
Figure 4. TASK-3 currents are insensitive to combined depletion of PI(4,5)P2 and PI(4)P by pseudojanin.
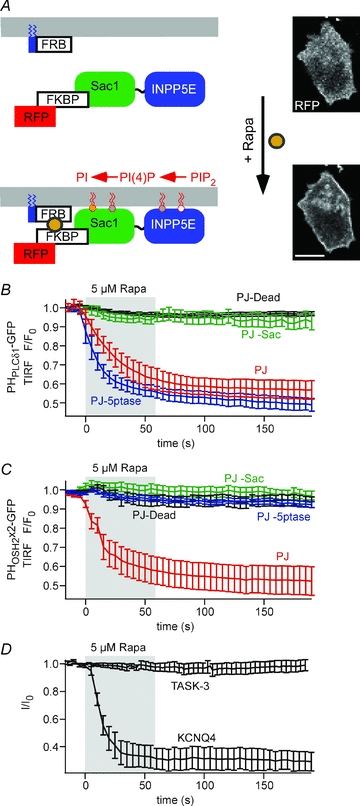
A, schematic representation of the pseudojanin membrane targeting construct (RF–PJ) and its recruitment to the plasma membrane by rapamycin. Confocal images of RF–PJ before (top) and after addition of 5 μm rapamycin (bottom) show efficient translocation of PJ. Scale bar, 10 μm. B, TIRF recordings were performed on cells co-transfected with PHPLCδ1-GFP, Lyn11-FRB, and either of the RF–PJ variants indicated. Dissociation of PHPLCδ1-GFP from the membrane upon application of rapamycin (shaded) was only observed with intact INPP5E 5-phosphatase domain. (n = 9 cells/5 independent experiments, 5/3, 10/4 and 21/8, for PJ, PJ-Sac, PJ-5ptase and PJ-Dead, respectively). C, TIRF recordings from cells co-transfected with PHOSH2×2-GFP, Lyn11-FRB and either of the RF–PJ variants indicated. Dissociation of PHOSH2×2-GFP from the membrane required functional Sac1 and INPP5E phosphatase domains (n = 14 cells /8 independent experiments, 11/5, 15/5 and 14/21, for PJ, PJ-Sac, PJ-5ptase and PJ-Dead, respectively). D, whole-cell voltage-clamp recordings from cells co-transfected with Lyn11-FRB, fully intact PJ and either TASK-3 or KCNQ4. While KCNQ currents were robustly inhibited by recruitment of PJ (residual currents, 30.6 ± 7.8%, n = 6), TASK currents were not affected (residual currents, 97.4 ± 3.9%, n = 7). Voltage protocols as shown in Fig. 1.