
Figure 5.
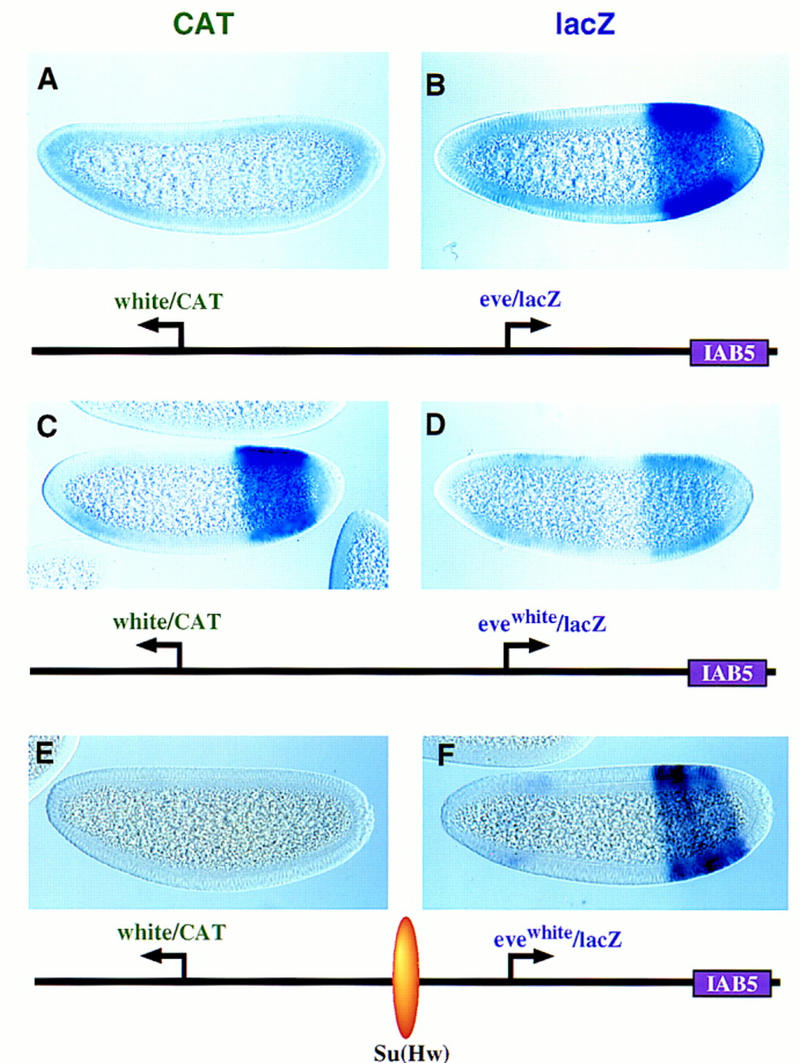
The IAB5 enhancer prefers TATA-containing promoters. Transgenic embryos were stained and oriented as described in the previous figure legends, except that these are younger embryos (between cellularization and the onset of gastrulation). The IAB5 enhancer was placed downstream of the rightward lacZ reporter gene. The distal CAT gene is under the control of the white promoter. The proximal lacZ gene is driven by eve (B) or an evewhite chimeric promoter (D,F) whereby the eve TATA region was replaced with the corresponding sequences in white. (A,B) CAT (A) and lacZ (B) staining patterns obtained with linked white/CAT and eve/lacZ genes. The IAB5 enhancer selects eve over white, so that the eve/lacZ reporter gene exhibits strong expression whereas white/CAT is silent. (C,D) Same as A and B except that the proximal lacZ gene is under the control of the evewhite chimeric promoter (D). There is only weak expression of the lacZ reporter in the presumptive abdomen (D). IAB5 now mediates strong expression of the distal white/CAT fusion gene (C). The residual lacZ staining observed in anterior regions (D) may be a position effect resulting from the site of P insertion. (E,F) Same as C and D except that the 340-bp gypsy insulator DNA [su(Hw)] was placed between the leftward CAT gene and rightward lacZ reporter. The insulator blocks IAB5–white interactions, so that CAT is not expressed above backround levels. Instead, IAB5 directs strong expression of the evewhite/lacZ in the presumptive abdomen, indicating that the chimeric promoter is not defective. The weak staining seen in head regions is caused by sequences contained within the P-transformation vector (Small et al. 1992).