
Figure 3. Circuit integrity and uniformity of the olivo-cortico-nucleo-olivary (OCNO) loop.
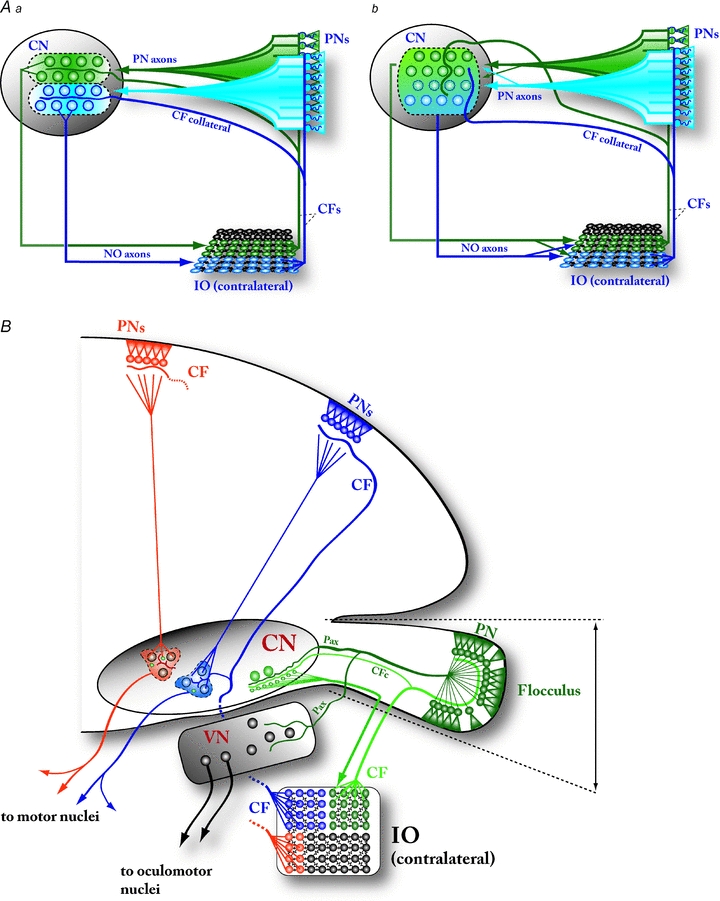
A, the concept of cerebellar microzones is partly built on the anatomical conservation of dedicated OCNO loops. Aa, according to the OCNO conservation model, single climbing fibres (CFs) diverge onto 5–10 PNs that are strictly localized in narrow parasagittal bands in the cerebellar cortex, and CFs originating from same inferior olivary (IO) areas target PNs in the same bands (‘blue’ IO neurons targeting PNs in the blue band, and ‘green’ IO neurons to the green band). This division into specialized bands is thought to extend into the CN by way of a precise topographic cortico-nuclear connection (the PN axons), that targets specific groups of nucleo-olivary (NO) neurons projecting to the IO region belonging to the same ‘loop’ (schematized as ‘blue’ or ‘green’ areas). This organization is further accentuated by collaterals of the CFs that are known to target NO neurons (and possibly other CN neurons) again within the same band. Strict conservation of OCNO bands allows the cerebellar cortex to respond to olivary signals (complex spikes) in each cortical zone independently. Ab, even though the strict zonal arrangement of CFs between IO and PNs, as well as a relatively clear topographical organization of cortico-nuclear PN axons, are well supported by anatomical evidence, the possibility of inter-zonal crosstalk at the level of CN is not excluded. First, the strict restriction of the olivo-nuclear projection via CF collaterals to CN neurons receiving input from PNs within the same parasagittal band has not been demonstrated, even though the majority of CF collaterals target CN neurons within the coarser anatomical groups (‘A–D2’, Voogd & Glickstein, 1998; ‘I–V’, Sugihara, 2010). Second, the role of the nucleo-olivary connection as a modulator of interneuronal coupling and synchronization among clusters of IO neurons suggests a more diffuse targeting. ‘Fuzzy’ OCNO loops allow activity in one cerebellar receptive field to influence IO-originating complex spikes (and the resulting effects on PN firing and PF–PN plasticity) across other, possibly not directly related cerebellar zones. B, schematic drawing of the cerebellar circuitry in a frontal view. The floccular cerebellum (FC; far right) and the related behavioural function (vestibulo-ocular reflex) have been the focus of intense study, and features described in this system (such as conservation of OCNO pathway on a single-cell level) have been thought to generalize to the entire cerebellum. However, the connections of the vestibulocerebellum (incorporating the floccular cerebellar cortex, the ventralmost part of the lateral CN, and specific regions of the contralateral IO, the ventrolateral olive and dorsal cap of Kooy) to the CN exhibit many structural differences compared to those from the cerebellar hemispheres. Most strikingly, the olivo-nuclear CF collaterals and PN axons in the FC target an area in the CN that has significantly fewer large glutamatergic projection neurons than the rest of the CN, and part of the FC signalling to its extracerebellar targets (the oculomotor nuclei) is conveyed via the projection neurons within the vestibular nuclei (VN) that are outside the OCNO loop. The areas in CN targeted by PNs from the cerebellar hemispheres have, on the other hand, fewer NO neurons compared to glutamatergic projection neurons. Whether the CF collaterals in these areas conform to the strict OCNO arrangement is not known. Abbreviations: CN, cerebellar nuclei; PNs, Purkinje neurons; CFs, climbing fibres; IO, inferior olive; NO, nucleo-olivary; CFc, climbing fibre collateral; Pax, Purkinje neuron axon; VN, vestibular nuclei.