

Abstract
Presence of an abnormal form a host-encoded prion protein (PrPC) that is protease resistant, pathologic and infectious characterizes prion diseases such as Chronic Wasting Disease (CWD) of cervids and scrapie in sheep. The Prion hypothesis asserts that this abnormal conformer constitutes most or all of the infectious prion. The role of the immune system in early events in peripheral prion pathogenesis has been convincingly demonstrated for CWD and scrapie 1-3. Transgenic and pharmacologic studies in mice revealed an important role of the Complement system in retaining and replicating prions early after infection 4-6. In vitro and in vivo studies have also observed prion retention by dendritic cells 7-10, although their role in trafficking remains unclear 11-16. Macrophages have similarly been implicated in early prion pathogenesis, but these studies have focused on events occurring weeks after infection 3,11,17. These prior studies also suffer from the problem of differentiating between endogenous PrPC and infectious prions. Here we describe a semiquantitative, unbiased approach for assessing prion uptake and trafficking from the inoculation site by immune cells recruited there. Aggregated prion rods were purified from infected brain homogenate by detergent solubilization of non-aggregated proteins and ultracentrifugation through a sucrose cushion. Polyacrylamide gel electrophoresis, coomassie blue staining and western blotting confirmed recovery of highly enriched prion rods in the pelleted fraction. Prion rods were fluorochrome-labeled then injected intraperitoneally into mice. Two hours later immune cells from peritoneal lavage fluid, spleen and mediastinal and mesenteric lymph nodes were assayed for prion rod retention and cell subsets identified by multicolor flow cytometry using markers for monocytes, neutrophils, dendritic cells, macrophages and B and T cells. This assay allows for the first time direct monitoring of immune cells acquiring and trafficking prions in vivo within hours after infection. This assay also clearly differentiates infectious, aggregated prions from PrPC normally expressed on host cells, which can be difficult and lead to data interpretation problems in other assay systems. This protocol can be adapted to other inoculation routes (oral, intravenous, intranervous and subcutaneous, e.g.) and antigens (conjugated beads, bacterial, viral and parasitic pathogens and proteins, egg) as well.
Protocol
1. Purifying and Labeling Prion Rods
This protocol is adapted from one previously published 18
Homogenize 100 grams of prion-infected brain tissue in 900 mL of ice cold homogenizing buffer (HB, 1X PBS containing 320 mM sucrose, 150 mM NaCl and 4mM EDTA) 1 min at maximum speed in a commercial blender, then 2 min on ice. Repeat three times.
Centrifuge homogenate for 10 min at 3000 x g and 4°C. Remove and save supernatant on ice. Re-suspend pellet in 1L HB and repeat steps 1.1 and 1.2.
Pool supernatants and centrifuge for 60 min at 100,000 x g, 0°C. Discard supernatant.
Re-suspend pellet in 100 mL of 1X Tris buffered saline (10 mM Tris-HCl, 0.1 mM NaCl and 1 mM EDTA) containing 2% Triton X-100 (TBST). Estimate protein concentration by the Bradford or similar assay and adjust to 5 mg/ mL in TBST. Chill on ice for 30 min.
Centrifuge for 30 min at 100,000 x g, 0°C. Discard supernatant and wash pellet with 50 mL TBST then 100 mL TBS. Re-suspend pellet in 100 mL 1X PBS containing 1% sarcosyl and protease inhibitor cocktail and stir for 120 min at 37°C.
Layer sample on 900 mL of 320 mM sucrose and centrifuge for 60 min at 100,000 x g, 10°C. Discard supernatant and re-suspend pellet in 10 mL 2.3M NaCl, 5% sarcosyl.
Sonicate sample 5 x 40 seconds at 70% maximum power at 1 min intervals on ice.
Aliquot 1 mL of sample into eppendorf tubes and centrifuge for fifteen minutes at 13,000 x g, 4oC. Wash pellets twice with TBS and store @ -70°C or conjugate to fluorochrome in step 1.9.
Conjugate 1 tube of purified prion rods using 1 vial of DyLight 649 fluorochrome according to manufacturer's protocol.
Exchange DyLight labeling buffer for PBS by dialysis or centrifugation through filter columns. Store in 20 μL aliquots at -70°C away from light until ready to use.
2. Prion Inoculation and Cell Recovery
This section encompasses intraperitoneal prion inoculation and recovery of cells from the peritoneum, mediastinal and mesenteric lymph nodes and spleen.
Dilute fluorescent prion rods 1:50 in PBS immediately prior to use. Scruff mouse by the dorsal neck fur with thumb and index finger, gently turn mouse to face you and restrain tail with pinky finger. Supinate mouse, elevating hind torso above head and holding him slightly upside down.
Using a 30G insulin syringe, inject 100 μL of fluorescent prion rods 1 cm lateral of the midline and 1-2 cm anterior to the sacroiliac joint. Insert the needle 1 cm through the skin directed 45° medially. Use 1:50 dilution of fluorescent beads or PBS alone as controls. Incubate mice for allotted time before analysis.
Euthanize mice according to approved animal care and use committee protocols. Inject 10 mL of PBS into the peritoneal cavity using a 12 mL syringe and 25G needle. Gently rock carcass longitudinally for one minute. Recover cell suspension from peritoneum using the same syringe and a 16G needle into a 15 mL conical tube and reserve on ice while carcass dissection continues.
Wet the ventral side of the carcass thoroughly with 70% ethanol. Lift the skin at the midline just below the ribcage with forceps and cut a small incision through the skin with scissors. Starting there, make a 3-5 cm long incision through the skin from in both directions to head and tail, then two 2-cm incisions from the midline laterally from each end. Peel back and pin the skin to reveal the peritoneal and thoracic cavities. Carefully cut through the thin translucent membranes surrounding the peritoneum and reflect the thorax to reveal the mediastinum.
Find and remove 2-4 mediastinal lymph nodes, slightly dorsal and laterally adjacent to the thymus, medially adjacent to the brachiocephalic artery, near the ventral side of the trachea. Find and remove up to six mesenteric lymph nodes, embedded in the soft white mesenteric tissue that anchors the intestines to the posterior abdominal wall. Find and remove the spleen, a long, dark red organ located just lateral of the midline and dorsal to the intestines on the left side of the abdomen. Reserve lymph nodes in 1 mL RPMI 1640 medium on ice until dissection is complete.
Macerate and press lymph nodes through a 40 μm mesh strainer into 3 mL of RPMI in a plastic Petri dish using the plunger of a 1 mL syringe. Rinse strainer with an additional 2 mL of RPMI and transfer the 5 mL cell suspension to a 15 mL conical tube. Rinse the Petri dish with an additional 5 mL of RPMI and transfer to the same tube. Centrifuge 5 min at 250 x g, discard supernatant, re-suspend the cell pellet in 1 mL FACS buffer (1X PBS, 1% bovine serum albumin and 10 mM EDTA) and transfer to a 1.5 mL Eppendorf tube.
3. Cell Staining and Flow Cytometry
The following is a typical staining protocol for identifying immune cell subsets using fluorescent antibodies against well-characterized immune cell surface markers. All antibodies were diluted into FACS buffer immediately before use.
Count cells and aliquot 106 cells into Eppendorf tubes. Centrifuge cells (see 2.7) and wash pellet 2X with 1 mL FACS buffer.
Incubate cells 20 min on ice in 100 μL of 2 ng/ mL rat anti-mouse CD16/32 to block endogenous Fc receptors. Pellet and wash cells with FACS buffer.
Add 100 μL of 1ng/ mL appropriate fluorescent antibod(y)ies to each sample and incubate on ice for one hour. Add 100 μL of FACS buffer alone to control cells. Pellet and wash cells twice with FACS buffer.
Re-suspend cells in 1 mL ACK buffer (150 mM NH4Cl, 1 mM KHCO3and 0.1 mM EDTA) Incubate on ice for 1 min to lyse red blood cells. Pellet and wash cells with FACS buffer. Re-suspend cells in 1 mL FACS buffer.
Collect data from cells using a flow cytometer capable of multicolor detection and analysis, typically with 405, 488 and 633 nm excitation lasers and five to ten fluorescence emission detectors.
4. Representative Results:
We purified prion rods from crude, prion-infected brain homogenate (Figure 1) Detergent solubilization and ultracentrifugation of the E2 elk brain homogenate (lanes 1 and 2) greatly enriched for prion rods (compare lanes 1 and 3), which was also protease K resistant (lane 4). Interestingly, undigested purified prion rods displayed a glycoform profile strikingly similar to digested E2 brain homogenate, indicating dramatic enrichment for prions. Identically-treated normal brain homogenate produced no PK-resistant PrPC (lanes 5 and 6).
Flow cytometric analysis of specific cell populations demonstrate that antigen presenting cells home to the peritoneum by two hours and retain fluorescent beads and prion rods, as evidenced by dramatically increased fluorescence over PBS injected controls (Figure 2). No cells harvested from the peritoneum of PBS-treated controls displayed fluorescence above background (Figs. 2A-G), while cells from bead (Figs. 2H-N) and prion (Figs. 2O-U)-inoculated mice displayed significant fluorescence, especially on monocytes (Figs. 2I and P), dendritic cells (Figs. 2K and R) and macrophages (Figs. 2L and S), and some Neutrophils (Figure 2J and Q) and fewer B cells (Figure 2T). Bead and Prion-bearing monocytes, dendritic cells and macrophages were also found in the mediastinal lymph nodes two hours after inoculation (Figs. 2AD and AK, AF and AM and AG and AN, respectively), while few or no PrPCWD-bearing Neutrophils, B or T cells were found there (Figs. 2AE and AL, AH and AG and AI and AP, respectively). We detected no beads or prion rods in the spleen or mesenteric lymph nodes (data not shown). In toto, these data demonstrate that immune cells traffic prions very similarly to other particulate antigens.
| REAGENT | FLUOROCHROME | EXCITATION LASER (nm) | PEAK EMISSION (nm) |
| Prion rods | DyLight 649 | 633 | 674 |
| 1 μm beads | AlexaFluor 660 | 633 | 685 |
| Antibodies | |||
| CD11b | eFluor 450 | 405 | 450 |
| Phycoerythrin-Cy7 | 488 | 760 | |
| CD11c | R-Phycoerythrin | 488 | 575 |
| Phycoerythrin-Cy7 | 488 | 760 | |
| Ly6C | Fluoroisothiocyante | 488 | 518 |
| Ly6G | R-Phycoerythrin | 488 | 575 |
| CD21 | DyLight 488 | 488 | 518 |
| B220 | Allophycocyanin-Cy7 | 633 | 785 |
| CD3 | R-Phycoerythrin | 488 | 575 |
Table 1. Spectral properties of fluorescent reagents
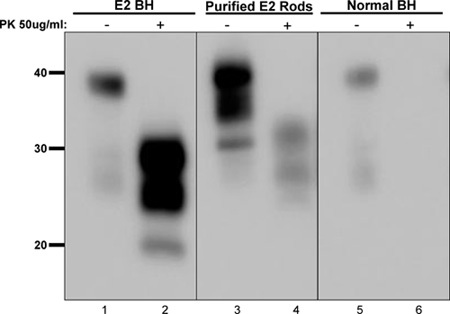 Figure 1. Purification of prion rods from infected brain homogenate. We used a 10% crude brain homogenate from a CWD-infected deer (E2, lanes 1and 2) as starting material from which we purified aggregated prion rods (lanes 3 and 4). Both crude and purified materials showed characteristic resistance to protease K treatment (lanes 1 - 4), whereas normal brain homogenate does not (lanes 5 and 6). Molecular weight markers are shown in Kd to the left of the blot. BH, brain homogenate.
Figure 1. Purification of prion rods from infected brain homogenate. We used a 10% crude brain homogenate from a CWD-infected deer (E2, lanes 1and 2) as starting material from which we purified aggregated prion rods (lanes 3 and 4). Both crude and purified materials showed characteristic resistance to protease K treatment (lanes 1 - 4), whereas normal brain homogenate does not (lanes 5 and 6). Molecular weight markers are shown in Kd to the left of the blot. BH, brain homogenate.
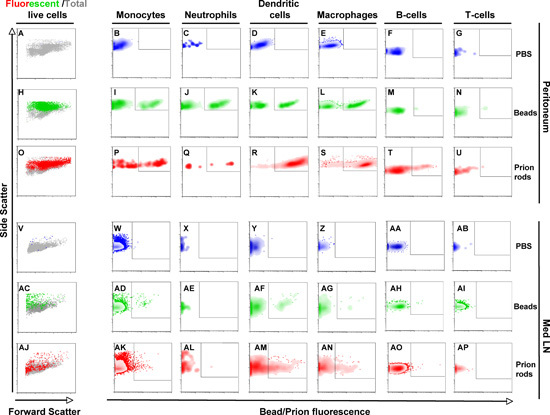 Figure 2. Flow cytometric analysis of immune cells trafficking prions from the peritoneum to mediastinal lymph nodes two hours after inoculation. PBS (panels A-G and V-AB), fluorescent beads (H-N and AC-AI) or Prion rods (O-U and AJ-AP) were injected into the peritoneum of mice and cells harvested from peritoneal lavage fluid (A-U) or mediastinal lymph nodes (V-AP) two hours later. Graphs in the first column show cells from mice treated with PBS (panels A and V), fluorescent beads (panel H and AC) and prion rods (panels O and AJ). Fluorescent cells (red or green dots) and total cells (grey dots) are plotted to show relative size (forward scatter), granularity (side scatter) and proportion of total live cells that fluoresce. Significant numbers of cells characteristic of granulocytes retained fluorescent beads and rods. Cells were also stained with antibodies against immune cell surface markers and gated for LyG- Ly6C+ monocytes (panels B, I, P, W, AD and AK), Ly6c- CD11c- CD11b+ Ly6G+ neutrophils (C, J, Q, X, AE and AL), Ly6G- Ly6C- CD11b+ CD11c+ dendritic cells (D, K, R, Y, AF and AM), Ly6G- Ly6C- CD11c- CD11b+ macrophages, (E, L, S, Z, AG and AN), CD3-B220+CD21+ B-cells (F, M, T, AA, AH and AO) and CD21-B220- CD3+ T cells (G, N, U, AB, AI and AP). Monocytes, dendritic cells and macrophages retained fluorescent beads and prion rods in the peritoneal cavity (panels I and P, K and R and L and S, respectively) and transported them to mediastinal lymph nodes (panels AD and AK, AF and AM and AG and AN, respectively), demonstrating similar uptake and trafficking of these two particulates. Neutrophils retained a significant amount of beads (J) and fewer rods (Q), but failed to deliver them to lymph nodes (AE and AL). B cells retained and transported some rods (T and AO) but virtually no beads (M and AH). T cells trafficked neither beads nor rods (N, U, AI and AP).
Figure 2. Flow cytometric analysis of immune cells trafficking prions from the peritoneum to mediastinal lymph nodes two hours after inoculation. PBS (panels A-G and V-AB), fluorescent beads (H-N and AC-AI) or Prion rods (O-U and AJ-AP) were injected into the peritoneum of mice and cells harvested from peritoneal lavage fluid (A-U) or mediastinal lymph nodes (V-AP) two hours later. Graphs in the first column show cells from mice treated with PBS (panels A and V), fluorescent beads (panel H and AC) and prion rods (panels O and AJ). Fluorescent cells (red or green dots) and total cells (grey dots) are plotted to show relative size (forward scatter), granularity (side scatter) and proportion of total live cells that fluoresce. Significant numbers of cells characteristic of granulocytes retained fluorescent beads and rods. Cells were also stained with antibodies against immune cell surface markers and gated for LyG- Ly6C+ monocytes (panels B, I, P, W, AD and AK), Ly6c- CD11c- CD11b+ Ly6G+ neutrophils (C, J, Q, X, AE and AL), Ly6G- Ly6C- CD11b+ CD11c+ dendritic cells (D, K, R, Y, AF and AM), Ly6G- Ly6C- CD11c- CD11b+ macrophages, (E, L, S, Z, AG and AN), CD3-B220+CD21+ B-cells (F, M, T, AA, AH and AO) and CD21-B220- CD3+ T cells (G, N, U, AB, AI and AP). Monocytes, dendritic cells and macrophages retained fluorescent beads and prion rods in the peritoneal cavity (panels I and P, K and R and L and S, respectively) and transported them to mediastinal lymph nodes (panels AD and AK, AF and AM and AG and AN, respectively), demonstrating similar uptake and trafficking of these two particulates. Neutrophils retained a significant amount of beads (J) and fewer rods (Q), but failed to deliver them to lymph nodes (AE and AL). B cells retained and transported some rods (T and AO) but virtually no beads (M and AH). T cells trafficked neither beads nor rods (N, U, AI and AP).
Discussion
Here we demonstrate a protocol for tagging and tracking prions in vivo that greatly facilitates monitoring early events in peripheral prion infection. This protocol greatly improves on past attempts at monitoring prion uptake in vitro9 and in vivo3,12 by pre-labeling highly enriched prion inoculum to unambiguously differentiate it from endogenous PrPC. Because infectious prions and PrPC share the same primary amino acid sequence, generating prion-specific antibodies has been problematic. We have performed similar experiments using PrP null mice injected with PK-digested and methanol precipitated crude brain homogenate that we tracked using Dylight 649-labelelled anti-PrP antibodies (our unpublished data). These experiments produced nearly identical results as shown here, but PrP null mice are not commercially available, not susceptible to prion disease 19 and lack the only bona fide prion receptor known, PrPC. This protocol allows the use of common laboratory mouse strains that express endogenous PrPC, obviating potential problems using PrP null mice.
The data shown here indicates that immune cells capture and traffic prions and fluorescent beads similarly, although B cells were identified as trafficking the former but not the latter. This corroborates previous studies implicating B cells as prion traffickers 6. However, our highly enriched prion preparation likely contains minute quantities of additional labeled proteins, so we cannot rule out the possibility that immune cells traffic these proteins too. Whether very small amounts of contaminating proteins alter prion trafficking or mimics the true biological scenario remains to be determined.
Successful monitoring of immune cells trafficking two different particulates further suggests that this protocol can be adapted to a wide variety of antigens that can be tagged and tracked, such as parasites, bacteria, viruses and proteins.
When performing ultracentrifugation steps in this protocol, large sample volumes can be aliquoted to smaller ones for convenient centrifugation as long as ratios remain constant. For example, instead of layering 100 mL of sample over 900 mL sucrose, one could layer 10 mL over 90 mL in separate tubes, then centrifuge.
Elevating the mouse posteriorly during inoculations moves peritoneal organs anteriorly, minimizing accidental needle injury to them. Slowly and carefully insert needles when inoculating mice and injecting and aspirating PBS from the peritoneum to avoid rupturing this delicate membrane.
When dissecting mice, take care to cut only though the skin in the initial incisions, leaving the thin membrane covering the peritoneum and mediastinum intact, to avoid cutting blood vessels and organs that may interfere with lymph node identification and retrieval.
Lymph nodes can be difficult to identify initially, often looking like adipose tissue. Lymph nodes appear nearly round, off-white to cream-colored and firmer than adipose tissue. These become much easier to identify with practice. Pool lymph nodes from at least two mice to recover enough cells for flow cytometric analyses.
For simultaneously identifying multiple cell surface markers, we strongly recommend using fluorescent antibodies over unlabeled antibodies that require a secondary fluorescent anti-isotype antibody for detection, as the latter experimental design requires careful selection of antibodies generated from multiple species and many more additional control samples.
Disclosures
No conflicts of interest declared.
Acknowledgments
We thank Steve McBryant and Jeff Hansen for help with ultracentrifugation and Patti Kiser for help with mouse handling. The National Institute of Neurological Diseases and Stroke at the National Institutes of Health, grant 5R01NS056379-02 funded this work.
References
- Klein MA. A crucial role for B cells in neuroinvasive scrapie. Nature. 1997;390:687–687. doi: 10.1038/37789. [DOI] [PubMed] [Google Scholar]
- Mabbott NA, Farquhar CF, Brown KL, Bruce ME. Involvement of the immune system in TSE pathogenesis. Immunol Today. 1998;19:201–201. doi: 10.1016/s0167-5699(98)01253-5. [DOI] [PubMed] [Google Scholar]
- Sigurdson CJ. PrP(CWD) lymphoid cell targets in early and advanced chronic wasting disease of mule deer. J Gen Virol. 2002;83:2617–2617. doi: 10.1099/0022-1317-83-10-2617. [DOI] [PubMed] [Google Scholar]
- Klein MA. Complement facilitates early prion pathogenesis. Nat Med. 2001;7:488–488. doi: 10.1038/86567. [DOI] [PubMed] [Google Scholar]
- Mabbott NA. Temporary depletion of complement component C3 or genetic deficiency of C1q significantly delays onset of scrapie. Nat Med. 2001;7:485–485. doi: 10.1038/86562. [DOI] [PubMed] [Google Scholar]
- Zabel MD. Stromal Complement Receptor CD21/35 Facilitates Lymphoid Prion Colonization and Pathogenesis. J Immunol. 2007;179:6144–6144. doi: 10.4049/jimmunol.179.9.6144. [DOI] [PubMed] [Google Scholar]
- Cordier-Dirikoc S, Chabry J. Temporary depletion of CD11c+ dendritic cells delays lymphoinvasion after intraperitonal scrapie infection. J Virol. 2008;82:8933–8933. doi: 10.1128/JVI.02440-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dorban G. Oral scrapie infection modifies the homeostasis of Peyer's patches' dendritic cells. Histochem Cell Biol. 2007;128:243–243. doi: 10.1007/s00418-007-0303-9. [DOI] [PubMed] [Google Scholar]
- Flores-Langarica A. Scrapie pathogenesis: the role of complement C1q in scrapie agent uptake by conventional dendritic cells. J Immunol. 2009;182:1305–1305. doi: 10.4049/jimmunol.182.3.1305. [DOI] [PubMed] [Google Scholar]
- Huang FP, MacPherson GG. Dendritic cells and oral transmission of prion diseases. Adv Drug Deliv Rev. 2004;56:901–901. doi: 10.1016/j.addr.2003.09.006. [DOI] [PubMed] [Google Scholar]
- Ano Y, Sakudo A, Nakayama H, Onodera T. Uptake and dynamics of infectious prion protein in the intestine. Protein Pept Lett. 2009;16:247–247. doi: 10.2174/092986609787601642. [DOI] [PubMed] [Google Scholar]
- Aucouturier P. Infected splenic dendritic cells are sufficient for prion transmission to the CNS in mouse scrapie. J Clin Invest. 2001;108:703–703. doi: 10.1172/JCI13155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang FP. Migrating intestinal dendritic cells transport PrP(Sc) from the gut. J Gen Virol. 2002;83:267–267. doi: 10.1099/0022-1317-83-1-267. [DOI] [PubMed] [Google Scholar]
- Jeffrey M. Transportation of prion protein across the intestinal mucosa of scrapiesusceptible and scrapie-resistant sheep. J Pathol. 2006;209:4–4. doi: 10.1002/path.1962. [DOI] [PubMed] [Google Scholar]
- Raymond CR, Mabbott NA. Assessing the involvement of migratory dendritic cells in the transfer of the scrapie agent from the immune to peripheral nervous systems. J Neuroimmunol. 2007;187:114–114. doi: 10.1016/j.jneuroim.2007.05.006. [DOI] [PubMed] [Google Scholar]
- Mohan J, Hopkins J, Mabbott NA. Skin-derived dendritic cells acquire and degrade the scrapie agent following in vitro exposure. Immunology. 2005;116:122–122. doi: 10.1111/j.1365-2567.2005.02207.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilch S. CpG and LPS can interfere negatively with prion clearance in macrophage and microglial cells. FEBS J. 2007;274:5834–5834. doi: 10.1111/j.1742-4658.2007.06105.x. [DOI] [PubMed] [Google Scholar]
- Safar J. Molecular mass, biochemical composition, and physicochemical behavior of the infectious form of the scrapie precursor protein monomer. Proc.Natl.Acad.Sci.U.S.A. 1990;87:6373–6373. doi: 10.1073/pnas.87.16.6373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Büeler HR. Mice devoid of PrP are resistant to scrapie. Cell. 1993;73:1339–1339. doi: 10.1016/0092-8674(93)90360-3. [DOI] [PubMed] [Google Scholar]