

Abstract
The goal of this work is to discuss brain temperature as a physiological parameter, which reflects the balance between metabolism-related intra-brain heat production and heat loss by cerebral circulation to the rest of the body and then to the external environment. First, we present data on fluctuations in brain temperature occurring under physiological and behavioral conditions and discuss their mechanisms. Since most processes governing neural activity are temperature-dependent, we consider how naturally occurring temperature fluctuations could affect neural activity and neural functions. Then, we review brain temperature changes induced by psychomotor stimulants and show that the hyperthermic effects of these drugs are state-dependent and modulated by environmental conditions. Since high temperature could irreversibly damage neural cells and dramatically worsen various pathological processes, we consider the situations associated with pathological brain hyperthermia and evaluate its role in acute perturbations of brain functions, neurotoxicity, and neurodegeneration. We also discuss the complexities and limitations in consideration of brain temperature within the frameworks of physiological regulation and homeostasis. While different adaptive mechanisms could, within some limits, compensate for an altered heat balance of the brain, real life often creates situations when this balance could not be compensated, resulting in pathology and life-threatening health complications.
Keywords: Metabolism, Cerebral blood flow, Brain temperature, Hyperthermia, Hypothermia, Metabolic brain activation, Arousal, Behavior, Psychomotor stimulants, Neural injury, Blood-brain barrier, Neurotoxicity
2. INTRODUCTION
Temperature is one of the basic physical properties of any material object. While it could be intuitively defined as the degree of hotness or coldness, scientific definitions of temperature are quite complex, but it usually defines a quality of physical objects that indicates both whether heat will flow between them and in which direction it will flow. Temperature is closely related to heat, but if temperature is an object’s quality, heat is energy to or from the object because of a temperature gradient.
Although evidence suggests that changes in brain temperature could have profound effects on neural activity and neural functions, our knowledge of brain temperature, its normal and pathological fluctuations, and the mechanisms underlying brain thermal homeostasis is limited. Therefore, the first goal of this work is to consider available data on normal brain temperature, its fluctuations, relationships with body temperature, and mechanisms that determine these fluctuations. Being simultaneously a part of the body and the highest center for regulating physiological functions and behavior, the brain has some degree of autonomy from the rest of the body and differs from the body in regulating its basic parameters (i.e., metabolism, cerebral blood flow, temperature, water and ion content). While the brain plays a crucial role in detecting, via specialized sensory systems, variations in environmental temperatures, integration of this information, and adjusting, via effector mechanisms, heat production and/or heat loss to maintain a stable body temperature (1), it is difficult to apply this logic to brain temperature and it is unclear whether or not it could be viewed as true homeostatic parameter. In contrast to changes in environmental temperatures, which trigger thermoregulatory mechanisms of heat production and heat loss, we will demonstrate that robust changes in brain temperature could occur in a temperature-stable environment following exposure to various salient somatosensory stimuli and during different types of motivated behaviors. While there are different points of view on why these fluctuations occur and what their underlying mechanisms are, we will present evidence that alterations in metabolic brain activity are the primary cause of intra-brain heat accumulation and a force behind more delayed changes in body temperature. Although brain hyperthermia that occurs under specific physiological conditions is associated typically with peripheral vasoconstriction, which limits heat dissipation to the external environment, this change differs from fever, a stable increase in body temperature usually associated with viral and bacterial infections. In the latter case, the brain receives neural and humoral signals from the periphery and actively defends a new, increased temperature baseline by enhancing heat production and/or diminishing heat loss.
The second goal of this study is to consider changes in brain temperature induced by various pharmacological drugs that affect brain metabolism and heat dissipation to the external environment. In contrast to activation of somatosensory pathways which trigger brain activation, thus mediating physiological fluctuations in brain temperature, pharmacological shifts in brain temperature are induced by direct action of drugs on specific centrally and peripherally located neural substrates. Our special focus will be on psychomotor stimulants (i.e., methamphetamine or METH, ecstasy or MDMA), widely abused drugs that induce hyperthermia and could be neurotoxic. We will demonstrate that hyperthermic effects of these drugs are state-dependent and strongly modulated by the environmental conditions that limit heat dissipation, thus resulting in pathological brain hyperthermia.
The final topic of this work is pathological brain hyperthermia. We will consider the situations that could result in pathological changes in the brain’s thermal homeostasis and discuss the mechanisms responsible for this phenomenon. Although high temperature per se is a powerful factor that could directly damage brain cells, it also strongly potentiates negative impacts of other factors and triggers a number of pathophysiological processes that could result in life-threatening health complications and damage of brain cells. Here, we will present our recent data suggesting the role of brain temperature in regulating permeability of the blood-brain barrier (BBB) and the contribution of this mechanism to the development of brain edema and neurotoxicity induced by psychomotor stimulant drugs and other hyperthermic challenges.
3. PHYSIOLOGICAL FLUCTUATIONS IN BRAIN TEMPERATURE: CAUSES AND MECHANISMS
In contrast to abundant information on various sides of brain structure and functions, our knowledge of brain temperature is limited. Surprisingly, the first thermal recordings from the brain of an awake animal were conducted in the late 1860s by German physiologist Moritz Schiff (1870; cited by 2; p. 131), long before the first recordings of the brain’s electrical activity or even the realization that neural cells have electrical activity. Among other important observations, Schiff described a ~1°C brain temperature elevation in a hungry dog after the presentation of meat and showed that this brain response depends on the animal’s motivational state (i.e., hunger). Based on these findings as well as early thermal recordings from the human scalp (Lombard, 1879; Amidon, 1880; cited by 2), William James concluded that “brain-activity seems accompanied by a local disengagement of heat” and speculated that cerebral thermometry could be a valuable tool in experimental psychology, allowing the correlation of brain activity with psychic functions (2; p. 131).
Although this pioneering idea was not reinforced experimentally at the time, further thermorecording work in animals revealed relatively large brain temperature fluctuations (±2–3°C) that occur at stable ambient temperatures following exposure to various environmental challenges, spontaneous changes in activity states, and during different behaviors (3–10). Brain temperature fluctuations reported in these experiments correlated with the biological significance of the environmental challenges, spontaneous and stimuli-induced changes in EEG and motor activity, and had some structural specificity with respect to the modality of sensory stimuli. Although all these findings point toward neural activity as a primary cause of brain temperature fluctuations, temperature changes were generally correlative in different brain structures and associated typically with similar changes in body core temperature. Therefore, brain temperature fluctuations represent physiological reality, which is dependent on metabolic neural activity but is quite different from other traditional measures of neural activity.
Although the brain represents ~2% of the human body’s mass, it accounts for ~20% of the organism’s total oxygen consumption at rest (1, 11). Neurons require several orders of magnitude more energy than do other cells; the power consumption of a single central neuron is about 0.5–4.0 nW, 300–2500 times more than the average body cell (1.6 pW) (12). Biophysical measurements suggest that ~104 ATP molecules are used to transmit a bit of information at a chemical synapse and 106–107 ATP molecules are used for spike coding (13). Although most energy used for neuronal metabolism is spent restoring membrane potentials after electrical discharges (14–17), suggesting a basic relationship between metabolic and electrical neural activity, significant energy is also used on neural processes not directly related to electrical activity, particularly for synthesis of macromolecules and transport of protons across mitochondrial membranes. In addition to neurons, the brain contains metabolically active glial and endothelial cells, which numbers greatly exceed those of neural cells. Since all energy used for brain metabolism appears to be finally transformed into heat (11), intense heat production appears to be an essential feature of brain metabolic activity.
Our initial interest in brain temperature was motivated by the desire to find a simple, sensitive, and integrative index for characterization of alterations in neural activity during motivated behavior. We also knew that psychomotor stimulant drugs induce body hyperthermia, which is believed to be a leading contributor to neurotoxic effects of these drugs. It was unclear, however, whether this effect occurs in the brain, why it occurs, and its relations to body temperature are. Before addressing these issues, we examined brain temperature responses to environmental challenges of different modalities and determined their specifics in different brain structures and other body locations. Since circulation is the primary means of heat exchange between the brain and the rest of the body and the temperature differential between brain tissue and arterial blood inflow determines the direction of heat exchange, it was important to examine the relationships between temperature responses occurring in the brain and in its arterial blood supply.
In these experiments (18), male rats were chronically implanted with miniature copper-constantan thermocouple electrodes in several brain structures (dorsal and ventral striatum, cerebellum) and in the abdominal aorta assessed via the caudal artery. While our goal was to provide accurate measurement of arterial blood temperature that enters the brain and carotid artery seems to be the best measurement point, in rats it is quite difficult to record temperature in this location without full blocking or significantly decreasing blood flow, thus altering the recorded temperature. In contrast, the presence of a miniature catheter with a thermocouple electrode in the abdominal aorta does not result in an evident decrease in blood flow, thus allowing measurements be made in the body core—the area with the high temperature. In contrast to the carotid artery, where the recorded temperatures could be undervalued because of a cooler neck area, temperature values in the abdominal aorta could be only overvalued because of heat inflow from warmer surroundings. Temperature recordings were conducted with high temporal resolution (each 200 ms with subsequent averaging for 2 s) in freely moving rats exposed to a number of physiologically relevant arousing stimuli, ranging from simple sensory to clearly aversive (placement in the cage, 20-s sound, three-min social interaction with either a male or female companion, and three-min tail-pinch). These measurements were made at stable ambient temperatures (23°C).
This study produced several findings. First, each brain structure had its own basal temperature (ventral striatum 37.6°C; dorsal striatum 37.2°C; cerebellum 37.3°C), which was significantly (p<0.001) higher than that of arterial blood (36.6°C). These absolute temperatures typically decreased in each of four recording locations both within each individual session and between sessions, reflecting animal habituation to the recording environment. However, the differences between each brain location and arterial blood always remained positive and relatively similar. Second, all stimuli induced rapid, unique, and relatively long lasting temperature elevations in both brain structures and arterial blood, greatly exceeding the duration of stimulation (Fig. 1). Cage transfer incited the strongest change, social interaction and tail-pinch elicited moderate changes, and sound resulted in the smallest change. Third, temperature changes in each brain structure occurred significantly faster and had stronger amplitude than those in arterial blood (Fig. 1C). This disparity resulted in a significant increase in temperature differentials between brain structures and arterial blood (Fig. 1B). For example, temperature in the dorsal and ventral striatum became significantly higher than baseline from 8 s from the start of a 60-s tail-pinch, while in arterial blood temperature, increase became significant only at 38 s. Fourth, despite a generally correlative time-course, temperature changes had structural specificity. Both striatal divisions showed quite similar changes, but temperature increases in cerebellum were more delayed and prolonged following each stimulus. Finally, brain temperature increases induced by environmental challenges showed changes within repeated tests. While the elevations following tail-pinch, male-male, and male-female interactions remained relatively stable over five repeated daily sessions, responses to sound showed a clear habituation with a complete disappearance of the thermal response by the fifth session. The initial temperature increase associated with environmental change was relatively stable over repeated sessions, but on each subsequent day temperatures decreased more quickly to lower baselines, enhancing the relative response magnitude. Thus, during repeated habituation to the same experimental environment, rats showed a gradual trend of decreasing brain temperatures, which paralleled a decrease in general motor activity.
Figure 1.

Changes in brain (NAcc, dorsal striatum and hippocampus) and arterial blood temperature induced by 3-min tail-pinch in awake, freely moving male rats. A shows mean (±sem) values of absolute temperatures (°C); B shows differentials between relative temperature changes in each brain structure and in arterial blood; C shows rapid-time course resolution of temperature changes (2 s). Filled symbols in each graph indicate values significantly different from baseline. Vertical hatched lines show onset and offset of tail-pinch. For additional details see 18.
Given that the blood supply to the brain is cooler than the brain itself, and that brain temperatures rise more quickly and to a larger extent than do arterial blood and head muscle temperatures in response to all challenges, intra-brain heat production appears to be the primary cause of functional brain hyperthermia. While arterial blood temperature also gradually increases in response to all challenges, brain-arterial blood temperature differentials grow consistently during behavioral activation, showing an apparent increase in intra-brain heat production. Therefore, it seems that increased blood circulation that accompanies functional brain activation removes heat from the brain. Tail-pinch similar to that used in our study induced an almost two-fold increase in striatal blood flow; as with brain temperature, this increase was rapid and greatly exceeded the duration of stimulation (19). Rapid prolonged increases in striatal blood flow (80–120%) also occurred during grooming and eating (19), activities consistently associated with brain temperature increases (3, 4, 20). Hence, brain circulation is a significant factor in the re-distribution of locally released heat within brain tissue and in its removal from the brain, thus contributing to brain temperature fluctuations occurring under behavioral conditions.
Our subsequent studies (21, 22) revealed that increases in brain temperature induced by various arousing stimuli are accompanied by slightly delayed and weaker temperature increases in musculus temporalis, a non-locomotor head muscle that receives the same arterial blood supply as the brain from the carotid artery, as well as a biphasic temperature fluctuation in the skin (Fig. 2A). These between-site differences were evident immediately after the start of stimulation, resulting in significant increase in brain-muscle temperature differential (see Fig. 2B). This accelerated rise of brain vs. muscle temperatures (analyzed as a relative change) appears to be a true index of brain activation, correlating more tightly with locomotor activation and acute skin hypothermia. The latter response is an obvious consequence of peripheral vasoconstriction—a known adaptive organism’s response that is triggered by various stressful stimuli (23–25) and aimed at heat retention under conditions of potential danger. While changes in brain and muscle temperatures occurred with some latency, skin temperature response was quite rapid, typically being seen during the first 10–20 s after the start of stimulation, and correlating with the locomotor response (C). However, skin hypothermia was transient and inverted in rebound-like skin warming, suggesting enhanced heat dissipation. These changes in heat retention and loss are evident in skin-muscle differentials, which strongly decreased for 10–20 min after arousing stimuli but later increased above baseline.
Figure 2.

Changes in brain (NAcc), muscle (musculus temporalis) and skin temperatures induced in male rats by tail-pinch (left panel) and social interaction with female (right panel). A shows relative temperature changes; B shows NAcc-muscle and skin-muscle temperature differentials, and C shows locomotion. Vertical hatched lines in each graph show onset of stimulation and filled symbols denote values significantly different from pre-stimulus baseline. For additional details see 21.
Brain temperature responses induced by various arousing stimuli depend upon basal brain temperature. This correlation was found with all tested stimuli, as shown in Fig. 3 for the procedures of subcutaneous and intraperitoneal saline injections, tail-pinch, male-male social interaction, and presentation of sexually arousing stimuli originated from the receptive but inaccessible female partner to sexually experienced male rats. In each case, temperature increase was large when basal temperatures were low, and the response became gradually weaker when basal temperatures were high. In each case, regression lines crossed the line of no effect within 38–39°C, suggesting that the response should fully disappear at high temperatures. Such high temperatures, however, are well within the range of physiological fluctuations and they occurred, for example, during the rats’ relocation in novel environmental conditions (26) or natural copulatory behavior (22). This correlation appears to be valid for any arousing stimuli, reflecting some basic relationships between basal activity state (basal arousal) and its changes induced by environmental stimuli. This correlation, moreover, is weaker for stimuli that induce smaller temperature increases and stronger for powerful stimuli that result in larger temperature fluctuations. These observations may be viewed as examples of the 'law of initial values', which postulates that the magnitude and even direction of autonomic response to an «activating» stimulus is related to the pre-stimulus, basal values (27, 28). This relationship was evident for a number of homeostatic parameters, including arterial blood pressure, body temperature, and blood sugar levels.
Figure 3.
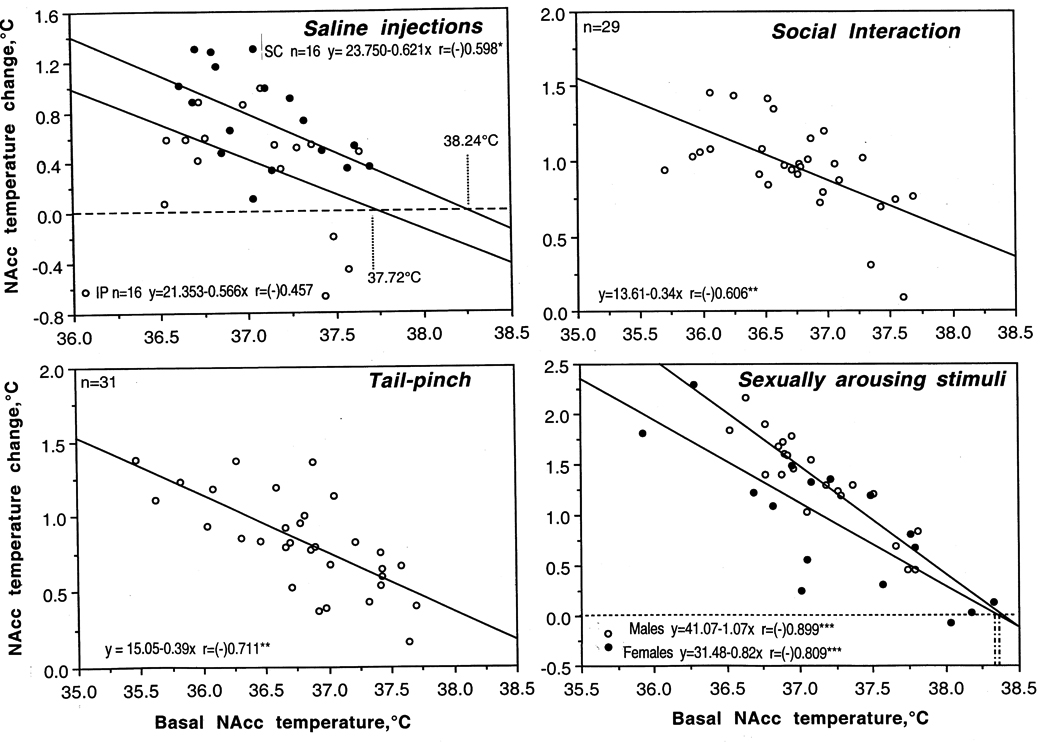
Relationships between changes in brain temperature induced in rats by various arousing stimuli [intraperitoneal (ip) and subcutaneous (sc) injections of saline (0.2 ml), social interaction with female rat, 3-min tail-pinch and exposure of sexually-experienced males to females] and basal brain temperature. Each graph shows dependence of NAcc temperature change induced by a stimulus from pre-stimulus basal NAcc temperatures. In each case, temperature elevation was negatively correlated with basal temperature (each graph shows regression lines, regression equations and coefficients of correlation, r). Original data were previously published (21, 90).
Although different brain structures show a generally similar pattern of temperature fluctuations, their basal temperatures differ significantly. Tested at the same conditions, more dorsally located structures were consistently cooler (dorsal striatum, 36.6°C; hippocampus, 35.6°C) than more ventrally located structures (ventral tegmental area of midbrain, 37.3°C; medial preoptic area of hypothalamus, 37.3°C; nucleus accumbens or NAcc, 37.3°C). Importantly, these between-structural differences exist in various environmental conditions despite large fluctuations in absolute temperatures. While these data support a dorso-ventral temperature gradient described in animal (4, 9, 29) and human brains (30–32), the reasons and underlying mechanisms for this gradient remain unclear. One point of view suggests that higher temperature in more ventrally located structures reflects their further location from the colder environment and more heating by, presumably, warm blood from the body (33–35). However, this assumption, inferred from rectal or core body measurements, is challenged by direct measurements of arterial blood temperatures (8, 18, 36), which suggest that in awake animals, the brain is always warmer than the arterial blood supply and thus cannot be warmed by arterial blood. While distance from the external environment could not be excluded as a factor, the impact of this factor seems low in light of the brain’s heavy insulation and intense blood flow.
Another point of view relates dorso-ventral temperature gradient to differences in metabolic activity, particularly in structure-specific expression of brain uncoupling proteins that regulate uncoupling in mitochondria and local heat production (29) and in their electrical activity. In the ‘cold’ cortex, for example, a great majority of neural cells (>95–98%) are silent at rest, but show phasic excitations as a result of either sensory stimulation or local glutamate application. The same is true for dorsal striatal cells, most of which (at least 90–95%) in freely moving rats maintain electric silence at rest but show phasic stimuli- or glutamate-induced and movement-related excitations (37). In contrast, most neurons of the ‘warm’ hypothalamus and VTA are spontaneously active at rest (38–40, 39). Dorso-ventral differences in neuronal impulse activity also have been reported in our previous work both within the diencephalon (38) and forebrain (41). Finally, dorso-ventral temperature differences, especially in small animals, may be due partially to technical aspects of thermorecording. When the tip of a temperature-sensitive metal probe is not in tight contact with brain tissue or is implanted superficially, heat dissipation from the probe to the external environment may be greater and measured temperatures will be undervalued. Even greater mistakes will occur if temperatures are measured from an open skull.
Numerous data suggest that increased brain metabolism is accompanied by increased brain circulation, and changes in local cerebral blood flow are widely used as a measure of functional brain activation. However, the relationships between brain metabolism and cerebral blood flow are complex and are currently poorly understood (see 42, 43 for review). While temperature is usually omitted from equations relating metabolism and blood flow or viewed as a passive parameter (44, 45), direct relationships between temperature and blood flow are well established in peripheral tissues. Strong linear increases in blood flow during local temperature increases were shown in skin (46, 47), muscular tissue (48), the intestine (49), and the liver (50). The same relationships were also observed in the brain of monkeys (51), rats (52), and humans (53). Increases in local brain temperature resulting from enhanced neural metabolism can therefore be understood as one of the factors that increase local blood flow. This factor could explain, at least partially, the well-known but little-understood phenomenon of a blood flow increase that exceeds metabolic activity of brain tissue (42). Since metabolic activation is accompanied by heat production and increased local temperature, a greater blood flow response could reflect not only an increased metabolism but also an increased brain temperature. Using this mechanism, the brain is able to increase blood flow more and in advance of actual metabolic demands, thus providing a crucial advantage for successful goal-directed behavior and the organism’s adaptation to potential energy demands. By increasing blood flow above the brain’s current demand, more oxygen and nutrients are delivered to the areas of potential demand and more potentially dangerous metabolic heat is removed from intensively working brain tissue. While special experiments with parallel measurements of temperature and blood flow (which is a quite complex task, especially in small animals) are necessary to clarify this issue, it appears that the brain could “regulate” its own temperature by temperature-dependent changing of its blood flow.
4. ALTERATIONS IN BRAIN TEMPERATURE HOMEOSTASIS DUE TO INSUFFICIENCIES OF HEAT DISSIPATION
Although heat cannot be delivered to the brain from the periphery, it could be accumulated in the brain when it cannot be properly dissipated to the external environment. In this case, brain hyperthermia results from temperature rise in the body core and arterial blood temperature, decreasing heat dissipation from brain tissue. Such hyperthermia usually occurs from a combination of excessive heat production coupled with insufficient heat loss. It is known that intense physical activity is associated with robust energy consumption and significant heat production. In humans, oxygen consumption increases up to ten-fold in the transition between quiet resting conditions and intense running (1, 54). This increase corresponds to whole-body metabolic heat production, which increases from ~ 1 W/kg at rest to ~10 W/kg during heavy exercise (55). Enhanced heat production is compensated by enhanced heat loss, which in humans depends on a well-developed ability to sweat and on a dynamic range of blood flow rates in the skin, which are much higher than those in other species. Skin blood flow in humans, for example, can increase from ~0.2–0.5 l/min in thermally neutral conditions to 7–8 l/min under maximally tolerable heat stress (56). Sweat rates under these conditions may reach up to 2.0 l/h, providing a potential evaporative rate of heat loss in excess of 1 kW (or ~14 W/kg), i.e., more than maximally possible heat production. These compensatory mechanisms, however, become progressively less effective in hot, humid conditions, resulting in progressive heat accumulation in the organism. For example, body temperatures measured at the end of a marathon run on a warm day were found to be as high as 40°C (57) and cases of fatigue during marathon running were associated with even higher temperatures (58). While 90-min of intense cycling in experienced cyclists at normal environmental temperatures increased body temperature less than 1°C, 2.0–2.5°C increases were found when participants cycled in water-impermeable suits that restricted heat loss via skin surfaces (53).
Although body hyperthermia associated with intense physical activity under conditions that restrict heat dissipation is a known phenomenon, the impact of these conditions on the brain is a matter of speculation and controversy (59–62). Since the direct recording of brain temperature in humans is usually impossible, tympanic temperature has been used as an indirect measure of brain temperature (63, 64). Using this approach during exercise with and without active cooling of the head, it was assumed that brain temperatures during intense body heat production remain lower than body temperatures, suggesting selective brain cooling as a mechanism preventing brain over-heating during extreme body hyperthermia. Direct measurements of brain temperature in neurological patients (31, 65, 66, 67) and applications of more sophisticated physiological recordings (53, 68), however, called the existence of a brain cooling mechanism in humans into question. By directly measuring temperatures in the carotid artery and internal jugular vein in human volunteers, it was shown that at rest, venous blood exiting the brain is ~0.3°C warmer than arterial blood entering the brain. Although during 10-min cycling both temperatures increased about 1°C and the increase was stronger in the carotid artery than in the jugular vein, the venous-arterial difference always remained positive. When the cycling was done in water-impermeable suits, the increases were more dramatic (up to 39.3 and 39.5°C for arterial and venous blood, respectively), but the difference still remained positive. After termination of exercise, arterial blood temperature dropped rapidly, while venous temperature decreased more slowly. Simultaneous measurements of tympanic temperatures under these conditions revealed that their values are consistently lower than, and independent of, arterial and venous temperatures. Therefore, human brain temperature may also increase during intense physical exercise, reaching relatively high levels under conditions that restrict heat dissipation to the external environment. Although arterial blood always remains cooler than brain tissue and removes metabolic heat from the brain, progressive intra-brain heat accumulation occurring during intense physical activity reflects a failure of this natural cooling mechanism due to an inability to dissipate metabolic heat to the external environment and gradual warming of arterial blood arriving to the brain.
While diminished heat outflow from the brain appears to be the primary cause of intra-brain heat accumulation, physical exercise is also associated with increased brain metabolism (69, 70) and consequently enhanced intra-brain thermogenesis. Taking into account uptake of oxygen, glucose, and lactate, an almost two-fold increase in global brain metabolism was reported during 10-min intense cycling at normal environmental temperatures. Although cerebral blood flow increases during physical exercise and this increase is in excess of enhanced cerebral metabolic activity (53, 68), cerebral blood flow decreases gradually during maximal exercise at hyperthermic conditions because of a hyperventilation-induced decrease in CO2 pressure (68). In contrast to more intense heat removal from the brain by blood flow during physical exercise under normal conditions, under hyperthermic conditions compromised cerebral blood outflow is an additional, powerful factor that diminishes heat dissipation from the brain, thus promoting intra-brain heat accumulation.
Although robust brain hyperthermia (~39.5°C) did occur on human volunteers during intense cycling under conditions that restrict heat dissipation (53), this activity did not result in clear fatigue. Hypothalamic temperatures associated with forced exercise-induced fatigue in rats were higher (40.1–42.1°C; 71) than those (40.0–40.7°C for body and deep muscles) found in humans during self-motivated exercise (72). These temperatures exceeded the upper limits of physiological increases (~39.5–40.0° for deep brain structures; see section 3) and were close to the threshold values associated with disturbances in permeability of the brain-blood barrier, development of brain edema, and irreversible damage of neural cells (see section 5). Although fatigue during intense exercise may protect the brain against further overheating, robust brain hyperthermia associated with this condition is an obvious contributor to serious acute and long-term health complications reported in sportsmen and military recruits engaged in extreme physical activity under harsh environmental conditions.
Although temperatures of venous blood exiting the brain may be viewed as the best indirect measure of brain temperature in humans, direct measurements in the brain, jugular bulb, and body core (rectum) revealed unexpected differences between these areas. First, temperatures in the cortex were found to be 0.2–0.8°C higher than in the body core (31, 65, 66, 67). Second, cortical temperatures were ~1.0°C higher than those in the jugular vein, which were similar to core body temperature (31, 65). Third, jugular temperatures were found to correlate more strongly with rectal than brain temperature (65). Brain temperatures in humans remained higher than body temperature not only during normothermic conditions but also during fever, suggesting that the human brain has no specific protection against thermal impact (65, 67). Although these data were obtained in neurological patients, they suggest that the use of jugular vein temperatures as an index of brain temperature may in fact underestimate real brain temperature. This underestimation may be related to the procedural features of measurement. Because venous blood flow is slow and the catheter with the thermosensor is in tight contact with the vessel’s walls, the measured temperatures are influenced by cooler neighboring tissues and thus are lower than true brain temperature. If jugular venous measurements underestimate brain temperature, the real temperature difference between brain and arterial blood under resting conditions may be greater than 0.3°C (53), as suggested by our measurements in rats (18).
5. PATHOLOGICAL BRAIN HYPERTHERMIA INDUCED BY PSYCHOMOTOR STIMULANTS: CAUSES AND MECHANISMS
Most psychoactive drugs have direct effects on the brain and organism metabolism. For example, amphetamine-like psychomotor stimulant drugs (i.e., amphetamine, methamphetamine or METH, MDMA or Ecstasy) induce behavioral and sympathetic activation, increase brain metabolism (73, 74) and elevate temperatures (75–79). These drugs also have peripheral vasoconstrictive effects (80, 81), thus diminishing heat dissipation to the external environment. Finally, these drugs are widely abused and often taken under conditions of high psychophysiological activation and in warm, humid environments, potentiating drug-induced metabolic activation and complicating proper heat dissipation. This combination of drug effects, the activity state, and environmental conditions could result in pathological hyperthermia, which appears to be tightly related to dangerous, and even fatal, acute decompensation of vital functions, and neurotoxicity (82–85), a clinically important complication of chronic use of these addictive drugs.
To examine how these drugs affect brain temperature and how their effects are modulated by environmental conditions that mimic human use, we examined temperature changes in two brain structures (NAcc, hippocampus) and temporal muscle induced by METH (1–9 mg/kg, sc) in male rats in quiet resting conditions at normal laboratory temperatures (23°C), during social interaction with a female, and at moderately warm ambient temperatures (29°C) (86–88).
Figure 4 shows the effects of METH at the same high dose (9 mg/kg, sc) in these three conditions. As can be seen, METH strongly increased brain (NAcc and hippocampus) and temporal muscle temperatures during quiet resting conditions at normal ambient temperatures (left column; A). The increase was stronger in both brain sites than in the muscle, exceeding that following natural arousing stimuli (B; compare with Fig. 1). A stronger temperature increase in brain vs. muscle increased brain-muscle differentials (C); this effect was also more profound and prolonged than that seen with natural stimuli. Therefore, intra-brain heat production associated with metabolic brain activation appears to be the primary cause of METH-induced brain hyperthermia and a factor behind more delayed and weaker body hyperthermia. This temperature increase was about 3.5–4.0°C above normal baseline (mean ~40°C) and could be viewed as a pathological hyperthermia. Although some animals showed even stronger temperature increases (up to 41.0°C), they always survived this drug exposure.
Figure 4.
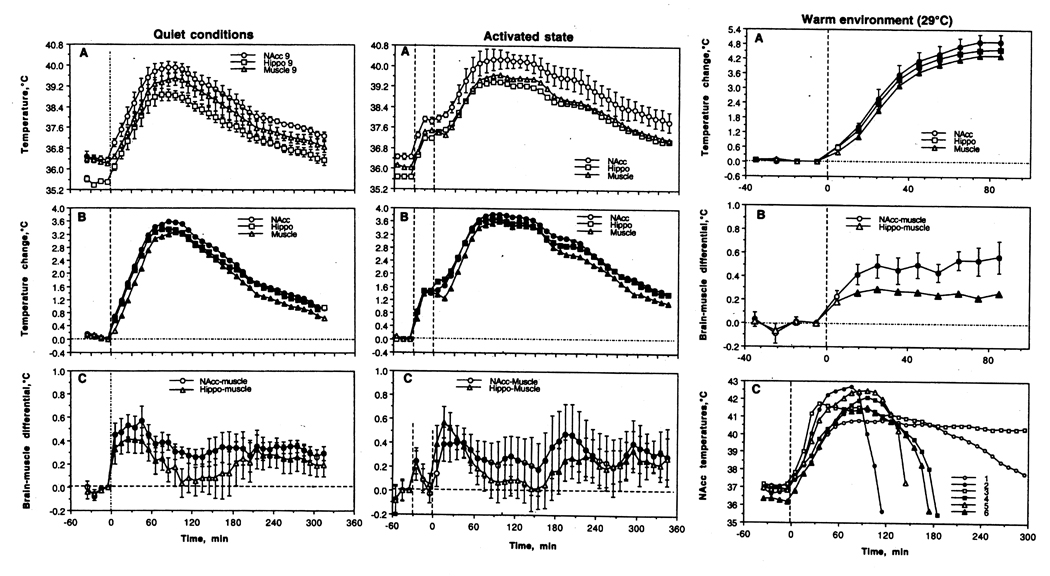
Changes in brain (nucleus accumbens or NAcc and hippocampus or Hippo) and temporal muscle temperatures induced in male rats by methamphetamine (9 mg/kg) administered under different environmental conditions. Left panel shows the effects in quiet resting conditions at normal ambient temperatures (23°C), middle panel shows the effects under conditions of physiological activation (interaction with female started 30-min before drug injection; shown as first hatched line), and right panel shows the effects in warm environmental conditions (29°C). The moment of drug injection is shown by vertical hatched line at 0 min. Graphs in left and central panels represent absolute (A) and relative temperature (B) changes as well as brain-muscle differentials (C). Filled symbols mark values significantly different from baseline. Since most rats exposed to METH at 29°C died during the experiment, mean temperature data for this group (right panel) are shown only for the period when all rats were alive (A, relative temperature change; B, brain-muscle temperature. C in this panel shows temperature dynamics in all experimental rats hours 5 hours following drug injection. Four rats (2,4,5 and 6) that showed robust hyperthermia (41–42.5°C) died between 2 and 3 hours post-injection. Another rat (1) that survived the period of recording died over-night. For other details see 87, 88.
Although METH-induced temperature increases were larger in brain locations vs. temporal muscle (~0.4°C), this brain hyperthermia is a part of general hyperthermia, which is evident in different body locations. While skin temperature transiently decreases after METH administration, reflecting drug-induced peripheral vasoconstriction, it later increases well above baseline because of a progressive warming of arterial blood inflow.
When the same drug was injected 30-min after the start of social interaction with a female rat and pre-injection baselines were ~1°C higher, the hyperthermic effect of METH was significantly larger and more prolonged than that seen in quiet resting conditions (Fig. 4A and B, middle column). Importantly, METH administered under activated conditions induced stronger and more prolonged increases in brain-muscle differentials than those seen in controls (C). This finding once again suggests a role for metabolic brain activation and intra-brain heat production in the genesis of hyperthermia. While the potentiating effect of social interaction was not additive, mean brain temperatures in this case maintained above 40°C for more than two hours. Finally, the effects of METH were greatly potentiated by a slight increase in ambient temperature (29°C; Fig. 4, right column). This ambient temperature is close to normothermy in rats (89) and, as we showed before (90), did not affect animal behavior and basal temperatures. When administered at 29°C, METH increased temperatures rapidly in all animals, resulting in most animals in clearly pathological hyperthermia (>41–42°C) and death in five of six animals (C). Similar data were also obtained with MDMA (ecstasy), another widely abused amphetamine-like psychostimulant drug with psychedelic properties (88). In contrast to METH, MDMA at low doses slightly decreased brain and body temperatures and animal activity in standard laboratory conditions and its hyperthermic and stimulant effects at equal doses were weaker than those for METH. However, the hyperthermic effects of this drug were potentiated by social interaction even stronger, possibly due to its stronger and more prolonged vasoconstrictive effects (80, 81). Similar to METH, MDMA at a dose (9 mg/kg) about ~1/6 of LD50 (91) induced pathological hyperthermia and lethality in most rats when used at moderately warm ambient temperatures (29°C). Therefore, one (1.5 mg/kg) or two tablets of MDMA may be highly toxic in predisposed individuals if consumed in adverse environmental conditions. This powerful modulation of drug-induced toxicity by relatively weak changes in environmental conditions may explain exceptionally strong, sometimes fatal, responses of some individuals to amphetamine-like substances seen under conditions of raves (84).
6. BRAIN TEMPERATURE AS A FACTOR AFFECTING NEURAL ACTIVITY AND NEURAL FUNCTIONS, BBB PERMEABILITY, AND MORPHOLOGY OF BRAIN CELLS
In establishing the role brain temperature plays in normal physiological functions and the development of pathological processes, it is important to realize that it could be viewed from two different perspectives. From one perspective, changes in brain temperature could be viewed as an index of altered thermal homeostasis, which is determined by metabolic brain activity and heat loss via cerebral blood flow. Therefore, brain temperature fluctuations depend on alterations in neural activity, brain metabolism, and modulated by thermal influences from the body via alterations of cerebral blood flow. This perspective makes brain temperature an important and relatively easily recorded physiological parameter, which could provide valuable information on basic metabolic brain mechanisms involved in different behaviors. Although these mechanisms could be viewed as non-specific, it does not mean that they are not important or that they should be ignored. While these issues are not within the goals of this review article, brain thermorecording has been used previously to study sexual behavior (22, 92), feeding behavior (20) as well as heroin (93) and cocaine self-administration behavior (94).
From another perspective, temperature could be viewed as a factor that directly affects brain cells, neural activity, cerebral blood flow, permeability of the brain-blood barrier (BBB), and brain water and ion content, thus strongly modulating neural functions. Literature data and our own work to support this role will be discussed below.
6.1. Temperature modulation of neural activity
Although heat is an obvious “by-product” of metabolic activation and is re-distributed continuously within the brain via conduction and blood flow, brain temperature changes that are determined by heat production and loss may play important adaptive and integrative roles, involving and uniting numerous central neurons within the brain. Although this type of inter-neuronal communication is obviously not the main, nor the most efficient, it may have adaptive significance since most physical and chemical processes governing neural activity are temperature-dependent. While it is generally believed that most physical and chemical processes are affected by temperature with the average Van’t Hoff coefficient Q10=2.3 (i.e., doubling with 10°C change [95]), experimental evaluations using in vitro slices revealed widely varying effects of temperature on passive membrane properties, single spike and spike bursts as well as the neuronal responses (i.e., EPSP and IPSP) induced by electric stimulation of tissue or its afferents (96–99). While confirming that synaptic transmission is more temperature-dependent than the generation of action potentials (100), these studies showed that temperature dependence varies greatly for each parameter, the type of cells under study, and the nature of afferent input involved in mediating neuronal responses.
Although temperature-sensitive neurons were first described in the preoptic/anterior hypothalamus (101–103) and they were originally viewed as primary central temperature sensors, cells in many other structures (i.e., visual, motor and somatosensory cortex, hippocampus, medullar brain stem, thalamus) also show dramatic modulations of impulse activity by temperature. Many of these cells have a Q10 similar to classic warm-sensitive hypothalamic neurons. In the medial thalamus, for example, 22% of cells show a positive thermal coefficient >0.8 imp/s/°C (104), exceeding the number of temperature-modulated cells found in this study in both anterior (8%) and posterior (11.5%) hypothalamus. About 18% of neurons in the superchiasmatic nucleus are warm-sensitive (105), while >70% of these cells decrease their activity rate with cooling below physiological baseline (37–25°C) (106). Finally, high temperature sensitivity was found on electrophysiologically identified substantia nigra dopamine neurons in vitro (107). Within the physiological range (34–39°C), their discharge rate increases with warming (Q10=3.7) and dramatically decreases (Q10=8.5) during cooling below physiological range (34–29°C).
Temperature also strongly modulates release of various neuroactive substances. For example, Q10 for K+-induced glutamate release in vitro was 3.6–5.5, 3.5–6.3 for GABA release, and within 11.3–37.7 for K+- and capsaicin-induced release of calcitonin gene-related peptide (108, 109). However, in vivo, these dramatic changes in release are compensated by increased transmitter uptake. For example, within the physiological range, dopamine uptake almost doubles with a 3°C temperature increase (Q10=3.5–5.9; 110), a fluctuation easily achieved in the brain under conditions of physiological activation.
The fact that temperature has strong effects on various neural parameters, ranging from the activity of single ionic channels to such integrative processes as transmitter release and uptake has important implications. First, it suggests that naturally occurring fluctuations in brain temperature affect various parameters of neural activity and neural functions. While in vitro experiments permit individual cells to be studied and individual components of neural activity and synaptic transmission to be separated, neural cells in vivo are interrelated and interdependent. Therefore, their integral changes may be different from those of individual components assessed in vitro. For example, increased transmitter uptake should compensate for temperature-dependent increase in transmitter release, thus limiting fluctuations in synaptic transmission. By increasing both release and uptake, however, brain hyperthermia makes neurotransmission more efficient and neural functions more effective at reaching behavioral goals. Therefore, changes in temperature may play an important integrative role, involving and uniting numerous central neurons within the brain.
6.2. Temperature modulation of BBB permeability and brain water and ionic homeostasis
Along with the influences of temperature fluctuations on neural activity, strong temperature increases or decreases could adversely affect brain cells and brain functions. While brain cells seems to tolerate well low temperatures (111, 112), multiple in vitro studies suggest that high temperature (>40.0°C) has destructive effects on various cells (113, 114), especially profound in metabolically active brain cells (115–119), including neuronal, glial, endothelial, and epithelial cells (120, 121). Rapid damage to brain cells has been also documented in vivo during extreme environmental warming (122–124) and acute METH intoxication (125, 126), which both result in robust brain hyperthermia (see above) as well as increased permeability of the blood-brain barrier (BBB) and vasogenic edema (122, 124, 125, 127). The integrity of the BBB is also compromised during opiate withdrawal (128), intense physical exercise in a warm environment (129), and during restraint and forced swim stress (130–132)—conditions associated with brain hyperthermia (53, 133). Although all these data implicate brain hyperthermia as a leading factor in BBB leakage and subsequent damage to brain cells, these changes may be also affected by many other factors (i.e., metabolic brain activation, oxidative stress, alterations in cerebral blood flow, hypoxia) (134–136), which contribute to alterations in BBB permeability and subsequent structural brain damage (137–139).
To clarify the role of brain temperature in modulating BBB permeability, it is essential to delineate this physical factor from other possible contributors. To reach this goal, we examined several brain parameters in pentobarbital-anesthetized rats, whose bodies were passively warmed to produce different levels of brain temperature, which was monitored by chronic brain thermocouple probes. As shown in our previous study (140), sodium pentobarbital administered at a typical anesthetic dose (50 mg/kg, ip) under standard laboratory conditions (23°C) induces robust brain and body hypothermia (~31–33°C) associated with relative skin warming, suggesting loss of vascular tone and increased heat loss to the environment. In addition to metabolic inhibition that is a primary cause for brain and body hypothermia, anesthetized rats became very sensitive to environmental temperatures, becoming hypothermic at low ambient temperatures and hyperthermic when their bodies are warmed. Therefore, by changing the intensity of body warming, we were able to produce a wide range of brain temperatures from very low, hypothermic (>32°C) to very high, hyperthermic (<42°C). This paradigm was used to evaluate the role of brain temperature in regulating BBB permeability and alterations in tissue water and ions as well as the development of acute morphological abnormalities of brain cells (Kiyatkin and Sharma, unpublished observations). The brains were taken at the same time after the start of anesthesia (90 min) and the initiation of body warming, but at different levels of brain temperature and analyzed for several brain parameters. Similar to other studies, NAcc here was chosen as a representative deep brain structure. To evaluate the integrity of BBB we used immunohistochemistry for endogenous albumin, a relatively large plasma protein (molecular weight 59 kDa, molecular diameter 70 nm) that is normally confined to the luminal side of the endothelial cells and is not present in the brain. Thus, the appearance of albumin-positive cells and albumin immunoreactivity in the neuropil indicates a breakdown of the BBB. Brains were also evaluated for water and ion (Na+, K+, Cl−) content and for the presence of morphologically abnormal cells, using Nissl staining.
As shown in Fig. 5A (left panel), the number of albumin-positive cells is strongly dependent on brain temperature, being minimal at normothermic values (34.2–38.0°C), slightly higher (2–4-fold) at hypothermic values (34.2–32.2°C), and dramatically higher (~26-fold) at hyperthermic values (38.0–42.5°C). The increase was evident from ~39°C, progressed at higher temperatures, and plateaued at high levels at 41–42°C. Temperature-dependence of albumin immunoreactivity was evident in each tested brain structure, with some structural differences (Fig. 5B). There were also some differences in albumin immunoreactivity in different cortical areas, with much stronger changes in the piriform cortex (Fig. 5C).
Figure 5.

Temperature dependence of albumin imminoreactivity and cellular brain abnormalities. Data (the cell counts within the same areas within well-defined anatomical structures) are shown for the brain as a whole (A; open circle show original data and closed circle show mean averages), individual brain structures (B), and individual cortical areas (C). For details see the text.
It is well established that albumin entry from the peripheral circulation to the brain results in increased tissue water content associated with robust alterations in ionic brain balance (141, 142). These alterations result in vasogenic edema—a dangerous and often fatal complication of various pathological processes in the brain as well as conditions associated with brain hyperthermia (i.e., heat stress, opiate withdrawal, and METH and MDMA intoxication). As shown in this study, tissue water content (evaluated in the cortex and thalamus) was also strongly dependent on NAcc temperature (Fig. 6). Cortical water content was lowest during hypothermia, significantly higher during normothermia, and maximal during hyperthermia. Within the range of recorded temperatures, cortical water differed within ~4%. In the thalamus, water content was clearly higher during hyperthermia, but values at low and normal temperatures were virtually identical. Cortical water content during anesthesia in normothermic conditions was virtually identical to that in awake control conditions (see hatched lines), but significantly higher (edema) in hyperthermia and significantly lower (dehydration) in hypothermia. A similar trend was seen in the thalamus, where water content during anesthesia was lower than in awake controls at low and moderate brain temperatures and similar to control at high temperatures. Moreover, the numbers of albumin-positive cells and tissue water were closely interrelated both in the thalamus and cortex (r=0.96 and 0.89, respectively; see Fig. 7A). This correlation was highly linear in the thalamus, but had some divergence in the cortex at values that correspond to extreme hypothermia (see a circle in Fig. 7A). Despite the presence of few albumin-positive cells (see also Fig. 5.B), cortical water was relatively lower, suggesting that the tight correlation between brain albumin and water, which exists within the entire range of normal and high temperatures, could be distorted at very low temperatures. Therefore, it appears that extreme hypothermia also results in BBB leakage, but “hypothermic” brains appear to be dehydrated compared to “normothermic” brains taken from awake and anesthetized animals. It is unclear, however, whether these disturbances in water homeostasis are related to hypothermia per se or to pentobarbital’s inhibiting action on metabolism (143, 144). Importantly, brain temperatures never drop below 34–35°C (in deep structures) in any physiological conditions and such extreme hypothermia could only be seen during general anesthesia, near-lethal environmental cooling, and overdose with powerful sedative drugs.
Figure 6.

Temperature dependence of tissue water shown separately for cortex and thalamus. Horizontal hatched lines show “normal” values evaluated in brains of awake, drug-free rats at normothermia. Vertical hatched lines show limits of “normal” brain temperature. In both structure, tissue water content was directly and strongly dependent on brain temperature (regression lines, regression equations, and coefficients of correlation are shown for the cortex). For details see the text.
Figure 7.

Correlative relationships between individual brain parameters assessed in urethane-anesthetized rats passively warmed to different brain temperatures. A shows the relationships between numbers of albumin-positive cells and tissue water evaluated separately in the cortex and thalamus. B shows the relationships between the numbers of albumin-positive and morphologically abnormal cells evaluated in the brain as a whole. C shows the relationships between tissue water content and amounts of morphologically abnormal cells evaluated separately in the cortex and thalamus. Each graph contains coefficients of correlation and regression lines. Hatched line in B shows a line of equality. Each correlation coefficient value is highly significant (p<0.001). For additional details see the text.
These data indicate that brain hyperthermia, independently of its cause, could be a strong factor for eliciting breakdown of the BBB. Importantly, albumin-positive cells appeared in the brain relatively quickly (20–80 min) and within the range of physiological hyperthermia (38.5–39.5°C), suggesting that increased BBB permeability is not solely pathological but also a normal physiological phenomenon occurring during various conditions associated with hyperthermia. Such hyperthermia, for example, occurs during copulatory behavior and heroin self-administration (133). Although temperature dependence of BBB permeability was evident in all tested brain structures, suggesting its generalized nature, there were also minor but significant differences among individual brain areas.
6.3. Temperature effects on morphology of brain cells: role of BBB leakage and edema
Strong temperature dependence has been also found with respect to the number of morphologically abnormal cells (Fig. 5, right panel). Structural abnormalities were absent at low and normal temperatures but their numbers linearly increased during hyperthermia. This pattern was evident in each of four tested structures (Fig. 5B), although some areas (i.e., thalamus) or cortical sub-areas (i.e. piriform cortex), which showed robust BBB leakage, also showed more profound structural cell abnormalities (Fig. 5B and C). The numbers of morphologically abnormal cells were closely related to counts of albumin-immunoreactive cells (Fig. 7B). Abnormal cells were absent when there were no albumin-positive cells and both parameters increased proportionally.
This study confirms multiple in vitro observations, suggesting that brain cells are exceptionally sensitive to thermal damage and demonstrates that the number of structurally abnormal cells directly and strongly depends on brain temperature. A few abnormal cells were found at ~38.5°C, and their numbers gradually increased as temperature rose. While the counts of albumin-positive cells plateaued at high temperatures, morphological abnormalities linearly increased and peaked at the maximal detected temperature (42.4°C). Similar to other parameters, structural abnormalities occurred relatively quickly and were closely related to BBB leakage and increased tissue water content (see Fig. 7B and C). Therefore, even with passive warming, morphological damage reflects not only the effect of temperature per se, but also BBB leakage and associated edema.
Although virtually no abnormal cells were seen in the brain in normothermic or hypothermic conditions (see Fig. 5A and B), a small but significant effect was found in the hypothalamus and especially in the piriform cortex during extreme hypothermia. In contrast to robust degeneration of cell nucleus and pyknosis with clearly edematous neuropil during hyperthermia, some cells in the piriform cortex were shrunken during hypothermia and the neuropil was more compact than in normothermic conditions. Although some evidence of more compact neuropil was seen in other structures, these changes were within the normal range and not recognized as abnormal. Although the reasons behind a higher susceptibility of the piriform cortex to hypothermia-induced structural cell abnormalities remain unclear, this structure was especially prone to structural damage during environmental warming (145), temporal lobe epilepsy (146) as well as kainite (147) and urethane intoxications (148).
7. SUMMARY AND HUMAN HEALTH PERSPECTIVES
In contrast to general beliefs that brain temperature is a stable, tightly regulated homeostatic parameter, data presented in this review demonstrate a relatively wide range of fluctuations (~3°C) occurring in rats following exposure to salient somato-sensory stimuli and during various motivated behaviors. Although rats and humans have obvious differences in temperature regulation (149) and human data are very limited, similarity between rats and monkeys in general patterns of brain temperature fluctuations (6) suggests applicability of animal data to human conditions. It is true that humans have very efficient mechanisms of heat dissipation that allow them to loose more heat than can potentially be released, but under specific real-life conditions these mechanisms could be blocked, resulting in pathological brain hyperthermia. While intense physical exercise in professional cyclists increased brain temperature to about 1°C or less, the same exercise conducted in water-impermeable cloth, which prevented proper heat dissipation, resulted in about 2.5 or 3°C (up to 40°C) increases (53) that clearly exceed ‘normal’ physiological limits. Robust pathological hyperthermia and adverse, sometimes fatal, health complications could also occur in humans that take psychomotor stimulant drugs under the heat-compiling environmental conditions at “raves”. In addition to powerful metabolism-related heat production, these drugs have strong and prolonged vasoconstrictive effects, which prevent proper heat dissipation. Therefore, dangerous intra-brain heat accumulation and acute decompensation of physiological functions could occur in this case due to a combination of enhanced heat production induced by the drug and intense physical and psychoemotional activation and impaired heat dissipation due to drug-induced peripheral vasoconstriction, hot and humid environment, and cloth that prevents heat loss. The strength of potentiation of the hyperthermic effects by environmental conditions is really surprising. When ambient temperature was set at only ~6°C above laboratory standard (29°C), about 80% of rats died after exposure to both METH and MDMA at 1/5 of their LD50 (87, 88). While lethality resulting from acute intoxication with club drugs is not reported so often, it is a significant public health problem world wide because of the numbers of recreational drug users. According to global survey conduced the UN Office of Drugs and Crime, more than 500 tons of METH and MDMA were consumed in 2003, with more than 40 million people using them in the last 12 months (150). Therefore, the drug that is usually perceived as “safe” could be really dangerous.
Brain cells are exceptionally sensitive to heat, with some irreversible damage starting to occur at ~40°C, only about 3°C above normal baseline, and progressing exponentially with slight increases above these levels (151, 152). Although heat per se could selectively damage brain cells, hyperthermia also increases BBB permeability, thus allowing entry to the brain from peripheral circulation of various potentially neurotoxic substances, ions, and water. Therefore, damage of brain cells under conditions of hyperthermia reflects the effects of not only temperature, but multiple potentially dangerous influences. Robust leakage of the BBB under conditions of hyperthermia could be an important factor in brain entry of several small viruses and neurotoxic products of viral metabolism that are retained in the periphery under normal conditions. This effect could explain the unusually high incidence of neuro-AIDS in METH users as well as high co-morbidity of neuro-AIDS with malaria, a disease characterized by episodes of robust hyperthermia. High brain temperature could also promote brain entry of antibiotics that are usually retained by the BBB under normal conditions (153, 154). Some of these drugs are neurotoxic and its entry into brain environment could explain neurological complications seen in young children treated by these drugs for viral and bacterial infections. Therefore, temperature is an extremely important variable in both normal brain functioning and development of brain pathology.
ACKNOWLEDGEMENTS
This work was supported by the Intramural Research Program of the NIH, NIDA. Data described in this review were obtained in collaboration with Dr. Hari S. Sharma, P. Leon Brown, Robert Mitchum, David Bae, and Michael S. Smirnov, whose valuable contribution is greatly appreciated. I wish to thank Michael S. Smirnov and Mary Pfeiffer for editorial assistance, and Dr. Clark Blatteis for comments and suggestions made on the early variant of this review.
REFERENCES
- 1.Schmidt-Nielsen K. Adaptation and Environment. 5th Edition. Cambridge: Cambridge University Press; 1997. Animal Physiology. [Google Scholar]
- 2.James W. Briefer Course. New York: Henry Holt; 1892. Psychology. 1892. [Google Scholar]
- 3.Abrams R, Hammel HT. Hypothalamic temperature in unanesthetized albino rats during feeding and sleeping. Am. J. Physiol. 1964;206:641–646. doi: 10.1152/ajplegacy.1964.206.3.641. [DOI] [PubMed] [Google Scholar]
- 4.Delgado JMR, Hanai T. Intracerebral temperatures in free-moving cats. Am. J. Physiol. 1966;211:755–769. doi: 10.1152/ajplegacy.1966.211.3.755. [DOI] [PubMed] [Google Scholar]
- 5.Feitelberg S, Lampl H. Warmetonung der Grosshirnrinde bei Erregung und Ruhe Functionshemmung. Arch. Exp. Path. Pharmak. 1935;177:726–736. (in German) [Google Scholar]
- 6.Hayward JN, Baker MA. Role of cerebral arterial blood in the regulation of brain temperature in the monkey. Am. J. Physiol. 1968;215:389–403. doi: 10.1152/ajplegacy.1968.215.2.389. [DOI] [PubMed] [Google Scholar]
- 7.Kovalzon VM. Brain temperature variations during natural sleep and arousal in white rats. Physiol. Beh. 1972;10:667–670. doi: 10.1016/0031-9384(73)90141-8. [DOI] [PubMed] [Google Scholar]
- 8.McElligott JC, Melzack R. Localized thermal changes evoked in the brain by visual and auditory stimulation. Exp. Neurol. 1967;17:293–312. doi: 10.1016/0014-4886(67)90108-2. [DOI] [PubMed] [Google Scholar]
- 9.Serota HM. Temperature changes in the cortex and hypothalamus during sleep. J. Neurophysiol. 1939;2:42–47. [Google Scholar]
- 10.Serota HM, Gerard RW. Localized thermal changes in cat’s brain. J. Neurophysiol. 1938;1:115–124. [Google Scholar]
- 11.Siesjo B. Brain Energy Metabolism. New York: Wiley; 1978. [Google Scholar]
- 12.Gerasimov V. Information processing in human body. 1998 http://vadim.www.media.mit.edu/MAS862/Project.html.
- 13.Laughlin SB, de Ruyter van Steveninck RR, Anderson JC. The metabolic cost of neural information. Nature Neurosci. 1998;1:36–41. doi: 10.1038/236. [DOI] [PubMed] [Google Scholar]
- 14.Hodgkin AL. The Conduction of the Nervous Impulse. Liverpool: Liverpool University Press; 1967. [Google Scholar]
- 15.Ritchie JM. Energetic aspects of nerve conduction: The relationships between heat production, electrical activity and metabolism. Progr. Biophys. Molec. Biol. 1973;26:147–187. doi: 10.1016/0079-6107(73)90019-9. [DOI] [PubMed] [Google Scholar]
- 16.Shulman RG, Rothman DL, Behar KL, Hyder F. Energetic basis of brain activity: implications for neuroimaging. Trends Neurosci. 2004;27:489–495. doi: 10.1016/j.tins.2004.06.005. [DOI] [PubMed] [Google Scholar]
- 17.Sokoloff L. Energetics of functional activation in neural tissues. Neurochem Res. 1999;24:321–329. doi: 10.1023/a:1022534709672. (1999) [DOI] [PubMed] [Google Scholar]
- 18.Kiyatkin EA, Brown PL, Wise RA. Brain temperature fluctuation: a reflection of functional neural activation. Eur. J. Neurosci. 2002;16:164–168. doi: 10.1046/j.1460-9568.2002.02066.x. [DOI] [PubMed] [Google Scholar]
- 19.Fellows LK, Boutelle MG, Fillenz M. Physiological stimulation increases nonoxidative glucose metabolism in the brain of the freely moving rat. J. Neurochem. 1993;60:1258–1263. doi: 10.1111/j.1471-4159.1993.tb03285.x. [DOI] [PubMed] [Google Scholar]
- 20.Smirnov MS, Kiyatkin EA. Fluctuations in central and peripheral temperatures associated with feeding behavior in rats. Amer. J. Physiol. 2008;295:R1415–R1424. doi: 10.1152/ajpregu.90636.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kiyatkin EA, Bae D. Behavioral and brain temperature responses to salient environmental stimuli and intravenous cocaine in rats: Effects of diazepam. Psychopharmacology. 2008;196:343–356. doi: 10.1007/s00213-007-0965-y. [DOI] [PubMed] [Google Scholar]
- 22.Kiyatkin EA, Mitchum R R. Fluctuations in brain temperatures during sexual behavior in male rats: An approach for evaluating neural activity underlying motivated behavior. Neuroscience. 2003;119:1169–1183. doi: 10.1016/s0306-4522(03)00222-7. [DOI] [PubMed] [Google Scholar]
- 23.Altschule MD. Emotion and circulation. Circulation. 1951;3:444–454. doi: 10.1161/01.cir.3.3.444. [DOI] [PubMed] [Google Scholar]
- 24.Baker M, Cronin M M, Mountjoy D. Variability of skin temperature in the waking monkey. Am. J. Physiol. 1976;230:449–455. doi: 10.1152/ajplegacy.1976.230.2.449. [DOI] [PubMed] [Google Scholar]
- 25.Solomon GF, Moos RH, Stone GC GC, Fessel WJ WJ. Peripheral vasoconstriction induced by emotional stress in rats. Angiology. 1964;15:362–365. doi: 10.1177/000331976401500806. [DOI] [PubMed] [Google Scholar]
- 26.Kiyatkin RA, Wise RA. Striatal hyperthermia associated with arousal: intracranial thermorecordings in behaving rats. Brain Res. 2001;918:141–152. doi: 10.1016/s0006-8993(01)02985-7. [DOI] [PubMed] [Google Scholar]
- 27.Wilder J. The law of initial value in neurology and psychiatry; facts and problems. J. Nerv. Ment. Dis. 1957;125:73–86. doi: 10.1097/00005053-195701000-00009. [DOI] [PubMed] [Google Scholar]
- 28.Wilder J. Modern psychophysiology and the law of initial value. Am. J. Psychother. 1958;12:199–221. doi: 10.1176/appi.psychotherapy.1958.12.2.199. [DOI] [PubMed] [Google Scholar]
- 29.Horvath TL, Warden CH, Hajos M, Lombardi A, Goglia F, Diano S. Brain uncoupling protein 2: Uncoupled neuronal mitochondria predict thermal synapses in homeostatic centers. J. Neurosci. 1999;19:10417–10427. doi: 10.1523/JNEUROSCI.19-23-10417.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Mellergard P, Nordstrom CH. Epidural temperatures and possible intracerebral temperature gradients in man. Br. J. Neurosurg. 1990;4:31–38. doi: 10.3109/02688699009000679. [DOI] [PubMed] [Google Scholar]
- 31.Rumana CS, Gopinath SP, Uzura M, Valadka AB, Robertson CS. Brain temperatures exceed systemic temperatures in head-injured patients. Clin. Care Med. 1998;26:562–567. doi: 10.1097/00003246-199803000-00032. [DOI] [PubMed] [Google Scholar]
- 32.Schwab S, Spranger M, Aschoff A, Steiner T, Hacke W. Brain temperature monitoring and modulation in patients with severe MCA infarction. Neurology. 1997;48:762–767. doi: 10.1212/wnl.48.3.762. [DOI] [PubMed] [Google Scholar]
- 33.Andersen P, Moser EI. Brain temperature and hippocampal function. Hippocampus. 1995;5:491–498. doi: 10.1002/hipo.450050602. [DOI] [PubMed] [Google Scholar]
- 34.Moltz H. Fever: causes and consequences. Neurosci. Biobehav. Rev. 1993;17:237–269. doi: 10.1016/s0149-7634(05)80009-0. [DOI] [PubMed] [Google Scholar]
- 35.Moser EI, Mathiesen LI. Relationships between neuronal activity and brain temperature in rats. NeuroReport. 1996;7:1876–1880. doi: 10.1097/00001756-199607290-00038. [DOI] [PubMed] [Google Scholar]
- 36.Erikson K, Lanier W. Anesthetic technique influences brain temperature, independently of core temperature, during craniotomy in cats. Anesth. Analg. 2003;96:1460–1466. doi: 10.1213/01.ANE.0000061221.23197.CE. [DOI] [PubMed] [Google Scholar]
- 37.Kiyatkin EA, Rebec GV. Modulation of striatal neuronal activity by glutamate and GABA: Iontophoresis in awake, unrestrained rats. Brain Res. 1999;822:88–106. doi: 10.1016/s0006-8993(99)01093-8. [DOI] [PubMed] [Google Scholar]
- 38.Kiyatkin EA. Statistical analysis of impulse activity of diencephalic units during restraint stress. J. Higher Nerv. Act. 1982;32:502–510. [Google Scholar]
- 39.Kiyatkin EA. Functional properties of presumed dopamine-containing and other ventral tegmental area neurons in conscious rats. Int. J. Neurosci. 1988;42:21–43. doi: 10.3109/00207458808985756. [DOI] [PubMed] [Google Scholar]
- 40.Kiyatkin EA, Rebec GV. Heterogeneity of ventral tegmental area neurons: Single-unit recording and iontophoresis in awake, unrestrained rats. Neuroscience. 1998;85:1285–1309. doi: 10.1016/s0306-4522(98)00054-2. [DOI] [PubMed] [Google Scholar]
- 41.E.A. Kiyatkin EA, Rebec GV. Dopaminergic modulation of glutamate-induced excitations of neurons in the neostriatum and nucleus accumbens of awake, unrestrained rats. J. Neurophysiol. 1996;75:142–153. doi: 10.1152/jn.1996.75.1.142. [DOI] [PubMed] [Google Scholar]
- 42.Fox RT, Raichle ME. Focal physiological uncoupling of cerebral blood flow and oxidative metabolism during somatosensory stimulation in human subjects. Proc. Natl. Acad. Sci. 1986;83:1140–1144. doi: 10.1073/pnas.83.4.1140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Trubel HKF, Sacolick LI, Hyder F. Regional temperature changes in the brain during somatosensory stimulatio. J. Cereb. Blood Flow Metab. 2006;26:68–78. doi: 10.1038/sj.jcbfm.9600164. [DOI] [PubMed] [Google Scholar]
- 44.Sukstanskii AL, Yablonskiy DA. Theoretical model of temperature regulation in the brain during changes in functional activity. Proc. Natl. Acad. Sci. 2006;103:12144–12149. doi: 10.1073/pnas.0604376103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yablonskiy DA, Ackerman JH, Raichle ME. Coupling between changes in human brain temperature and oxidative metabolism during prolonged visual stimulation. Proc. Nat. Acad. Aci. 2000;97:7603–7608. doi: 10.1073/pnas.97.13.7603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Charkoudian N. Skin blood flow in adult human thermoregulation: how it works, when it does not, and why? Mayo Clin. Proc. 2003;78:603–612. doi: 10.4065/78.5.603. [DOI] [PubMed] [Google Scholar]
- 47.Ryan KL, Taylor WF, Bishop VS. Arterial baroreflex modulation of heat-induced vasodilation in the rabbit ear. J. Appl. Physiol. 1997;83:2091–2097. doi: 10.1152/jappl.1997.83.6.2091. [DOI] [PubMed] [Google Scholar]
- 48.Oobu K. Experimental studies on the effect of heating on blood flow in the tongue of golden hamsters. Fukuoka Igaku Zasshi. 1993;84:497–511. (1993) [PubMed] [Google Scholar]
- 49.Nagata Y, Katayama K, Manivel CJ, Song CW. Changes in blood flow in locally heated intestine of rats. Int. J. Hyperthermia. 2000;16:159–170. doi: 10.1080/026567300285358. [DOI] [PubMed] [Google Scholar]
- 50.Nakajima T, Rhee JC, Song CW, Onoyama Y. Effect of a second heating on rat liver blood flow. Int. J. Hyperthermia. 1992;8:679–687. doi: 10.3109/02656739209038003. [DOI] [PubMed] [Google Scholar]
- 51.Moriyama E. Cerebral blood flow changes during localized hyperthermia. Neurol. Med. Chir. (Tokio) 1990;30:923–929. doi: 10.2176/nmc.30.923. [DOI] [PubMed] [Google Scholar]
- 52.Uda M, Tanaka Y. Arterial blood flow changes after hyperthermia on normal liver, normal brain, and normal small intestine. Gan No. Rinsho. 1990;36:2362–2366. [PubMed] [Google Scholar]
- 53.Nybo L, Secher NH, Nielson B. Inadequate heat release from the human brain during prolonged exercise with hyperthermia. J. Physiol. 2002;545:697–704. doi: 10.1113/jphysiol.2002.030023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Margaria R, Cretelli P, Aghemo P, Sassi G. Energy cost of running. J. Appl. Physiol. 1963;18:367–370. doi: 10.1152/jappl.1963.18.2.367. [DOI] [PubMed] [Google Scholar]
- 55.Donaldson GC, Keatinge WR, Saunders RD. Cardiovascular responses to heat stress and their adverse consequences in healthy and vulnerable human populations. Int. J. Hyperthermia. 2003;19:225–235. doi: 10.1080/0265673021000058357. [DOI] [PubMed] [Google Scholar]
- 56.Rowell LB. Cardiovascular aspects of human thermoregulation. Circ. Res. 1983;52:367–376. doi: 10.1161/01.res.52.4.367. [DOI] [PubMed] [Google Scholar]
- 57.Cheuvront SN, Haymes EM. Thermoregulation and marathon running: biological and environmental influences. Sports Med. 2001;31:743–762. doi: 10.2165/00007256-200131100-00004. [DOI] [PubMed] [Google Scholar]
- 58.Schaefer CF. Possible teratogenic hyperthermia and marathon running. JAMA. 1979;241:1892. [PubMed] [Google Scholar]
- 59.Brengelmann GL. Specialized brain cooling in humans? FASEB J. 1993;7:1148–1153. doi: 10.1096/fasebj.7.12.8375613. [DOI] [PubMed] [Google Scholar]
- 60.Cabanac M. Selective brain cooling in humans: “fancy” or fact? FASEB J. 1993;7:1143–1147. doi: 10.1096/fasebj.7.12.8375612. [DOI] [PubMed] [Google Scholar]
- 61.Cabanac M, Caputa M. Natural selective cooling of the human brain: evidence of its occurrence and magnitude. J. Physiol. 1979;286:255–264. doi: 10.1113/jphysiol.1979.sp012617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Simon E. Tympanic temperature is not suited to indicate selective brain cooling in humans: a re-evaluation of the thermohysiological basics. Eur. J. App. Physiol. 2007;101:19–30. doi: 10.1007/s00421-007-0449-0. [DOI] [PubMed] [Google Scholar]
- 63.Brinnel H, Nagasaka T, Cabanac M. Enhanced brain protection during passive hyperthermia in humans. Eur. J. Appl. Physiol. 1987;56:540–545. doi: 10.1007/BF00635367. [DOI] [PubMed] [Google Scholar]
- 64.Cabanac M, Germain M, Brinnel H. Tympanic temperatures during hemiface cooling. Eur. J. Appl. Physiol. 1987;56:534–539. doi: 10.1007/BF00635366. [DOI] [PubMed] [Google Scholar]
- 65.Mariak Z, Jadeszko M, Lewko J, Lebkowski W, Lyson T. No specific brain protection against thermal stress in fever. Acta Neurochir. (Wien) 1998;140:585–590. doi: 10.1007/s007010050144. [DOI] [PubMed] [Google Scholar]
- 66.Mariak Z, Lebkowski W, Lyson T, Lewko J, Piekarski P. Brain temperature during craniotomy in general anesthesia. Neurol. Neurochir. Pol. 1999;33:1325–1327. [PubMed] [Google Scholar]
- 67.Mariak Z, Lyson T, Peikarski P, Lewko J, Jadeszko M, Szydlik P. Brain temperature in patients with central nervous system lesions. Neurol. Neurosurg. Pol. 2000;34:509–522. [PubMed] [Google Scholar]
- 68.Nybo L, Nielson N. Middle cerebral artery blood velocity is reduced with hyperthermia during prolonged exercise in humans. J. Physiol. 2001;534:279–286. doi: 10.1111/j.1469-7793.2001.t01-1-00279.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ide K, Secher NH. Cerebral blood flow and metabolism during exercise. Prog. Neurobiol. 2000;61:397–414. doi: 10.1016/s0301-0082(99)00057-x. [DOI] [PubMed] [Google Scholar]
- 70.Ide K, Schmalbruch IK, Quistorff B, Horn A, Secher NH. Lactate, glucose, and oxygen uptake in human brain during recovery from maximal exercise. J. Physiol. 2000;522:159–164. doi: 10.1111/j.1469-7793.2000.t01-2-00159.xm. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Walters TJ, Rynan KL, Tate LM, Mason PA. Exercise in the heat is limited by a critical internal temperature. J. Appl. Physiol. 2000;89:799–806. doi: 10.1152/jappl.2000.89.2.799. [DOI] [PubMed] [Google Scholar]
- 72.Nielsen B, Hyldig T, Bidsrup F, Gonzalez-Alonso J, Christoffersen CR. Brain activity and fatigue during prolonged exercise in the heat. Pflugers Arch. 2001;442:41–48. doi: 10.1007/s004240100515. [DOI] [PubMed] [Google Scholar]
- 73.Estler CJ. Dependence on age of metamphetamine-produced changes in thermoregulation and metabolism. Experientia. 1975;31:1436–1437. doi: 10.1007/BF01923231. [DOI] [PubMed] [Google Scholar]
- 74.Makisumi T, Yoshida K, Watanabe T, Tan N, Murakami N, Morimoto A. Sympatho-adrenal involvement in methamphetamine-induced hyperthermia through skeletal muscle hypermethanolism. Eur. J. Pharmacol. 1998;363:107–112. doi: 10.1016/s0014-2999(98)00758-4. [DOI] [PubMed] [Google Scholar]
- 75.Alberts DS, Sonsalla PK. Methamphetamine-induced hyperthermia and dopaminergic neurotoxicity in mice: Pharmacological profile of protective and nonprotective agents. J. Pharmacol. Exp. Ther. 1995;275:1104–1114. [PubMed] [Google Scholar]
- 76.Green AR, Mechan AO, Elliott JM, O’Shea E, Colado MI. The pharmacology and clinical pharmacology of 3,4-methylenedioxymethamphetamine (MDMA, “Ecstasy”) Pharmacol Rev. 2003;55:463–508. doi: 10.1124/pr.55.3.3. [DOI] [PubMed] [Google Scholar]
- 77.Mechan AO, O’Shea E, Elliot JM, Colado MI, Green AR. A neurotoxic dose of 3,4-methylenedioxymethamphetamine (MDMA; ecstasy) to rats results in a long-term deficit in thermoregulation. Psychopharmacology. 2001;155:413–418. doi: 10.1007/s002130100735. [DOI] [PubMed] [Google Scholar]
- 78.Miller DB, O’Callaghan JP. Environment-, drug- and stress-induced alterations in body temperature affect the neurotoxicity of substituted amphetamines in the C57BL/6J mouse. J. Pharmacol. Exp. Ther. 1994;270:752–760. [PubMed] [Google Scholar]
- 79.Sandoval V, Hanson GR, Fleckenstein AE. Methamphetamine decreases mouse striatal dopamine transporter activity: roles of hyperthermia and dopamine. Eur. J. Pharmacol. 2000;409:265–271. doi: 10.1016/s0014-2999(00)00871-2. [DOI] [PubMed] [Google Scholar]
- 80.Gordon CJ, Watkinson WO, O’Callaghan JP, Miller DB. Effects of 3,4-methylenedioxymethamphetamine on autonomic thermoregulatory responses of the rat. Pharmacol. Biochem. Behav. 1991;38:339–344. doi: 10.1016/0091-3057(91)90288-d. [DOI] [PubMed] [Google Scholar]
- 81.Pederson NP, Blessing WW. Cutaneous vasoconstriction contributes to hyperthermia induced by 3,4-methylenedioxymethamphetamine (ecstasy) on conscious rabbits. J. Neurosci. 2001;21:8648–8654. doi: 10.1523/JNEUROSCI.21-21-08648.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ali SF, Newport GD, Slikker W. Methamphetamine-induced dopaminergic toxicity in mice: role of environmental temperature and pharmacological agents. Ann. N.Y. Acad. Sci. 1996;801:187–198. doi: 10.1111/j.1749-6632.1996.tb17441.x. [DOI] [PubMed] [Google Scholar]
- 83.Davidson C, Gow AJ, Lee TH, Ellinwood EH. Methamphetamine neurotoxicity: necrotic and apoptotic mechanisms and relevance to human abuse and treatment. Brain Res. Rev. 2001;36:1–22. doi: 10.1016/s0165-0173(01)00054-6. [DOI] [PubMed] [Google Scholar]
- 84.Kalant H. The pharmacology and toxicology of “ecstasy” (MDMA) and related drugs. Can. Med. Assoc. J. 2001;165:917–928. [PMC free article] [PubMed] [Google Scholar]
- 85.Schmued LC. Demonstration and localization of neuronal degeneration in the rat forebrain following a single exposure to MDMA. Brain Res. 2003;974:127–133. doi: 10.1016/s0006-8993(03)02563-0. [DOI] [PubMed] [Google Scholar]
- 86.Brown PL, Wise RA, Kiyatkin EA. Brain hyperthermia is induced by methamphetamine and exacerbated by social interaction. J. Neurosci. 2003;23:3924–3929. doi: 10.1523/JNEUROSCI.23-09-03924.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Brown PL, Kiyatkin EA. Brain hyperthermia induced by MDMA (“ecstasy”): modulation by environmental conditions. Eur. J. Neurosci. 2004;20:51–58. doi: 10.1111/j.0953-816X.2004.03453.x. [DOI] [PubMed] [Google Scholar]
- 88.Brown PL, Kiyatkin EA. Fatal intra-brain heat accumulation induced by meth-amphetamine at normothermic conditions in rats. Int. J. Neurodegener. Neuroregener. 2005;1:86–90. [Google Scholar]
- 89.Romanovsky AA, Ivanov AI, Shimansky YP. Ambient temperature for experiments in rats: a new method for determining the zone of thermal neutrality. J. Appl. Physiol. 2002;92:2667–2679. doi: 10.1152/japplphysiol.01173.2001. [DOI] [PubMed] [Google Scholar]
- 90.Kiyatkin EA, Brown PL. Modulation of physiological brain hyperthermia by environmental temperature and impaired blood outflow in rats. Physiol. Behav. 2004;83:467–474. doi: 10.1016/j.physbeh.2004.08.032. [DOI] [PubMed] [Google Scholar]
- 91.Davis WM, Hatoum HT, Walters IW. Toxicity of MDA (2,4-methylenedioxyamphetamine) considered for relevance to hazards of MDMA (Ecstasy) abuse. Alcohol Drug Res. 1987;7:123–134. [PubMed] [Google Scholar]
- 92.Mitchum R, Kiyatkin EA. Brain hyperthermia and temperature fluctuations during sexual interaction in female rats. Brain Res. 2004;1000:110–122. doi: 10.1016/j.brainres.2003.12.024. [DOI] [PubMed] [Google Scholar]
- 93.Kiyatkin EA, Wise RA. Brain and body hyperthermia associated with heroin self-administration in rats. J. Neurosci. 2002;22:1072–1080. doi: 10.1523/JNEUROSCI.22-03-01072.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Kiyatkin EA, Brown PL. Fluctuations in neural activity during cocaine self-administration: Clues provided by brain thermorecording. Neuroscience. 2003;116:525–538. doi: 10.1016/s0306-4522(02)00711-x. [DOI] [PubMed] [Google Scholar]
- 95.Swan H. Thermoregulation and Bioenergetics. New York: Elsevier; 1974. [Google Scholar]
- 96.Lee JCF, Callaway JC, Foehring RC. The effects of temperature on calcium transients and Ca2+-dependent afterhyperpolarizations in neocortical pyramidal neurons. J. Physiol. 2005;93:2012–2020. doi: 10.1152/jn.01017.2004. [DOI] [PubMed] [Google Scholar]
- 97.Thompson SM, Musakawa LM, Rince DA. Temperature dependence of intrinsic membrane properties and synaptic potentials in hippocampal CA1 neurons in vitro. J. Neurosci. 1985;5:817–824. doi: 10.1523/JNEUROSCI.05-03-00817.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Tryba AK, Ramirez J-M. Hyperthermia modulates respiratory pacemaker bursting properties. J. Neurophysiol. 2004;92:2844–2852. doi: 10.1152/jn.00752.2003. [DOI] [PubMed] [Google Scholar]
- 99.Volgushev M, Vidyasagar TR, Chistiakova M, Eysel UT. Synaptic transmission in the neocortex during reversible cooling. Neuroscience. 2000;98:9–22. doi: 10.1016/s0306-4522(00)00109-3. [DOI] [PubMed] [Google Scholar]
- 100.Katz B, Miledi R. The effect of temperature on the synaptic delay at the neuromuscular junction. J. Physiol. 1965;181:656–670. doi: 10.1113/jphysiol.1965.sp007790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Berner NJ, Heller HC. Does the preoptic anterior hypothalamus receive thermoafferent information? Am. J. Physiol. 1998;274:R9–R18. doi: 10.1152/ajpregu.1998.274.1.R9. [DOI] [PubMed] [Google Scholar]
- 102.Boulant JA. Role of the preoptic-anterior hypothalamus in thermoregulation and fever. Clin. Infect. Dis. 2000;31(Suppl 5):S157–S161. doi: 10.1086/317521. [DOI] [PubMed] [Google Scholar]
- 103.Nadel E. Regulation of body temperature. In: Boron WF, Boulpaep EL, editors. Medical Physiology. Philadelphia: Saunders; 2003. pp. 1231–1241. [Google Scholar]
- 104.Travis KA, Bockholt HJ, Zardetto-Smith AM, Johnson AK. In vitro thermosensitivity of the midline thalamus. Brain Res. 1995;686:17–22. doi: 10.1016/0006-8993(95)00428-s. [DOI] [PubMed] [Google Scholar]
- 105.Burgoon PW, Boulant JA. Temperature-sensitive properties of rat suprachiasmatic nucleus neurons. Am. J. Physiol. 2001;281:R706–R715. doi: 10.1152/ajpregu.2001.281.3.R706. [DOI] [PubMed] [Google Scholar]
- 106.Ruby NF, Heller HC. Temperature sensitivity of the suprachiasmatic nucleus of ground squirrels and rats in vitro. J. Biol. Rhythms. 1996;11:126–136. doi: 10.1177/074873049601100205. [DOI] [PubMed] [Google Scholar]
- 107.Guatteo E, Chung KK, Bowala TK, Bernardi G, Mercuri ND, Lipski J J. Temperature sensitivity of dopaminergic neurons of the substantia nigra pars compacta: involvement of transient receptor potential channels. J Neurophysiol. 2005;94:3069–3080. doi: 10.1152/jn.00066.2005. [DOI] [PubMed] [Google Scholar]
- 108.Nakashima K, Todd MM. Effects of hypothermia on the rate of excitatory amino acid release after ischemic depolarization. Stroke. 1996;27:913–918. doi: 10.1161/01.str.27.5.913. [DOI] [PubMed] [Google Scholar]
- 109.Vizi ES. Different temperature dependence of carrier-mediated (cytoplasmic) and stimulus-evoked (exocytotic) release of transmitter: a simple method to separate the two types of release. Neurochem. Int. 1998;33 doi: 10.1016/s0197-0186(98)00040-0. 359-356. [DOI] [PubMed] [Google Scholar]
- 110.Xie T, McGann UD, Kim S, Yuan J, Ricaurte GA. Effect of temperature on dopamine transporter function and intracellular accumulation of methamphetamine: Implications for methamphetamine-induced dopaminergic neurotoxicity. J. Neurosci. 2000;20:7838–7845. doi: 10.1523/JNEUROSCI.20-20-07838.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Arai H, Uto A, Ogawa Y, Sato K K. Effect of low temperature on glutamate-induced intracellular calcium accumulation and cell death in cultured hippocampal neurons. Neurosci. Lett. 1993;163:132–134. doi: 10.1016/0304-3940(93)90363-p. [DOI] [PubMed] [Google Scholar]
- 112.Lucas JH, Emery DG, Wang G, Rosenberg-Schaffer LJ, Jordan RS, Gross GW. In vitro investigation of the effect of nonfreezing low temperatures on injured and uninjured mammalian spinal neurons. J. Neurotrauma. 1994;11:35–61. doi: 10.1089/neu.1994.11.35. [DOI] [PubMed] [Google Scholar]
- 113.Iwagami Y. Changes in ultrastructure of human cell related to certain biological responses under hyperthermic culture condition. Human Cell. 1996;9:353–366. [PubMed] [Google Scholar]
- 114.Willis WT, Jackman MR, Bizeau ME, Pagliassotti MJ, Hazel JR. Hyperthermia impairs liver mitochondrial functions. Am. J. Physiol. 2000;278:R1240–R1246. doi: 10.1152/ajpregu.2000.278.5.R1240. [DOI] [PubMed] [Google Scholar]
- 115.Chen YZ, Xu RX, Huang QJ, Xu ZJ, Jiang XD, Cai YO. Effect of hyperthermia on tight junctions between endothelial cells of the blood-brain barrier model in vitro. Di Yi Jun Da Xue Xue Bao. 2003;23:21–24. [PubMed] [Google Scholar]
- 116.Oifa AI, Kleshchnov VN. Ultrastructural analysis of the phenomenon of acute neuronal swelling. Zh. Nevropatol. Psikhiatr Im S.S. Korsakova. 1985;85:1016–1020. [PubMed] [Google Scholar]
- 117.Lee SY, Lee SH, Akuta K, Uda M, Song CW. Acute histological effects of interstitial hyperthermia on normal rat brain. Int. J. Hyperthermia. 2000;16:73–83. doi: 10.1080/026567300285439. [DOI] [PubMed] [Google Scholar]
- 118.Li Y-Q, Chen P, Jain V, Reilly RM, Wong CS. Early radiation-induced endothelial cell loss and blood-spinal cord barrier breakdown in the rat spinal cord. Radiat. Res. 2004;161:143–152. doi: 10.1667/rr3117. [DOI] [PubMed] [Google Scholar]
- 119.Lin PS, Quamo S, Ho KC, Gladding J. Hyperthermia enhances the cytotoxic effects of reactive oxygen species to Chinese hamster cells and bovine endothelial cells in vitro. Radiat. Med. 1991;126:43–51. [PubMed] [Google Scholar]
- 120.Bechtold DA, Brown IR. Induction of Hsp27 and Hsp32 stress proteins and vimentin in glial cells of the rat hippocampus following hyperthermia. Neurochem. Res. 2003;28:1163–1173. doi: 10.1023/a:1024268126310. [DOI] [PubMed] [Google Scholar]
- 121.Sharma HS, Hoopes PJ. Hyperthermia-induced pathophysiology of the central nervous system. Int. J. Hyperthermia. 2003;19:325–354. doi: 10.1080/0265673021000054621. [DOI] [PubMed] [Google Scholar]
- 122.Cervos-Navarro J, Sharma HS, Westman J, Bongcum-Rudloff E. Glial cell reactions in the central nervous system following heat stress. Progr. Brain Res. 1998;115:241–274. doi: 10.1016/s0079-6123(08)62039-7. [DOI] [PubMed] [Google Scholar]
- 123.Lin M-T. Heatstroke-induced cerebral ischemia and neuronal damage. Involvement of cytokines and monoamines. Ann. N.Y. Acad. Sci. 1997;813:572–580. doi: 10.1111/j.1749-6632.1997.tb51748.x. [DOI] [PubMed] [Google Scholar]
- 124.Sharma HS, Zimmer C, Westman J, Cervos-Navarro J. Acute systemic heat stress increases glial fibrillary acidic protein immunoreactivity in brain. An experimental study in the conscious normotensive young rats. Neuroscience. 1992;48:889–901. doi: 10.1016/0306-4522(92)90277-9. [DOI] [PubMed] [Google Scholar]
- 125.Kiyatkin EA, Brown PL, Sharma HS. Brain edema and breakdown of the blood-brain barrier during methamphetamine intoxication: Critical role of brain temperature. Eur. J. Neurosci. 2007;26:1242–1253. doi: 10.1111/j.1460-9568.2007.05741.x. [DOI] [PubMed] [Google Scholar]
- 126.Sharma HS, Kiyatkin EA. Rapid morphological brain abnormalities during acute methamphetamine intoxication in the rat: An experimental study using light and electron microscopy. J Chem. Neuroanat. doi: 10.1016/j.jchemneu.2008.08.002. Xx, xx-xx. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Bowyer JF, Ali S. High doses of methamphetamine that cause disruption of the blood-brain barrier in limbic regions produce extensive neuronal degeneration in mouse hippocampus. Synapse. 2006;60:521–532. doi: 10.1002/syn.20324. [DOI] [PubMed] [Google Scholar]
- 128.Sharma HS, Ali SF. Alterations in blood-brain barrier function by morphine and methamphetamine. Ann. N.Y. Acad. Sci. 2006;1074:198–224. doi: 10.1196/annals.1369.020. [DOI] [PubMed] [Google Scholar]
- 129.Watson P, Shirreffs SM, Maughan RJ. Blood-brain barrier integrity may be threatened by exercise in a warm environment. Am. J. Physiol. 2005;288:R1689–R1694. doi: 10.1152/ajpregu.00676.2004. [DOI] [PubMed] [Google Scholar]
- 130.Esposito P, Cheorghe D, Kendere K, Pang X, Connoly R, Jaconson S, Theodorides TC. Acute stress increases permeability of the blood-brain barrier through activation of brain must cells. Brain Res. 2001;888:117–127. doi: 10.1016/s0006-8993(00)03026-2. [DOI] [PubMed] [Google Scholar]
- 131.Ovadia H, Abramsky O, Feldman S, Weidenfeld J. Evaluation of the effects of stress on the blood-brain barrier: critical role of the brain perfusion time. Brain Res. 2001;905:21–25. doi: 10.1016/s0006-8993(01)02361-7. [DOI] [PubMed] [Google Scholar]
- 132.Sharma HS, Dey PK. Influence of long-term immobilization stress on regional blood-brain permeability, cerebral blood flow and 5-HT levels in conscious normotensive young rats. J. Neurol. Sci. 1986;72:61–76. doi: 10.1016/0022-510x(86)90036-5. [DOI] [PubMed] [Google Scholar]
- 133.Kiyatkin EA. Brain hyperthermia as physiological and pathological phenomena. Brain Res. Rev. 2005;50:27–56. doi: 10.1016/j.brainresrev.2005.04.001. [DOI] [PubMed] [Google Scholar]
- 134.Cadet JL, Thiriet N, Jayanthi S. Involvement of free radicals in MDMA-induced neurotoxicity in mice. Ann. Med. Intern. 2001;152(Suppl. 3):IS57–IS59. [PubMed] [Google Scholar]
- 135.Nybo L. Hyperthermia and fatigue. J. Appl. Physiol. 2008;104:871–877. doi: 10.1152/japplphysiol.00910.2007. [DOI] [PubMed] [Google Scholar]
- 136.Sharma HS. Hyperthermia-induced brain edema: Current status and future perspectives. Indian J. Med. Res. 2006;123:629–652. [PubMed] [Google Scholar]
- 137.Haorah J, Ramirez SH, Schall K, Smith D, Pandya R, Persidsky Y. Oxidative stress activates protein kinase and matrix metalloproteinases leading to blood-brain barrier disfunction. J. Neurochem. 2007;101:566–576. doi: 10.1111/j.1471-4159.2006.04393.x. [DOI] [PubMed] [Google Scholar]
- 138.Hom S, Egleton RD, Huber JD, Davis TP. Effect of reduced flow on blood-brain barrier transport systems. Brain Res. 2001;890:38–48. doi: 10.1016/s0006-8993(00)03027-4. [DOI] [PubMed] [Google Scholar]
- 139.Kaur C, Ling EA. Blood brain barrier in hypoxic-ischemic conditions. Curr. Neurovasc. Res. 2008;5:71–81. doi: 10.2174/156720208783565645. [DOI] [PubMed] [Google Scholar]
- 140.Kiyatkin EA, Brown PL. Brain and body temperature homeostasis during sodium pentobarbital anesthesia with and without body warming in rats. Physiol. Behav. 2005;84:563–570. doi: 10.1016/j.physbeh.2005.02.002. [DOI] [PubMed] [Google Scholar]
- 141.Rapoport SI. Blood-brain Barrier in Physiology and Medicine. New York: Raven Press; 1976. [Google Scholar]
- 142.Zlokovic BV. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron. 2008;57:178–201. doi: 10.1016/j.neuron.2008.01.003. [DOI] [PubMed] [Google Scholar]
- 143.Crane P, Braun L, Cornford E, Cremer J, Glass J, Oldendorf M. Dose-dependent reduction of glucose utilization by pentobarbital in rat brain. Stroke. 1978;9:12–18. doi: 10.1161/01.str.9.1.12. [DOI] [PubMed] [Google Scholar]
- 144.Penicaud L, Ferre P, Kande J, Leturque A, Issad T, Girard J. Effect of anesthesia on glucose production and utilization in rats. Am. J. Physiol. 1987;252:E365–E369. doi: 10.1152/ajpendo.1987.252.3.E365. [DOI] [PubMed] [Google Scholar]
- 145.Sharma HS, Cervos-Navarro J, Dey PK. Acute heat exposure causes cellular alterations in cerebral cortex of young rats. NeuroReport. 1991;2:155–158. doi: 10.1097/00001756-199103000-00012. [DOI] [PubMed] [Google Scholar]
- 146.Van Vliet EA, da Costa Araujo S, Redeker A, van Schalk C, Aronica E, Gorter JA. Brain-blood leakage may lead to progression of temporal lobe epilepsy. Brain. 2007;130:521–534. doi: 10.1093/brain/awl318. [DOI] [PubMed] [Google Scholar]
- 147.Candelario-Jalil E, Al-Dalain SM, Castello R, Martinez G, Fernandez OS. Selective vulnerability to kainite-induced oxidative damage in different rat brain regions. J. Appl. Toxicol. 2001;21:403–407. doi: 10.1002/jat.768. [DOI] [PubMed] [Google Scholar]
- 148.Thompson KW, Wasterlain CG. Urethane anesthesia produces selective damage in the piriform cortex of the developing brain. Dev. Brain Res. 2001;130:167–171. doi: 10.1016/s0165-3806(01)00228-0. [DOI] [PubMed] [Google Scholar]
- 149.Gordon CJ, Spencer PJ, Hotchkiss J, Miller DB, Hinderliter PM, Pauluhn J. Thermoregulation and its influence on toxicity assessment. Toxicology. 244:87–97. doi: 10.1016/j.tox.2007.10.030. [DOI] [PubMed] [Google Scholar]
- 150.United Nations Office on Drugs and Crime: Ecstasy and amphetamines: Global Survey 2003. Vienna: United Nations Publications; [Google Scholar]
- 151.Lepock JR. Cellular effects of hyperthermia: relevance to the minimum dose for thermal damage. Int. J. Hyperthermia. 2003;19:252–266. doi: 10.1080/0265673031000065042. [DOI] [PubMed] [Google Scholar]
- 152.Lepock JR. How do cells respond to their thermal environment? Int. J. Hyperthermia. 2005;21:681–687. doi: 10.1080/02656730500307298. [DOI] [PubMed] [Google Scholar]
- 153.Kearney BP, Aweeka FT. The penetration of anti-infectives into the central nervous system. Neurol. Clin. 17:883–900. doi: 10.1016/s0733-8619(05)70171-7. [DOI] [PubMed] [Google Scholar]
- 154.Lutsar I, Friedland IR. Pharmacokinetics and pharmacodynamics of cephalosporins in cerebrospinal fluid. Clin. Pharmacokinet. 39:335–343. doi: 10.2165/00003088-200039050-00003. [DOI] [PubMed] [Google Scholar]