


Abstract
The quality of germ cell DNA is critical for the fate of the offspring, yet there is limited knowledge of the DNA repair capabilities of such cells. One of the main DNA repair pathways is base excision repair (BER) which is initiated by DNA glycosylases that excise damaged bases, followed by incision of the generated abasic (AP) sites. We have studied human and rat methylpurine-DNA glycosylase (MPG), uracil-DNA glycosylase (UNG), and the major AP endonuclease (HAP1/APEX) in male germ cells. Enzymatic activities and western analyses indicate that these enzymes are present in human and rat male germ cells in amounts that are at least as high as in somatic cells. Minor differences were observed between different cellular stages of rat spermatogenesis and spermiogenesis. Repair of methylated DNA was also studied at the cellular level using the Comet assay. The repair was highly efficient in both human and rat male germ cells, in primary spermatocytes as well as round spermatids, compared to rat mononuclear blood cells or hepatocytes. This efficient BER removes frequently occurring DNA lesions that arise spontaneously or via environmental agents, thereby minimising the number of potential mutations transferred to the next generation.
INTRODUCTION
Male germ cells pass their DNA on to the next generation and an efficient surveillance of the integrity of their genome is crucial for healthy offspring. DNA repair activity in male germ cells may be different from that in somatic cells, since meiosis and spermatogenesis are characterised by a number of changes affecting the structural and functional organisation of the genome. The expression of DNA repair enzymes in male germ cells is largely unknown, whereas in somatic cells expression of certain repair enzymes varies according to the cell type (1–4). In another article in this issue we show that nucleotide excision repair (NER) appears to be suppressed or lacking in male germ cells from rats (5,6). There is reason to believe that other pathways of repair are functional in male germ cells based on the induced unscheduled DNA synthesis (UDS) following treatment with DNA damaging agents (7,8). Furthermore, DNA single-strand and double-strand breaks are efficiently repaired (9–11).
Base excision repair (BER) is a major pathway for removal of DNA lesions arising from endogenous processes as well as those induced by exposure to exogenous chemicals and irradiation (12). BER is initiated by DNA glycosylases that excise aberrant bases from DNA by cleavage of the N-glycosidic bond linking the base to its deoxyribose sugar. DNA glycosylases are specific with regard to their substrate; each enzyme removes only a particular set of DNA lesions. The generated apurinic/apyrimidinic (AP) sites are incised by specific endonucleases, inappropriate ends are removed, the missing bases are filled by DNA polymerases and nicks are ultimately sealed by DNA ligases. The BER pathway involves the replacement of one (short patch BER) or several nucleotides (long patch BER) at the site of the lesion (13).
The DNA of human cells may be alkylated by endogenous methylating agents or by exposure to cigarette smoking, industrial chemicals or cancer chemotherapeutic agents (14,15). In human and rat somatic cells, methylpurine-DNA glycosylase (MPG) catalyses the excision of a variety of modified bases, including N-methylpurines, hypoxanthine (Hx) (16), and 1,N6-ethenoadenine (ɛA) (17,18). MPG has been detected in a wide variety of organisms including man (hMPG) and rat (rMPG) (19,20).
Uracil (U) represents an endogenous DNA lesion arising from deamination of cytosine or incorporation of U instead of T during DNA synthesis (21). Deamination of cytosine leads to U:G mispairs at a frequency of 100–500 events per cell per day, ultimately producing C:G→T:A transitions if not repaired prior to replication (12,22). U residues may also be induced by methotrexate treatment or dietary folate deficiency (23–25). U is predominantly removed by uracil-DNA glycosylase (UNG) (26–28). A number of other enzymes, such as the single-strand-selective monofunctional UNG (SMUG1) (29), also possess U-excising activities, but these seem to play a minor role, although they are important as back-up enzymes in UNG-deficient mice (28–31).
AP sites are formed in high numbers by the spontaneous hydrolysis of bases generating the loss of up to 10 000 purines per day per mammalian cell (32). In addition, AP sites arise by exposure of cells to DNA damaging agents or by removal of bases by DNA glycosylases. They are non-instructional to the DNA polymerases and are potentially pro-mutagenic. They can inhibit DNA synthesis and, as a consequence, induce cell death (33).
We have studied BER in male germ cells from specific stages of spermatogenesis from rats and from human testis biopsies. Here we report on the activities of the DNA repair enzymes MPG, UNG/SMUG1 and AP-endonuclease I (HAP1/APEX) in cellular extracts using DNA substrates containing selected DNA lesions. Furthermore, repair proteins have been quantified by western analysis and repair of alkylation damage in single cells ex vivo was measured by means of the Comet assay. Enriched male germ cell populations and a ploidy-specific Comet assay were used to obtain information about cells of different stages of spermatogenesis. The data demonstrate that BER proteins are present and functional in pre- and post-meiotic germ cells from both humans and rats.
MATERIALS AND METHODS
Chemicals
RPMI 1640 culture medium with 25 mM HEPES buffer, l-glutamine, and fetal calf serum (FCS) were from Gibco BRL (Life Technologies, Paisley, UK). Lymphoprep‘ was from Nycomed-Amersham (Oslo, Norway). Hoechst 33258 and Hoechst 33342 were from Calbiochem-Boehringer (La Jolla, CA). Monoclonal mouse anti-vimentin (clone V9) and FITC-conjugated swine anti-rabbit IgG were from DAKO (Glostrup, Denmark). FITC-conjugated goat anti-mouse IgG was from Becton Dickinson (San Jose, CA). N-[3H]methyl-N′-nitrosourea ([3H]MNU; 18.0 Ci/mmol), [γ-32P]ATP (5000 Ci/mmol), NAP-5 columns and ECL‘ kit were from Amersham Pharmacia Biotech (Buckinghamshire, UK). Oligonucleotides were from Eurogentech (Seraing, Belgium). HRP-conjugated donkey anti-rabbit IgG was from Jackson Laboratories (Bar Harbor, ME). AP-conjugated mouse anti-rabbit IgG and Protoblot® kit were from Promega (Madison, WI), and the ATP Determination Kit was from Molecular Probes (Eugene, OR).
Cells
Cells were isolated and kept in RPMI 1640 medium supplemented with 10% FCS and antibiotics (100 IU/ml penicillin; 0.1 mg/ml streptomycin). The medium for male germ cells was further supplemented with pyruvate (0.1 mg/ml). Male germ cells were prepared from human testis biopsies obtained from organ transplant donors and from rat testes of sexually mature male rats (MOL: WIST, 200–300g; Møllegaard, Ejby, Denmark) by enzymatic digestion with collagenase and trypsin as described (10). This study was conducted in accordance with the Norwegian Law concerning transplantation, hospital autopsies and donation of corpses (revision of Jan. 1, 1994). Viability was >90% as measured by trypan blue exclusion, and for some samples also by staining with Hoechst 33342 (10 µg/ml) and propidium iodide (PI; 10 µg/ml) in RPMI 1640 medium for 30 min at room temperature in the dark. Crude human testicular cell suspensions contained 9% tetraploid cells (±3%) (4C), 25% diploid cells (±7%) (2C) and 64% haploid cells (±9%) (1C), whereas crude rat testicular cell suspensions contained 7% tetraploid cells (±2%), 14% diploid cells (±5%), 60% haploid round spermatids (±7%) and 17% haploid elongating/elongated spermatids (±5%). The human organ donors had a median age and body weight of 32 years and 76 kg, with ranges of 14–60 years and 60–105 kg, respectively. Testis biopsies were kept on ice for 1–10 h before enzymatic digestion of the tissue. Cellular extracts were prepared from testis biopsies of the following human organ donors: (1) 39 years/70 kg; (2) 29 years/80 kg; (3) 14 years/60 kg; (4) 60 years/85 kg; (5) 37 years/95 kg; (6) 23 years/75 kg; (7) 26 years/70 kg; (8) 21 years/76 kg; (9) 34 years/75 kg. Human testicular cells used in Comet assays were from the following donors: (10) 49 years/105 kg, (11) 29 years/75 kg and (12) 57 years/90 kg. The proportion of somatic cells in human and rat testicular cell suspensions was 16% (±7%; n = 5) and 11% (±6%), respectively, whereas fractions enriched in rat spermatocytes or round spermatids contained 7% (±4%) and 9% (±2%) somatic cells, respectively. Blood mononuclear cells (MNC) were isolated from fresh blood taken from human volunteers or male rats by density centrifugation using Lymphoprep‘ as described (34). Primary hepatocytes were prepared from mature male rats by a collagenase perfusion method as described (35). Viability of human MNCs and rat hepatocytes, by trypan blue exclusion, was >90%.
Enrichment and characterisation of male germ cells
Rat testicular cells were separated by centrifugal elutriation (36) as modified by Bjørge et al. (37). Three cell fractions were collected containing predominantly primary spermatocytes, round spermatids or elongating/elongated spermatids, respectively. The elongating/elongated spermatids were further enriched by metrizamide density gradient centrifugation (37).
The composition of testicular cell suspensions stained with 1 µg/ml Hoechst 33258 (37) was analysed using a Bio-Rad Bryte HS™ flow cytometer. The percentage of 1C, 2C and 4C cells was estimated from dual-parameter cytograms and was confirmed by microscopic evaluation of cells stained with Hoechst 33342 and PI. Cell smears were evaluated using a Leica DMLB fluorescence microscope (1000×).
Vimentin is a cytoskeletal protein expressed solely in somatic cells (38), which was used to evaluate the proportion of somatic cells in the testicular cell suspensions. Cells were fixed in 0.1% paraformaldehyde in ice-cold PBS for 30 min followed by permeabilisation with digitonin (10 µg/ml), stained with anti-vimentin (38) and analysed by fluorescence microscopy and dual-parameter flow cytometry. The Synaptonemal Complex Protein 3 (SCP3) is part of the synaptonemal complex in primary spermatocytes, and was used to determine the proportion of primary spermatocytes in testicular cell suspensions. Cells stained with rabbit anti-SCP3 (a gift from Dr Peter de Boer, Wageningen University, Netherlands) followed by FITC-conjugated swine anti-rabbit IgG were scored by fluorescence microscopy.
Preparation of cellular extracts
Extracts were made by plasmolysis as described (39), and their protein contents were determined by the method of Lowry using bovine serum albumin (BSA) as the standard.
Preparation of DNA substrates
A substrate containing 3H-labelled methylated bases was prepared as described (40) with modifications. Calf thymus DNA was treated with [3H]MNU (18 Ci/mmol) in 10 mM sodium cacodylate pH 7.0 for 2 h at 37°C. DNA was precipitated twice with ethanol, washed and dissolved in 10 mM Tris–HCl pH 8/1 mM EDTA (TE), and dialysed in TE for 16 h. Specific activity was 1.5 Ci/mmol.
Duplex DNA substrates containing one defined DNA lesion were prepared essentially as described (41). The DNA lesions were Hx, ɛA, U and tetrahydrofuran (THF). Oligonucleotides containing the lesion were 5′-end labelled by [γ-32P]ATP and T4 polynucleotide kinase (37°C for 30 min), followed by separation from the residual [γ-32P]ATP using NAP-5 columns and hybridisation with complementary oligonucleotides. Duplex DNA was purified by separation by non-denaturing PAGE and electroelution. The DNA sequences of the oligonucleotides were as follows: 5′-GCTCATGCGCAG[Hx]CAGCCGTACTCG, 5′-CGAGTACGGCGG[ɛA]GGGCGCATGAGC, 5′-GTCCATGCGCAG[U]CAGCCGTACTCG, 5′-GCTGTTGAGATCCAGTTCG[THF]AGTAACCCACTCGTGC and complementary oligonucleotides.
DNA glycosylase assays
The reactions were carried out using 1–4 separate extract preparations of each cell type in 70 mM MOPS pH 7.5/1 mM EDTA/1 mM dithiothreitol/5% glycerol for 30 min at 37°C, unless otherwise indicated.
The release of methylated DNA bases was performed as described (40,42). Extracts were incubated with 3H-methylated DNA (300 fmol) in a total volume of 50 µl. The reactions were stopped by adding 75 µl solution A (2 mg calf thymus DNA/80 mg BSA/0.5 M NaAc pH 5.5). DNA was ethanol precipitated, centrifuged, and an aliquot of the supernatant was mixed with 5 ml Aquasafe scintillation fluid. The released 3H-methylated bases were quantified using a liquid scintillation analyser (Packard, Tri-Carb, model 1500).
The oligonucleotide nick assay was carried out as described (41). Extracts were incubated with 32P-labelled substrate (1–5 fmol; 100–2000 c.p.m./reaction) in a total volume of 5 µl. The reactions were terminated by adding 3.5 µl gel loading dye solution (95% formamide/20 mM EDTA pH 8/0.05% bromophenol blue/0.05% xylene cyanol). The cleavage products were denatured (1 min, 80°C) and separated by 20% denaturing PAGE, followed by imaging of the dried gel using a Molecular Dynamics PhosphorImager model 445 SI. The relative amount of cleaved substrate was calculated from the amount of cleaved substrate divided by the total amount of substrate.
Western analysis
Standard western analysis was performed using 20–40 µg extract per lane. Primary antibodies were protein-A purified rabbit anti-MPG serum (1:3125 dilution; the crude serum was a gift from Dr Timothy O’Connor, City of Hope National Medical Center, Duarte, CA), rabbit anti-UNG (1:2000 dilution; a gift from Dr Hans Krokan, Norwegian University of Science and Technology, Trondheim, Norway), polyclonal rabbit anti-HAP1 (1:3000 dilution; a gift from Dr Ian Hickson, ICRF, John Radcliffe Hospital, Oxford, UK) and polyclonal rabbit anti-Ref-1 for the detection of APEX (1:7500 dilution). The secondary antibodies were HRP-conjugated donkey anti-rabbit IgG (1:3000 dilution) or AP-conjugated mouse anti-rabbit IgG (1:5000 dilution). Bands were visualised using ECL™ followed by autoradiography, or by the development of colour (Protoblot®).
DNA repair assays in cells
Cells (4 × 106 cells per sample) were exposed to methyl methanesulfonate (MMS) (in 0.5% DMSO) in 2 ml RPMI 1640 culture medium for 30 min at 32°C (testicular cells) or 37°C (somatic cells). Reactions were placed on ice for termination and diluted with medium. For repair incubation, cells in medium were kept in Petri dishes in a humidified atmosphere at 32°C, 5% CO2 for up to 2 h. Viability was monitored by staining with PI and Hoechst 33342, and was >90%. Rat testicular cells exposed to 0.2 mM MMS and incubated for up to 2 h were also analysed with respect to ATP content, using the ATP Determination Kit. Boiled cells (inactivation of endogenous phosphatases) were lysed by four freeze–thaw cycles, and 10 µl cell lysate was added to an ATP standard solution (90 µl) as recommended by the manufacturer. The samples were left at 28°C for 25 min through the initial chemiluminescence flash period, and chemiluminescence was measured for 30 s in a scintillation counter.
The Comet assay was performed as described, with modifications (37,43–44). In short, slides were prepared as a sandwich consisting of two layers of agarose. The first layer was 100 µl normal melting agarose (1%) and the second was 10 µl cells (8 × 104) mixed with 75 µl low melting agarose (0.7%). Following lysis of cells for 90 min and unwinding of the DNA in alkaline electrophoresis buffer for 40 min at 10°C, electrophoresis was performed for 20 min at 20 V/m. Slides were neutralised and stained with ethidium bromide, and comets were visualised using a Leitz fluorescence microscope (Ortholux II) plus a CCD camera, with the Fenestra Comet version 3 (Kinetic Imaging LTD, Liverpool, UK) image analysis system analysing 50 or 100 comets per slide. Tail moments (the percentage of DNA in the tail multiplied by the tail length, measured from the centre of the nucleus), were used as a quantitative measure of the DNA damage. The total fluorescence (proportional to the DNA content of each comet) was recorded to allow analysis of cells of different ploidy (1C ∼spermatids; 2C ∼Sertoli cells, Leydig cells, secondary spermatocytes, leukocytes; 4C ∼primary spermatocytes).
RESULTS
Enrichment and characterisation of cells
All human testis biopsies revealed normal spermatogenesis (1C > 2C > 4C) as determined by flow cytometry (45). Enriched rat male germ cell populations were characterised by flow cytometry and microscopic analysis and contained 50% spermatocytes (±15%), 93% round spermatids (±4%) and 94% elongating/elongated spermatids (±2%). The proportion of rat primary spermatocytes identified as SCP3-positive cells in crude rat testicular cell suspensions was 11% (±4%; n = 7), whereas fractions enriched with primary spermatocytes or round spermatids contained 46% (±8%; n = 5) and 1% (±0%; n = 3) SCP3-positive cells, respectively. The viability of cells was >90%.
MPG
Substrates containing methylated bases, ɛA or Hx were used to study MPG activity. All extracts from human and rat male germ cells were able to excise methylated bases (Fig. 1A). The excision activities were similar in all the enriched rat male germ cells, whereas rat hepatocytes showed 2–3-fold lower activities. Human testicular cells exhibited ∼75% higher activity compared to human MNCs. The excision of ɛA and Hx, both base-paired with T, was measured as cleavage of 32P-labelled linear double-stranded oligonucleotides containing single DNA lesions (oligonucleotide nick assay) (Fig. 1). Rat male germ cells exhibited 3–7-fold higher ɛA removal than hepatocytes per µg cellular extract, with primary spermatocytes showing the highest activity (Fig. 1A and B). A similar pattern of incision activities was observed in the rat for Hx (Fig. 1A and C). Human testicular cells exhibited very low activities for Hx-removal when enzymatic activity per µg extract was compared between human and rat testicular extracts, even when 5-fold more human than rat extracts were used. These differences were not related to cell viability. The removal of Hx base-paired with A, C or G was also measured in rat male germ cells, and was much lower than for Hx base-paired with T (data not shown).
Figure 1.

MPG activity in male germ cell extracts. (A) Release of N-methylated purines, ɛA and Hx by extracts from different cell populations. Methylated bases: excision of 3H-methylated bases from DNA measured as a function of the amount of extract [mean ± SD, background subtracted; 25 µg rat or 10 µg human extract (organ donors 1–4)]. ɛA and Hx: incision of defined DNA lesions calculated as percent cleaved linear double-stranded oligonucleotide containing one defined ɛA or Hx residue, respectively (mean ± SD, or 1 measurement only where no SD is given). The amount of extract was 3 µg for incision of ɛA, and 1 and 5 µg for rat and human extracts (donors 5–7), respectively, for the incision of Hx. ND = no data. Three independent extract preparations of each cell type were used unless otherwise stated. Abbreviations: TC, testicular cells; SC, spermatocytes; RS, round spermatids; ES, elongating/elongated spermatids; HEP, primary hepatocytes; MNC, blood mononuclear cells. Rat or human samples are indicated with r or h, respectively. (B) Image of gel showing the incision of ɛA as described in (A). Each lane represents one distinct extract. Upper band, substrate; lower band, cleavage product. Controls: lane 1, buffer alone; lane 2, E.coli alkylbase-DNA glycosylase II (AlkA) and Formamidopyrimidine-DNA glycosylase (Fpg); lane 3, MPG and Fpg; lane 4, Fpg. Abbreviations as in (A). (C) As in (B), but the substrate contains Hx. Lane 1, buffer alone; lane 2, AlkA and Nfo; lane 3, Nfo.
A prerequisite for the detection of monofunctional DNA glycosylase activity in the oligonucleotide nick assay is the subsequent incision of AP sites. Since the assays are conducted in the presence of EDTA (no Mg2+), the only functional incision activities in the extracts are AP-lyase activities such as those of bifunctional DNA glycosylases, and their presence may thus be rate limiting in the oligonucleotide nick assay. Therefore, an excess amount of purified Escherichia coli endonuclease IV (Nfo) was added to each reaction (Table 1). With Nfo present, the amount of extract required for cleavage of Hx–DNA decreased 2-fold for rat and 10-fold for human extracts when compared to reactions without Nfo (Table 1). The relative MPG activities were up to 5-fold higher in rats compared to humans. No major relative differences between the various rat male germ cells were observed except for somewhat higher Hx removal by round spermatids. With respect to somatic cells, the Hx removal of rat hepatocytes was similar to that of rat testicular cells. Furthermore, in the presence of Nfo both human testicular and MNC extracts exhibited removal of Hx (Table 1), in contrast to the non-detectable activity in the absence of Nfo (Fig. 1A and B). The addition of Nfo resulted in increased cleavage of ɛA DNA. With excess Nfo it was apparent that the incision rates for ɛA among the various human and rat extracts resembled that for Hx, and also that for the excision of methylated bases.
Table 1. Cleavage of ɛA and Hx in linear double-stranded DNA by different human and rat cellular extracts in the presence of Nfo.
| Substrate | Rata | Humana | |||||
| |
rTC |
rSC |
rRS |
rES |
rHEP |
hTC |
hMNC |
| ɛA (%)b | 20 ± 4 | 20 ± 4 | 29 ± 6 | 19 ± 4 | 18 ± 2 | 7 ± 3 | 8 ± 2 |
| Hx (%)b | 16 ± 5 | 13 ± 1 | 26 ± 2 | 19 ± 5 | 22 ± 5 | 5 ± 1 | 9 ± 3 |
aAt least three independent cellular extracts (0.5 µg protein per reaction) of a defined cell origin were tested. Nfo was added in excess to each reaction (10 ng, rat; 6 ng, human).
bData are represented as mean percent cleavage of the substrate (±SD). For abbreviations see legend to Figure 1.
Both human and rat male germ cell extracts exhibited MPG as detected by western analysis (Fig. 2). A strong single band at ∼39 kDa was detected in extracts from the enriched fractions of rat male germ cells, demonstrating high expression of MPG (Fig. 2; upper panel). The results indicate that round spermatids express slightly higher amounts of MPG than the other cell types, which was also seen by enzymatic assays in the presence of Nfo (Table 1). Rat hepatocytes, on the other hand, expressed very low levels of MPG compared to rat male germ cells. Human extracts exhibited staining of two proteins of different molecular weight, at ∼32 and ∼40 kDa (Fig. 2; lower panel). Their expression varied among the six human testicular extracts, but the expression was, in all cases, substantially higher than in human MNCs. MPG is thus expressed in both human and rat male germ cells, and the expression is higher than in the somatic cell types tested. We note here that extended time periods between dissection of human testis biopsies and preparation of extracts did not lead to degradation of MPG, and that storage of cellular extracts at –80°C for different time periods did not affect MPG activity.
Figure 2.
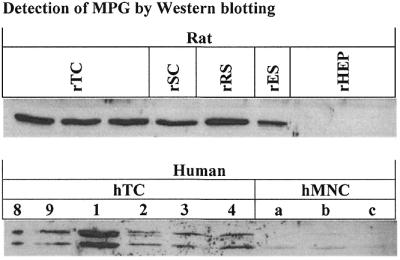
Detection of MPG by western analysis. Upper panel, rat extracts; lower panel, human extracts. Each lane represents one independent extract of each cell type. Human testicular extracts are denoted with donor numbers as described in Materials and Methods and human MNC extracts with letters (a–c). Abbreviations are described in the legend to Figure 1.
U excision and the presence of UNG
Extracts from different rat male germ cell types, and also rat hepatocytes, showed similar incision rates for U from U:G in the oligonucleotide nick assay (Fig. 3A). The extracts also incised substrates with U base-paired with A, T or C (data not shown). Three separate human testicular extracts cleaved 25–52% of a U:G-substrate, compared to 40% by human MNCs (Fig. 3B). In general, less human than rat extract was needed to cleave U:G. Furthermore, the addition of Nfo led to increased cleavage of U:G per µg extract, both for human and rat male germ cells (data not shown). Western analyses showed the presence of UNG in both human and rat male germ cells (Fig. 3C). The different rat male germ cells exhibited approximately similar amounts of the mitochondrial UNG1, compared to lower amounts in rat hepatocytes. Rat hepatocytes exhibited staining of a protein of higher molecular weight (38 kDa), which probably represents the cytoplasmic pre-version of UNG (46). The different human testicular extracts exhibited variable expression of the mitochondrial UNG1, yet they were higher than in human MNCs. Human extracts also expressed variable amounts of the nuclear UNG2, which was non-detectable in rat extracts.
Figure 3.
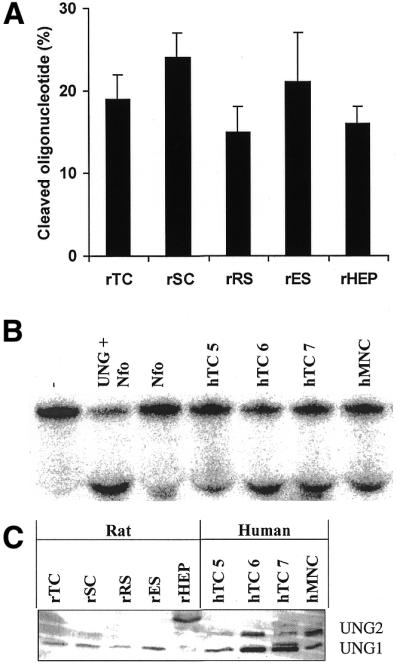
Uracil excision by male germ cell extracts. (A) Incision of U from a linear double-stranded oligonucleotide containing one U residue base-paired with G measured as percent cleaved substrate (mean ± SD). Three independent rat extract preparations (each 1 µg) of each cell type were tested. (B) Incision of U:G as described in (A). Three independent human testicular extracts (donors 5–7) or one human mononuclear cell extract (0.25 µg each) were tested. Lanes from the left: –, no extract (buffer alone); UNG + Nfo, E.coli UNG (1 U) in combination with Nfo (10 ng) as a positive control; Nfo, Nfo (10 ng; determination of the level of AP sites in the substrate). Upper band, substrate; lower band, cleavage product. (C) Detection of UNG in rat and human (donors 5–7) extracts by western analysis. The positions of the nuclear (UNG2) and mitochondrial (UNG1) UNG are indicated. Abbreviations are as described in the legend to Figure 1.
AP endonuclease
AP sites are processed by AP endonucleases and the major enzymatic activities in humans and rats are HAP1 and APEX, respectively. The AP endonuclease activity was measured using the oligonucleotide nick assay in the presence of Mg2+, utilising a substrate containing a single synthetic AP site, THF, which functions as a specific DNA substrate for detecting AP-endonuclease activity (47). The different rat male germ cell extracts exhibited similar levels of incision activity, whereas in rat hepatocytes they were 2-fold higher (Fig. 4A). Human testicular extracts exhibited higher activities than rat extracts, although the activity varied between the different human extracts (Fig. 4A). Furthermore, human testicular cells showed similar levels of activity as human MNCs. The specific presence of the 37 kDa HAP1 or APEX in both human and rat male germ cells, respectively, were demonstrated by western analyses (Fig. 4B). The expression of HAP1 in human testicular cells was variable but always higher than in human MNCs. Rat hepatocytes expressed very low amounts of APEX at the expected molecular weight, but there was an additional band at a higher molecular weight. This may represent an alternative version of APEX.
Figure 4.
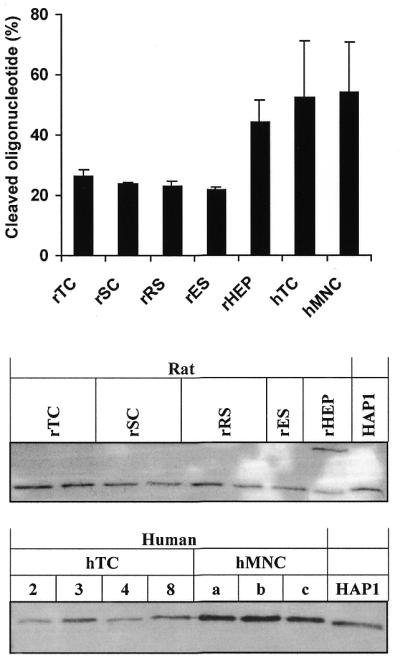
AP endonuclease in male germ cell extracts. (A) Incision of linear double-stranded oligonucleotide containing one THF residue calculated as percent cleaved substrate (mean ± SD). Three independent extract preparations (1 ng each) of each cell type were tested; human testicular extracts (donors 1–4). Nfo (5 ng) was used as a positive control. (B) Rat APEX (upper panel) and human HAP1 (lower panel) detected by western analysis. Purified HAP1 (20 ng) was used as positive control. Human testis biopsies are denoted with numbers and human MNCs with letters. Abbreviations are as described in the legend to Figure 1.
Cellular repair of MMS-induced DNA lesions
The enzymatic activities detected in extracts and western analysis consistently showed that human and rat male germ cells contain functional MPG, UNG/SMUG1 and HAP1/APEX. As a complementary approach we studied repair at the cellular level following treatment with MMS using the Comet assay. The cell types studied showed linear dose–effect relationships (data not shown). Rat male germ cells exhibited markedly higher tail moments than the other cell types, when exposed to similar doses of MMS, with 5-fold higher tail moments than human testicular cells at 0.4 mM MMS. To obtain similar levels of initial DNA lesions, the various cell populations were treated with different concentrations of MMS. Efficient repair of methylated DNA was demonstrated in male germ cells from rats and three different humans, and the repair kinetics were broadly similar between the two species (Fig. 5A). Within 2 h, these cells repaired 60–80% of the initial DNA damage. The energy level of rat male germ cells was monitored by measuring the cellular ATP level, and was sustained throughout the 2 h repair periods (data not shown). The repair rates of MNCs and hepatocytes from rats were markedly lower compared to rat male germ cells (Fig. 5B). The Comet assay allows analysis of cells of different ploidy in heterogeneous male germ cell suspensions, thereby facilitating the investigation and comparison of repair capacity between distinct spermatogenic cell types. Grouping the results accordingly demonstrated rapid repair by both rat primary spermatocytes and round spermatids (Fig. 5C). Efficient repair was also consistently observed in the two cell types from human testis biopsies (Fig. 5D). Round spermatids displayed somewhat higher initial tail moments than primary spermatocytes exposed to similar concentrations of MMS both in humans and in rats (Fig. 5C and D). However, we have preliminary results showing that primary spermatocytes have apparently 25% smaller tail moments than round spermatids at similar doses of X-rays, which induce DNA lesions uniformly (unpublished data).
Figure 5.

Induction and repair of DNA lesions in different cells exposed to MMS. DNA damage (tail moment; mean ± SD) was measured with the Comet assay. (A) Repair of MMS-induced DNA lesions in human and rat male germ cells. Rat testicular cells were exposed to 0.2 mM MMS, whereas human testicular cells (donors 10–12) were exposed to 0.2, 0.6 and 0.8 mM MMS, respectively, and incubated for up to 2 h to allow repair. The initial tail moment is defined as 100%. Broken line with diamonds shows rat testicular cells; solid lines with squares, triangles and crosses are human testicular cells from human donors 10, 11 and 12, respectively. (B) Repair of MMS-induced DNA lesions in testicular compared to somatic cells. Rat testicular cells, MNCs and hepatocytes were treated with 0.2, 0.5 and 0.5 mM MMS, respectively and analysed as in (A). Diamonds, squares and triangles represent testicular cells, hepatocytes and MNCs, respectively. Filled symbols show exposed cells; open symbols show non-exposed cells (controls). (C) Repair of MMS-induced DNA lesions in rat male germ cell of different spermatogenic stages. Rat testicular cells were exposed to 0.2 mM MMS, analysed as in (A) and grouped according to ploidy: haploid, round spermatids; predominantly tetraploid primary spermatocytes. Diamonds and triangles represent round spermatids and primary spermatocytes, respectively. Filled symbols show exposed cells; open symbols show non-exposed cells (control). (D) Repair of MMS-induced DNA lesions in different human spermatogenic stages. Conditions were similar as in (C) except that the cells were exposed to 0.8 mM MMS. One representative experiment out of three is shown.
DISCUSSION
Cellular repair
Both human and rat male germ cells efficiently repair MMS-induced DNA lesions with similar repair rates. Furthermore, different spermatogenic cell types, primary spermatocytes and round spermatids, repair MMS-induced DNA lesions. BER thus seems to be proficient throughout spermatogenesis and spermiogenesis. It cannot be excluded, however, that the Comet assay may show repair by other pathways besides BER. The functionality of BER is supported by the presence and activity of BER-related enzymes in human testicular extracts and extracts from the enriched rat male germ cells. UDS in MMS-exposed rat spermatocytes has been shown to be 4–25-fold higher in situ in cultured seminiferous tubules, in which germ cells maintain their intimate relationship with Sertoli cells, than spermatocytes in suspension (48), indicating that repair may be even more efficient in vivo. The repair rates of male germ cells were markedly higher compared to MNCs and hepatocytes. This is consistent with the lower excision activities and expression of MPG detected by western analysis in somatic compared with male germ cell extracts. More efficient BER in male germ cells compared to somatic cells corresponds to the lower spontaneous mutation frequencies of mouse meiotic cells compared to somatic cells (49). Furthermore, it is consistent with the lack of increased mutation rates in male germ cells of Big Blue transgenic mice treated with MMS (50).
Methylated bases are thought to induce mutations indirectly via the generation of AP sites (51). MMS did not induce mutations in male germ cells of Big Blue transgenic mice even at doses that were lethal to the animal, suggesting efficient DNA repair of the methylated DNA or of the AP sites, although large deletions are not scored by this assay. Methylated bases are of nature labile and will be converted into pre-mutagenic AP sites, and prior to differentiation into spermatozoa, at which stage it is assumed that no repair occurs, efficient repair of such DNA lesions hence reduces the risk of inducing germline mutations. In accordance with the observed cellular repair, extracts of both human and rat male germ cells exhibit activities for the removal of all the DNA substrates tested in this study, and the activities were found to be similar in the different populations of enriched rat male germ cells. Furthermore, the results obtained by western analysis correspond well with the enzymatic activities detected within each species. The variable expression of BER enzymes observed in humans indicates inter-individual differences, which according to our results do not correlate with age, but may reflect genetic variation. Alternatively, such differences may in part be explained by lifestyle factors such as smoking, since MPG is induced to higher levels in smokers compared to non-smokers (52).
MPG
The two assays used in this study to measure MPG activity are functionally different; the release of methylated bases is measured directly whereas the excision of ɛA and Hx requires subsequent incision. The incision capacity (AP-lyase activity) of extracts may thus be rate-limiting when measuring the removal of ɛA and Hx. The changes in activity with or without Nfo present indicate that human male germ cells and rat hepatocytes contain less AP-lyase activity than rat male germ cells. The increased incision observed in the presence of Nfo may reflect a stimulated release of MPG from AP sites by an AP endonuclease. This has been shown previously for several other DNA glycosylases including UNG, thymine-DNA glycosylase (TDG), SMUG1, mammalian adenine-DNA glycosylase (MYH), and human 8-oxoguanine-DNA glycosylase (hOGG1) (28,53–56). The higher MPG activity of rat testicular compared to liver extracts was consistent with previous studies with mice (57,58). The observed differences in MPG activity between rat and human male germ cell extracts are in accordance with the mouse MPG having 2–3-fold higher rates of removal of 3mA and 7mG compared to the human MPG in vitro (59). Alternatively, the presence of other BER-related proteins may influence the excision/incision rate. One such protein is the human homologue (hHR23) of Rad23, which stimulates the removal of Hx when associated with MPG, and therefore has been proposed as a general DNA damage recognition protein involved in both NER and BER (60). A recent observation by Kartalou et al. (61) represents another interesting coupling of MPG with NER, in which MPG binds to cisplatin-adducts, representing lesions that are repaired via NER. The high amounts of MPG in male germ cells may thus interfere with a possible residual NER capacity in these cells (6), contributing to the highly efficient treatment of testicular cancer using cisplatin.
U excision and the presence of UNG
Human and rat male germ cells incised U from the U:G-substrate in vitro, and the activity may be ascribed to UNG and/or SMUG1 (28). UNG plays an important role for the removal of U when base-paired with guanine since ∼50% of the activity is lost in UNG-deficient mice (28). The residual activity of such UNG-deficient mice was not inhibited by the Ugi peptide inhibitor, suggesting that SMUG1 is responsible as back-up enzyme (28). Incision of U in male germ cells was similar to that of somatic cells. As with MPG, the increased cleavage observed in the presence of Nfo could be explained by stimulation of the DNA glycosylase activity in the presence of an AP endonuclease. Both UNG and SMUG1 have been shown to be stimulated by HAP1 (28,53,62). Western analyses confirmed the presence of UNG in human and rat male germ cells, and we propose that the mitochondrial UNG1 is the predominant version in male germ cells in both humans and rats.
The expression of mRNA for the mitochondrial UNG1 and the nuclear UNG2, both encoded by the same gene, has been measured in various tissues including the testis, which exhibited the highest levels for both (63). The UNG activity has been reported to be high (per cell number) in DNA- synthesising mouse male germ cells (spermatogonia and preleptotene spermatocytes); meiotic and post-meiotic cells exhibit markedly lower levels of UNG activity with a relative maximum in pachytene spermatocytes (64). This is in accordance with our results, since the primary spermatocytes have a higher UNG protein content than the spermatids on a per cell basis, due to the larger size and the higher total DNA and protein content of spermatocytes. The nuclear UNG2, partly localised in replication factories, removes U as it is incorporated into the genome (3). The enriched male germ cell populations are non-replicating and hence the need for the replication-associated repair by UNG2 is limited. Another function of UNG in male germ cells may be protection from cytotoxic AP sites, since UNG binds more tightly and rapidly to AP sites than to U in DNA (53). It hence appears that male germ cells repair U. This conclusion relies on the observed repair of methylated DNA, combined with the presence of U excising activities in extracts, and UNG detected by western analysis. Additional support for this is the observation that a 180 kDa protein complex isolated from bovine testis is able to conduct the complete U-initiated BER reaction in vitro (65).
HAP1/APEX
The AP-endonuclease activity was higher in human than in rat male germ cells, different from the results obtained with both MPG and UNG/SMUG1. Furthermore, rat male germ cells exhibit lower amounts of AP-endonuclease activity than rat hepatocytes. Since male germ cells were able to repair methylated DNA rapidly, the amount of HAP1/APEX seems sufficient for BER under the conditions employed. The level of APEX mRNA expression is reported to be highest in the testis and lowest in the liver of the various organs examined in the rat (66). APEX mRNA is present at modest levels in spermatogonia and spermatocytes, whereas the level in round spermatids is higher (67). HAP1 has a number of other enzymatic functions besides its AP-endonuclease activity. The expression of APEX mRNA is developmentally regulated in the rat testis indicating involvement in processes such as recombination (66), and HAP1 is localised in the cytoplasm of spermatocytes unlike the nuclear localisation of most other tissues (68,69).
In summary, MPG, UNG and HAP1/APEX are expressed in human and rat male germ cells, and are highly functional towards selected substrates in vitro. Furthermore, male germ cells in suspension efficiently repair methylated DNA, indicating functional and efficient BER. Human and rat male germ cells thus appear to be well protected against some of the most common DNA lesions.
Acknowledgments
ACKNOWLEDGEMENTS
We wish to thank the Department of Surgery and the transplantation team at the National Hospital, Oslo, Norway, for providing the human testis biopsies. Furthermore, we would like to thank Dr Timothy O’Connor for the MPG serum, Dr Hans Krokan for the UNG antibody, Dr Ian Hickson for the HAP1 antibody and Dr Peter de Boer for the SCP3 antibody. We also wish to thank Lena M. Jensen and Kirsti Haug for excellent technical assistance. E.S. and M.B. acknowledge support from the Norwegian Cancer Society and Anders Jahres Medical Foundation. The study was supported by the EU-Environment and Climate program (contract no. ENV4-CT95-0204) and the Norwegian Research Council (grant no. 129614/310).
References
- 1.Bouziane M., Miao,F., Bates,S.E., Somsouk,L., Sang,B., Denissenko,M. and O’Connor,T.R. (2000) Promoter structure and cell cycle dependent expression of the human methylpurine-DNA glycosylase gene. Mutat. Res., 461, 15–29. [DOI] [PubMed] [Google Scholar]
- 2.Luna L., Bjørås,M., Hoff,E., Rognes,T. and Seeberg,E. (2000) Cell-cycle regulation, intracellular sorting and induced overexpression of the human NTH1 DNA glycosylase involved in removal of formamidopyrimidine residues from DNA. Mutat. Res., 460, 95–104. [DOI] [PubMed] [Google Scholar]
- 3.Otterlei M., Warbrick,E., Nagelhus,T.A., Haug,T., Slupphaug,G., Akbari,M., Aas,P.A., Steinsbekk,K., Bakke,O. and Krokan,H.E. (1999) Post-replicative base excision repair in replication foci. EMBO J., 18, 3834–3844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Helland D.E., Male,R., Haukanes,B.I., Olsen,L., Haugan,I. and Kleppe,K. (1987) Properties and mechanism of action of eukaryotic 3-methyladenine-DNA glycosylases. J. Cell Sci., 6 (Suppl.), 139–146. [DOI] [PubMed] [Google Scholar]
- 5.Brunborg G., Holme,J.A. and Hongslo,J.K. (1995) Inhibitory effects of paracetamol on DNA repair in mammalian cells. Mutat. Res., 342, 157–170. [DOI] [PubMed] [Google Scholar]
- 6.Jansen J., Olsen,A.K., Wiger,R., Naegeli,H., de Boer,P., van der Hoeven,F., Holme,J.A., Brunborg,G. and Mullenders,L. (2001) Nucleotide excision repair in rat male germ cells: low level of repair in intact cells contrasts with high dual incision activity in vitro. Nucleic Acids Res., 29, 1791–1800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Beikirch H. (1977) Induction of unscheduled DNA synthesis by chemical mutagens in testicular cells of the mouse in vitro. Arch. Toxicol., 37, 195–201. [DOI] [PubMed] [Google Scholar]
- 8.Zbinden G. (1980) Unscheduled DNA synthesis in the testis, a secondary test for the evaluation of chemical mutagens. Arch. Toxicol., 46, 139–149. [DOI] [PubMed] [Google Scholar]
- 9.Ono T. and Okada,S. (1976) Radiation-induced DNA scissions and their rejoining in testicular cells of mouse. Mutat. Res., 36, 213–222. [DOI] [PubMed] [Google Scholar]
- 10.Bjørge C., Wiger,R., Holme,J.A., Brunborg,G., Scholz,T., Dybing,E. and Søderlund,E.J. (1996) DNA strand breaks in testicular cells from humans and rats following in vitro exposure to 1,2-dibromo-3-chloropropane (DBCP). Reprod. Toxicol., 10, 51–59. [DOI] [PubMed] [Google Scholar]
- 11.Sathees C.R. and Raman,M.J. (1999) Mouse testicular extracts process DNA double-strand breaks efficiently by DNA end-to-end joining. Mutat. Res., 433, 1–13. [DOI] [PubMed] [Google Scholar]
- 12.Krokan H.E., Nilsen,H., Skorpen,F., Otterlei,M. and Slupphaug,G. (2000) Base excision repair of DNA in mammalian cells. FEBS Lett., 476, 73–77. [DOI] [PubMed] [Google Scholar]
- 13.Klungland A. and Lindahl,T. (1997) Second pathway for completion of human DNA base excision-repair: reconstitution with purified proteins and requirement for DNase IV (FEN1). EMBO J., 11, 3341–3348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Barrows L.R. and Magee,P.N. (1982) Nonenzymatic methylation of DNA by S-adenosylmethionine in vitro. Carcinogenesis, 3, 349–351. [DOI] [PubMed] [Google Scholar]
- 15.Rydberg B. and Lindahl,T. (1982) Nonenzymatic methylation of DNA by the intracellular methyl group donor S-adenosyl-l-methionine is a potentially mutagenic reaction. EMBO J., 1, 211–216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Saparbaev M. and Laval,J. (1994) Excision of hypoxanthine from DNA containing dIMP residues by the Escherichia coli, yeast, rat and human alkylpurine DNA glycosylases. Proc. Natl Acad. Sci. USA, 91, 5873–5877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Singer B., Antoccia,A., Basu,A.K., Dosanjh,M.K., Fraenkel-Conrat,H., Gallagher,P.E., Kusmierek,J.T., Qiu,Z.H. and Rydberg,B. (1992) Both purified human 1,N6-ethenoadenine-binding protein and purified human 3-methyladenine-DNA glycosylase act on 1,N6-ethenoadenine and 3-methyladenine. Proc. Natl Acad. Sci. USA, 89, 9386–9390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Saparbaev M., Kleibl,K. and Laval,J. (1995) Escherichia coli, Saccharomyces cerevisiae, rat and human 3-methyladenine DNA glycosylases repair 1,N6-ethenoadenine when present in DNA. Nucleic Acids Res., 23, 3750–3755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lindahl T. and Wood,R.D. (1999) Quality control by DNA repair. Science, 286, 1897–1905. [DOI] [PubMed] [Google Scholar]
- 20.O’Connor T.R. and Laval,J. (1990) Isolation and structure of a cDNA expressing a mammalian 3-methyladenine-DNA glycosylase. EMBO J., 10, 3337–3342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lindahl T. (1993) Instability and decay of the primary structure of DNA. Nature, 362, 709–715. [DOI] [PubMed] [Google Scholar]
- 22.Friedberg E.C. (1995) Out of the shadows and into the light: the emergence of DNA repair. Trends Biochem. Sci., 20, 381. [DOI] [PubMed] [Google Scholar]
- 23.Goulian M., Bleile,B. and Tseng,B.Y. (1980) Methotrexate-induced misincorporation of uracil into DNA. Proc. Natl Acad. Sci. USA, 77, 1956–1960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Blount B.C., Mack,M.M., Wehr,C.M., MacGregor,J.T., Hiatt,R.A., Wang,G., Wickramasinghe,S.N., Everson,R.B. and Ames,B.N. (1997) Folate deficiency causes uracil misincorporation into human DNA and chromosome breakage: implications for cancer and neuronal damage. Proc. Natl Acad. Sci. USA, 94, 3290–3295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Duthie S.J. and Hawdon,A. (1998) DNA instability (strand breakage, uracil misincorporation and defective repair) is increased by folic acid depletion in human lymphocytes in vitro. FASEB J., 12, 1491–1497. [PubMed] [Google Scholar]
- 26.Slupphaug G., Eftedal,I., Kavli,B., Bharati,S., Helle,N.M., Haug,T., Levine,D.W. and Krokan,H.E. (1995) Properties of a recombinant human uracil-DNA glycosylase from the UNG gene and evidence that UNG encodes the major uracil-DNA glycosylase. Biochemistry, 34, 128–138. [DOI] [PubMed] [Google Scholar]
- 27.Nilsen H., Steinsbekk,K.S., Otterlei,M., Slupphaug,G., Aas,P.A. and Krokan,H.E. (2000) Analysis of uracil-DNA glycosylases from the murine Ung gene reveals differential expression in tissues and in embryonic development and a subcellular sorting pattern that differs from the human homologues. Nucleic Acids Res., 28, 2277–2285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Nilsen H., Rosewell,I., Robins,P., Skjelbred,C.F., Andersen,S., Slupphaug,G., Daly,G., Krokan,H.E., Lindahl,T. and Barnes,D.E. (2000) Uracil-DNA glycosylase (UNG)-deficient mice reveal a primary role of the enzyme during DNA replication. Mol. Cell, 6, 1059–1065. [DOI] [PubMed] [Google Scholar]
- 29.Haushalter K.A., Todd Stukenberg,M.W., Kirschner,M.W. and Verdine,G.L. (1999) Identification of a new uracil-DNA glycosylase family by expression cloning using synthetic inhibitors. Curr. Biol., 9, 174–185. [DOI] [PubMed] [Google Scholar]
- 30.Muller S.J. and Caradonna,S. (1991) Isolation and characterization of a human cDNA encoding uracil-DNA glycosylase. Biochim. Biophys. Acta, 1088, 197–207. [DOI] [PubMed] [Google Scholar]
- 31.Neddermann P., Gallinari,P., Lettieri,T., Schmid,D., Truong,O., Hsuan,J.J., Wiebauer,K. and Jiricny,J. (1996) Cloning and expression of human G/T mismatch-specific thymine-DNA glycosylase. J. Biol. Chem., 271, 12767–12774. [DOI] [PubMed] [Google Scholar]
- 32.Lindahl T. and Nyberg,B. (1972) Rate of depurination of native deoxyribonucleic acid. Biochemistry, 11, 3610–3618. [DOI] [PubMed] [Google Scholar]
- 33.Wallace S.S. (1988) AP endonucleases and DNA glycosylases that recognize oxidative DNA damage. Environ. Mol. Mutagen., 12, 431–477. [DOI] [PubMed] [Google Scholar]
- 34.Hongslo J.K., Brunborg,G., Steffensen,I.L. and Holme,J.A. (1993) Paracetamol inhibits UV-induced DNA repair in resting human mononuclear blood cells in vitro. Mutagenesis, 8, 423–429. [DOI] [PubMed] [Google Scholar]
- 35.Holme J.A., Soderlund,E. and Dybing,E. (1983) Drug metabolism activities of isolated rat hepatocytes in monolayer culture. Acta Pharmacol. Toxicol ., 52, 348–356. [DOI] [PubMed] [Google Scholar]
- 36.Meistrich M.L., Longtin,J., Brock,W.A., Grimes,S.R.,Jr and Mace,M.L. (1981) Purification of rat spermatogenic cells and preliminary biochemical analysis of these cells. Biol. Reprod., 25, 1065–1077. [DOI] [PubMed] [Google Scholar]
- 37.Bjørge C., Wiger,R., Holme,J.A., Brunborg,G., Andersen,R., Dybing,E. and Søderlund,E.J. (1995) In vitro toxicity of 1,2-dibromo-3-chloropropane (DBCP) in different testicular cell types from rats. Reprod. Toxicol., 9, 461–473. [DOI] [PubMed] [Google Scholar]
- 38.Suter L., Koch,E., Bechter,R. and Bobadilla,M. (1997) Three-parameter flow cytometric analysis of rat spermatogenesis. Cytometry, 27, 161–168. [DOI] [PubMed] [Google Scholar]
- 39.Bjelland S., Eide,L., Time,R.W., Stote,R., Eftedal,I., Volden,G. and Seeberg,E. (1995) Oxidation of thymine to 5-formyluracil in DNA: mechanisms of formation, structural implications and base excision by human cell free extracts. Biochemistry, 34, 14758–14764. [DOI] [PubMed] [Google Scholar]
- 40.Riazuddin S. and Lindahl,T. (1978) Properties of 3-methyladenine-DNA glycosylase from Escherichia coli. Biochemistry, 17, 2110–2118. [DOI] [PubMed] [Google Scholar]
- 41.Eide L., Bjørås,M., Pirovano,M., Alseth,I., Berdal,K.G. and Seeberg,E. (1996) Base excision of oxidative purine and pyrimidine DNA damage in Saccharomyces cerevisiae by a DNA glycosylase with sequence similarity to endonuclease III from Escherichia coli. Proc. Natl Acad. Sci. USA, 93, 10735–10740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bjelland S. and Seeberg,E. (1987) Purification and characterization of 3-methyladenine DNA glycosylase I from Escherichia coli. Nucleic Acids Res., 15, 2787–2801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Singh N.P., McCoy,M.T., Tice,R.R. and Schneider,E.L. (1988) A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res., 175, 184–191. [DOI] [PubMed] [Google Scholar]
- 44.Olive P.L., Wlodek,D., Durand,R.E. and Banath,J.P. (1992) Factors influencing DNA migration from individual cells subjected to gel electrophoresis. Exp. Cell Res., 198, 259–267. [DOI] [PubMed] [Google Scholar]
- 45.Kaufman D.G. and Nagler,H.M. (1987) Aspiration flow cytometry of the testes in the evaluation of spermatogenesis in the infertile male. Fertil. Steril., 48, 287–291. [DOI] [PubMed] [Google Scholar]
- 46.Slupphaug G., Markussen,F.H., Olsen,L.C., Aasland,R., Aarsaether,N., Bakke,O., Krokan,H.E. and Helland,D.E. (1993) Nuclear and mitochondrial forms of human uracil-DNA glycosylase are encoded by the same gene. Nucleic Acids Res., 21, 2579–2584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Takeshita M., Chang,C.N., Johnson,F., Will,S. and Grollman,A.P. (1987) Oligodeoxynucleotides containing synthetic abasic sites. Model substrates for DNA polymerases and apurinic/apyrimidinic endonucleases. J. Biol. Chem., 262, 10171–10179. [PubMed] [Google Scholar]
- 48.Bentley K.S. and Working,P.K. (1988) Use of seminiferous tubule segments to study stage specificity of unscheduled DNA synthesis in rat spermatogenic cells. Environ. Mol. Mutagen., 12, 285–297. [DOI] [PubMed] [Google Scholar]
- 49.Walter C.A., Intano,G.W., McCarrey,J.R., McMahan,C.A. and Walter,R.B. (1998) Mutation frequency declines during spermatogenesis in young mice but increases in old mice. Proc. Natl Acad. Sci. USA, 95, 10015–10019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ashby J., Gorelick,N.J. and Shelby,M.D. (1997) Mutation assays in male germ cells from transgenic mice: overview of study and conclusions. Mutat. Res., 388, 111–122. [DOI] [PubMed] [Google Scholar]
- 51.Elder R.H., Jansen,J.G., Weeks,R.J., Willington,M.A., Deans,B., Watson,A.J., Mynett,K.J., Bailey,J.A., Cooper,D.P., Rafferty,J.A., Heeran,M.C., Wijnhofen,S.W.P., van Zeeland,A.A. and Margison,G. (1998) Alkylpurine-DNA-glycosylase knockout mice show increased susceptibility to mutations by methyl methanesulfonate. Mol. Cell. Biol., 18, 5828–5837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hall J., Bresil,H., Donato,F., Wild,C.P., Loktionova,N.A., Kazanova,O.I., Komyakov,I.P., Lemekhov,V.G., Likhachev,A.J. and Montesano,R. (2000) Alkylation and oxidative-DNA damage repair activity in blood leukocytes of smokers and non-smokers. Int. J. Cancer, 54, 728–733. [DOI] [PubMed] [Google Scholar]
- 53.Parikh S.S., Mol,C.D., Slupphaug,G., Bharati,S., Krokan,H.E. and Tainer,J.A. (1998) Base excision repair initiation revealed by crystal structures and binding kinetics of human uracil-DNA glycosylase with DNA. EMBO J., 17, 5214–5226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Waters T.R., Gallinari,P., Jiricny,J. and Swann,P.F. (1999) Human thymine DNA glycosylase binds to apurinic sites in DNA but is displaced by human apurinic endonuclease 1. J. Biol. Chem., 274, 67–74. [DOI] [PubMed] [Google Scholar]
- 55.Yang H., Clendenin,W.M., Wong,D., Demple,B., Slupska,M.M., Chiang,J. and Miller,J.H. (2001) Enhanced activity of adenine-DNA glycosylase (Myh) by apurinic/apyrimidinic endonuclease (Ape1) in mammalian base excision repair of an A/GO mismatch. Nucleic Acids Res., 29, 743–752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hill J.W., Hazra,T.K., Izumi,T. and Mitra,S. (2001) Stimulation of human 8-oxoguanine-DNA glycosylase by AP-endonuclease: potential coordination of the initial steps in base excision repair. Nucleic Acids Res., 29, 430–438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Engelward B.P., Weeda,G., Wyatt,M.D., Broekhof,J.L, de Wit,J., Donker,I., Allan,J.M., Gold,B., Hoeijmakers,J.H. and Samson,L.D. (1997) Base excision repair deficient mice lacking the Aag alkyladenine DNA glycosylase. Proc. Natl Acad. Sci. USA, 94, 13087–13092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hang B., Singer,B., Margison,G.P. and Elder,R.H. (1997) Targeted deletion of alkylpurine-DNA-N-glycosylase in mice eliminates repair of 1,N6-ethenoadenine and hypoxanthine but not of 3,N4-ethenocytosine or 8-oxoguanine. Proc. Natl Acad. Sci. USA, 94, 12869–12874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Roy R., Kennel,S.J. and Mitra,S. (1996) Distinct substrate preference of human and mouse N-methylpurine-DNA glycosylases. Carcinogenesis, 17, 2177–2182. [DOI] [PubMed] [Google Scholar]
- 60.Miao F., Bouziane,M., Dammann,R., Masutani,C., Hanaoka,F., Pfeifer,G. and O’Connor,T.R. (2000) 3-Methyladenine-DNA glycosylase (MPG protein) interacts with human RAD23 proteins. J. Biol. Chem., 275, 28433–28438. [DOI] [PubMed] [Google Scholar]
- 61.Kartalou M., Samson,L.D. and Essigmann,J.M. (2000) Cisplatin adducts inhibit 1,N6-ethenoadenine repair by interacting with the human 3-methyladenine DNA glycosylase. Biochemistry, 39, 8032–8038. [DOI] [PubMed] [Google Scholar]
- 62.Bharati S., Krokan,H.E., Kristiansen,L., Otterlei,M. and Slupphaug,G. (1998) Human mitochondrial uracil-DNA glycosylase preform (UNG1) is processed to two forms one of which is resistant to inhibition by AP sites. Nucleic Acids Res., 26, 4953–4959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Haug T., Skorpen,F., Aas,P.A., Malm,V., Skjelbred,C. and Krokan,H.E. (1998) Regulation of expression of nuclear and mitochondrial forms of human uracil-DNA glycosylase. Nucleic Acids Res., 26, 1449–1457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Grippo P., Orlando,P., Locorondo,G. and Geremia,R. (1982) Uracil-DNA glycosylase in meiotic and post meiotic male germ cells of the mouse. Prog. Clin. Biol. Res., 85, 389–396. [PubMed] [Google Scholar]
- 65.Prasad R., Singhal,R.K., Srivastava,D.K., Molina,J.T., Tomkinson,A.E. and Wilson,S.H. (1996) Specific interaction of DNA polymerase β and DNA ligase I in a multiprotein base excision repair complex from bovine testis. J. Biol. Chem., 271, 16000–16007. [DOI] [PubMed] [Google Scholar]
- 66.Tan Y., Nakagawa,Y., Akiyama,K., Wakabayashi,H., Sarker,A.H. and Seki,S. (1996) cDNA cloning of rat major AP endonuclease (APEX nuclease) and analyses of its mRNA expression in rat tissues. Acta Med. Okayama, 50, 53–60. [DOI] [PubMed] [Google Scholar]
- 67.Wilson T.M., Rivkees,S.A., Deutsch,W.A. and Kelley,M.R. (1996) Differential expression of the apurinic/apyrimidinic endonuclease (APE/ref-1) multifunctional DNA base excision repair gene during fetal development and in adult rat brain and testis. Mutat. Res., 362, 237–248. [DOI] [PubMed] [Google Scholar]
- 68.Duguid J.R., Eble,J.N., Wilson,T.M. and Kelley,M.R. (1995) Differential cellular and subcellular expression of the human multifunctional apurinic/apyrimidinic endonuclease (APE/ref-1) DNA repair enzyme. Cancer Res., 55, 6097–6102. [PubMed] [Google Scholar]
- 69.Kakolyris S., Kaklamanis,L., Giatromanolaki,A., Koukourakis,M., Hickson,I.D., Barzilay,G., Turley,H., Leek,R.D., Kanavaros,P., Georgoulias,V., Gatter,K.C. and Harris,A.L. (1998) Expression and subcellular localization of human AP endonuclease 1 (HAP1/Ref-1) protein: a basis for its role in human disease. Histopathology, 33, 561–569. [DOI] [PubMed] [Google Scholar]