOI] [PubMed] [Google Scholar]
- 26.Okamura R.M., Sigvardsson,M., Galceran,J., Verbeek,S., Clevers,H. and Grosschedl,R. (1998) Redundant regulation of T cell differentiation and TCRα gene expression by the transcription factors LEF-1 and TCF-1. Immunity, 8, 11–20. [DOI] [PubMed] [Google Scholar]
- 27.Barker N., Huls,G., Korinek,V. and Clevers,H. (1999) Restricted high level expression of Tcf-4 protein in intestinal and mammary gland epithelium. Am. J. Pathol., 154, 29–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Korinek V., Barker,N., Willert,K., Molenaar,M., Roose,J., Wagenaar,G., Markman,M., Lamers,W., Destree,O. and Clevers,H. (1998) Two members of the Tcf family implicated in Wnt/β-catenin signaling during embryogenesis in the mouse. Mol. Cell. Biol., 18, 1248–1256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Korinek V., Barker,N., Moerer,P., van Donselaar,E., Huls,G., Peters,P.J. and Clevers,H. (1998) Depletion of epithelial stem-cell compartments in the small intestine of mice lacking Tcf-4. Nature Genet., 19, 379–383. [DOI] [PubMed] [Google Scholar]
- 30.Korinek V., Barker,N., Morin,P.J., van Wichen,D., de Weger,R., Kinzler,K.W., Vogelstein,B. and Clevers,H. (1997) Constitutive transcriptional activation by a β-catenin-Tcf complex in APC–/– colon carcinoma. Science, 275, 1784–1787. [DOI] [PubMed] [Google Scholar]
- 31.Morin P.J., Sparks,A.B., Korinek,V., Barker,N., Clevers,H., Vogelstein,B. and Kinzler,K.W. (1997) Activation of β-catenin-Tcf signaling in colon cancer by mutations in β-catenin or APC. Science, 275, 1787–1790. [DOI] [PubMed] [Google Scholar]
- 32.Fisher A.L. and Caudy,M. (1998) Groucho proteins: transcriptional corepressors for specific subsets of DNA-binding transcription factors in vertebrates and invertebrates. Genes Dev., 12, 1931–1940. [DOI] [PubMed] [Google Scholar]
- 33.Chen G., Nguyen,P.H. and Courey,A.J. (1998) A role for Groucho tetramerization in transcriptional repression. Mol. Cell Biol., 18, 7259–7268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Jimenez G., Paroush,Z. and Ish Horowicz,D. (1997) Groucho acts as a corepressor for a subset of negative regulators, including Hairy and Engrailed. Genes Dev., 11, 3072–3082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Aronson B.D., Fisher,A.L., Blechman,K., Caudy,M. and Gergen,J.P. (1997) Groucho-dependent and -independent repression activities of Runt domain proteins. Mol. Cell Biol., 17, 5581–5587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Eberhard D., Jimenez,G., Heavey,B. and Busslinger,M. (2000) Transcriptional repression by Pax5 (BSAP) through interaction with corepressors of the Groucho family. EMBO J., 19, 2292–2303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Choi C.Y., Kim,Y.H., Kwon,H.J. and Kim,Y. (1999) The homeodomain protein NK-3 recruits Groucho and a histone deacetylase complex to repress transcription. J. Biol. Chem., 274, 33194–33197. [DOI] [PubMed] [Google Scholar]
- 38.Choi C.Y., Lee,Y.M., Kim,Y.H., Park,T., Jeon,B.H., Schulz,R.A. and Kim,Y. (1999) The homeodomain transcription factor NK-4 acts as either a transcriptional activator or repressor and interacts with the p300 coactivator and the Groucho corepressor. J. Biol. Chem., 274, 31543–31552. [DOI] [PubMed] [Google Scholar]
- 39.Wang J.C., Waltner-Law,M., Yamada,K., Osawa,H., Stifani,S. and Granner,D.K. (2000) Transducin-like enhancer of split proteins, the human homologs of Drosophila groucho, interact with hepatic nuclear factor 3β. J. Biol. Chem., 275, 18418–18423. [DOI] [PubMed] [Google Scholar]
- 40.Fisher A.L., Ohsako,S. and Caudy,M. (1996) The WRPW motif of the hairy-related basic helix–loop–helix repressor proteins acts as a 4-amino-acid transcription repression and protein–protein interaction domain. Mol. Cell Biol., 16, 2670–2677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Paroush Z., Finley,R.L.,Jr, Kidd,T., Wainwright,S.M., Ingham,P.W., Brent,R. and Ish Horowicz,D. (1994) Groucho is required for Drosophila neurogenesis, segmentation and sex determination and interacts directly with hairy-related bHLH proteins. Cell, 79, 805–815. [DOI] [PubMed] [Google Scholar]
- 42.Jimenez G., Verrijzer,C.P. and Ish Horowicz,D. (1999) A conserved motif in goosecoid mediates groucho-dependent repression in Drosophila embryos. Mol. Cell Biol., 19, 2080–2087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tolkunova E.N., Fujioka,M., Kobayashi,M., Deka,D. and Jaynes,J.B. (1998) Two distinct types of repression domain in engrailed: one interacts with the groucho corepressor and is preferentially active on integrated target genes. Mol. Cell Biol., 18, 2804–2814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Grbavec D. and Stifani,S. (1996) Molecular interaction between TLE1 and the C-terminal domain of HES-1 containing the WRPW motif. Biochem. Biophys. Res. Commun., 223, 701–705. [DOI] [PubMed] [Google Scholar]
- 45.Ren B., Chee,K.J., Kim,T.H. and Maniatis,T. (1999) PRDI-BF1/Blimp-1 repression is mediated by corepressors of the Groucho family of proteins. Genes Dev., 13, 125–137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Dubnicoff T., Valentine,S.A., Chen,G., Shi,T., Lengyel,J.A., Paroush,Z. and Courey,A.J. (1997) Conversion of dorsal from an activator to a repressor by the global corepressor Groucho. Genes Dev., 11, 2952–2957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Stevanovic M., Lovell Badge,R., Collignon,J. and Goodfellow,P. (1993) SOX3 is an X-linked gene related to SRY. Hum. Mol. Genet., 2, 2013–2018. [DOI] [PubMed] [Google Scholar]
- 48.Chen G. and Courey,A.J. (2000) Groucho/TLE family proteins and transcriptional repression. Gene, 249, 1–16. [DOI] [PubMed] [Google Scholar]
- 49.Mallo M., del Amo,F and Gridley,T. (1993) Cloning and developmental expression of Grg, a mouse gene related to the groucho transcript of the Drosophila enhancer of split complex. Mech. Dev., 42, 67–76. [DOI] [PubMed] [Google Scholar]
- 50.Carlsson P., Waterman,M.L. and Jones,K.A. (1993) The hLEF/TCF-1 α HMG protein contains a context-dependent transcriptional activation domain that induces the TCR α enhancer in T cells. Genes Dev., 7, 2418–2430. [DOI] [PubMed] [Google Scholar]
- 51.Stifani S., Blaumueller,C.M., Redhead,N.J., Hill,R.E. and Artavanis-Tsakonas,S. (1992) Human homologs of a Drosophila enhancer of split gene product define a novel family of nuclear proteins [published erratum appears in Nature Genet. (1992), 4, 343]. Nature Genet., 2, 119–127. [DOI] [PubMed] [Google Scholar]
- 52.Rubinfeld B., Robbins,P., El Gamil,M., Albert,I., Porfiri,E. and Polakis,P. (1997) Stabilization of β-catenin by genetic defects in melanoma cell lines. Science, 275, 1790–1792. [DOI] [PubMed] [Google Scholar]
- 53.Chen G., Fernandez,J., Mische,S. and Courey,A.J. (1999) A functional interaction between the histone deacetylase Rpd3 and the corepressor groucho in Drosophila development. Genes Dev., 13, 2218–2230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Liu Y., Dehni,G., Purcell,K.J., Sokolow,J., Carcangiu,M.L., Artavanis Tsakonas,S. and Stifani,S. (1996) Epithelial expression and chromosomal location of human TLE genes: implications for notch signaling and neoplasia. Genomics, 31, 58–64. [DOI] [PubMed] [Google Scholar]
- 55.Mannervik M. and Levine,M. (1999) The Rpd3 histone deacetylase is required for segmentation of the Drosophila embryo. Proc. Natl Acad. Sci. USA, 96, 6797–6801. [DOI] [PMC free article] [PubMed] [Google Scholar]