

Abstract
Serum- and glucocorticoid-inducible kinase-1 (SGK1) is a serine/threonine protein kinase that responds to various stimuli and mediates cell survival. Although it is known that testicular torsion leads to testicular damage and male infertility, the role of SGK1 in torsion remains unclear. This study investigated whether torsion-induced apoptosis is associated with changes in phosphoinositide-dependent protein kinase-1 (PDK1), SGK1 and forkhead transcription factor FOXO3a expression and/or phosphorylation in rats. Sprague-Dawley rats were divided into four groups: sham (control), 1, 2 and 4 h of unilateral torsion. Bilateral testes, testicular interstitial fluid (TIF) and blood samples were collected immediately after torsion. Our results revealed that SGK1 protein and mRNA were abundantly present in testes and were induced by 2 h of torsion, but that phosphorylation of SGK1, PDK1 and FOXO3a decreased simultaneously. After 2 h of torsion, the testosterone secretion capacity of the primary Leydig cells and testicular interstitial cells (TICs) was impaired and apoptotic spermatogonia and TICs were observed; in addition, the mean seminiferous tubular diameter was decreased. Torsion increased plasma corticosterone levels, but decreased plasma luteinizing hormone and testosterone levels. However, the testosterone levels of the TIF in the ipsilateral testes were significantly enhanced after 2 h of torsion, but suppressed in the contralateral testes. This animal study suggests that PDK1, SGK1 and FOXO3a are involved in torsion-induced apoptosis and that medical therapy should be performed as early as 2 h after the occurrence of torsion to prevent further damage.
Keywords: apoptosis, FOXO3a, phosphoinositide-dependent protein kinase-1, serum- and glucocorticoid-inducible kinase-1, testicular torsion, testosterone
Introduction
Testicular torsion leads to infertility (Filho et al., 2004); it is caused by spermatic cord twisting (Turner & Brown, 1993; Turner et al., 2005). This obstruction of blood supply to the testes creates a condition in which there is a shortage of oxygen and nutrients from the blood and this results in testicular cell damage. Previous evidence from various ischaemia/reperfusion models has indicated that both necrosis and apoptosis are mechanisms for ischaemia-/reperfusion-induced cell death (Shiraishi et al., 2000, 2001). Specifically, necrosis rarely happens to testes examined immediately after torsion, but it appears late after prolonged ischaemia and ischaemia/reperfusion. A previous study indicated that testicular ischaemic injury produced by 1080° torsion did not lead to significant macroscopic or histological testicular necrosis after 1–4 h of torsion. Moreover, the testicular necrosis after 60 days detorsion was similar between groups with different periods of torsion (Romero et al., 2009). However, the detailed apoptotic mechanisms induced by up to 4 h testicular torsion remain to be determined.
The expression of serum- and glucocorticoid-inducible kinase-1 (SGK1), a serine threonine kinase, is mediated by a variety of extracellular stimuli (Richards, 2001); it controls cell volume, proliferation and apoptosis (Firestone et al., 2003; Lang et al., 2006). In cardiomyocytes, the inhibition of SGK1 activity can increase apoptosis from hypoxia or serum deprivation in vitro (Aoyama et al., 2005). SGK1 phosphorylation and activation exerts its anti-apoptotic activity through the inactivation of pro-apoptotic proteins such as forkhead transcription factor FOXO3a (also known as FKHRL1). FOXO3a has been shown to be negatively regulated by SGK1, the effect of which is to limit the arrest and apoptosis in the FOXO3a-dependent cell cycle (Brunet et al., 2001). Previous reports have shown that gene silencing with FOXO3a siRNA reverses oxygen-glucose deprivation-induced endothelial cell apoptosis and increases survival (Chong & Maiese, 2007), but that enhancing the phosphorylation of FOXO3a protects the mouse brain from ischaemic injury (Shioda et al., 2007). Moreover, to activate the SGK-mediated cell survival cascade, SGK1 binds to 3-phosphoinositide-dependent protein kinase-1 (PDK1) and induces phosphorylation at Thr256 by PDK1 (Kobayashi & Cohen, 1999; Park et al., 1999). This PDK1-dependent phosphorylation at SGK1 Thr256 is essential for the maximum stimulation of SGK1 activity (Biondi et al., 2001). However, the effects of ischaemia on the expression of PDK1, SGK1 and FOXO3a in vivo have not been studied. Hence, it would be a great interest to explore PDK1/SGK1/FOXO3a signal cascade in ischaemia-induced apoptosis.
Testicular torsion is a typical ischaemic model that directly leads to male infertility (Filho et al., 2004). The direct effects of testicular ischaemia on PDK1, SGK1 and FOXO3a have remained uninvestigated. Thus, we performed this animal study with the hypothesis that the PDK1/SGK1/FOXO3a signalling pathway is associated with testicular ischaemia-induced apoptosis in rats.
Materials and methods
Animals
Eight-week-old Sprague–Dawley male rats were maintained under controlled temperature (22 ± 2 °C) and light (06:00–20:00 hours) conditions, with food and water ad libitum. The use of the animals was approved by the Institutional Animal Care and Use Committee of National Yang-Ming University. All animals received human care in compliance with the Principles of Laboratory Animal Care and the Guide for Care and Use of Laboratory Animals (National Science Council, Taiwan).
Testicular torsion
Seventy-two male rats were divided randomly into four groups: sham (control group, n = 16), 1 h (n = 16), 2 h (n = 20) and 4 h (n = 20) of unilateral torsion. A left longitudinal scrotal incision was made under an intraperitoneal injection of pentobarbital (50 mg/mL/kg) to expose the left testis as described elsewhere (Yang et al., 2007). The left testis was twisted by turning it 720° counterclockwise, whereupon it was suture-fixed to the scrotal wall, to maintain the torsion state. Immediately after 1, 2 or 4 h of torsion, both the ipsilateral testis with torsion and the contralateral testis without torsion were removed and harvested for various measurements.
The functional capacity of the TICs and LCs to secrete testosterone
The preparation of testicular interstitial cells (TICs) and Leydig cells (LCs) has been described elsewhere (Huang et al., 2001). The preparations of TICs and LCs were found to contain approximately 20% and 85% Leydig cells (Lin et al., 1998), respectively. TICs (1 × 106 cells) and LCs (1 × 105 cells) were seeded into tubes, then challenged with human chorionic gonadotropin (hCG) (0 and 0.05 IU/mL) at 34 °C for 1 h. After being centrifuged at 50 g for 10 min, the supernatant was collected for testosterone radioimmunoassay (RIA).
Blood sampling and TIF collection
Blood samples were collected by heart puncture immediately after removal of the testes. The bilateral testes had their tunica albuginea pierced at the distal pole several times; then, the testicular interstitial fluid (TIF) was collected by centrifuging at 50 g for 15 min at 4 °C (Turner et al., 1984).
Histological examination
To determine morphological changes, the testes were fixed in Bouin's solution, dehydrated in an ethanol series and embedded in paraffin immediately after torsion. A number of 3-μm testes sections were prepared and then stained with haematoxylin and eosin (H&E). To measure the mean seminiferous tubular diameter (MSTD), each testis was assessed by measuring at least 25 separate tubular diameters, selecting the smallest and roundest seminiferous tubules in the field (Cosentino et al., 1986).
Measurement of apoptosis
Paraffin sections were prepared and then analysed using the DeadEnd Fluorometric TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick-end labelling) System (Promega, Madison, WI, USA). Immunofluorescence was observed using a Leica TCS SP2 confocal microscope (Leica Microsystems, Heidelberg, Germany). The average percentage of apoptotic tubules was estimated by examining 100 cross-sections of seminiferous tubules from each specimen. The seminiferous tubules that contained at least one TUNEL-stained nucleus were considered apoptotic (Yazawa et al., 2001).
Testosterone RIA
The concentrations of plasma and TIF testosterone were measured by RIA as described elsewhere (Wang et al., 1999; Chiao et al., 2002). The sensitivity of the testosterone RIA was 2 pg per tube. The intra- and inter-assay coefficients of variation were 4.1% (n = 6) and 4.7% (n = 10), respectively.
Corticosterone RIA
The concentration of plasma corticosterone was determined by RIA as described elsewhere (Pu et al., 2006; Chang et al., 2008). The sensitivity of corticosterone detection was 5 pg/mL. The intra- and inter-assay coefficients of variation were 3.4% (n = 5) and 9.5% (n = 5), respectively.
Determination of plasma LH levels
Plasma luteinizing hormone (LH) levels were analysed according to the manufacturer's protocol using the rat pituitary kit MILLIPLEX MAP by the Luminex xMAP technology (Millipore Corp., St Charles, MO, USA). The sensitivity of the bioassay was 4.9 pg/mL for LH. The intra- and inter-assay coefficients of variation were 15% (n = 12) and 9.5% (n = 5), respectively.
Immunoblotting
Testicular tissue suspensions were extracted with lysis buffer and disrupted by sonication. The protein extracts (20 μg) were separated by 8% sodium dodecyl sulphate-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride (PVDF) membrane. The following antibodies were used for immunoblotting: p-SGK1 Thr256, FOXO3a and SGK1 (Upstate Biotechnology, Lake Placid, NY, USA); p-SGK1 Ser422 (Santa Cruz Biotechnology, Santa Cruz, CA, USA); p-PDK1 Ser241, PDK1 and p-FOXO1 Thr24/FOXO3a Thr32 (Cell Signaling, Danvers, MA, USA); and β-actin (Chemicon, Temecula, CA, USA). An enhanced chemiluminescence kit (Amersham, Piscataway, NJ, USA) was used for immunodetection. The protein bands were quantified using the Image J software (NIH, Bethesda, MD, USA).
RNA extraction, RT-PCR and real-time PCR
Total RNA was extracted with Trlzol (Invitrogen, Carlsbad, CA, USA; Hsia et al., 2006). Reverse transcription (RT) and polymerase chain reactions (PCR) used SuperScript III Reverse Transcriptase (Invitrogen) and Taq DNA polymerase master mix (Ampliqon, Copenhagen, Denmark), respectively. The applied RT-PCR primers, the expected PCR product lengths and the thermal conditions are described elsewhere (Tsai et al., 2002). The hypoxanthine phosphoribosyltransferase gene (HPRT) was used as an internal control. The PCR products were size-fractionated on 2% agarose gels, and then stained with ethidium bromide before being photographed. The signals were measured using Image J software.
Real-time PCR was performed using the ABI PRISM 7700 sequence detection system with the TaqMan method (Applied Biosystems, Foster City, CA, USA). The sgk1 and HPRT primers and the fluorogenic probe (Hsu et al., 2009) were designed by Primer Express software and purchased from Applied Biosystems. The thermal conditions were 2 min at 50 °C and 10 min at 95 °C, followed by 40 cycles at 95 °C for 15 s and 60 °C for 1 min. The relative quantity of mRNA was estimated using a standard curve that was created by serial dilution of the RT product from control samples. The quantitative analysis of sgk1 gene expression was normalized to that of the HPRT gene expression.
Immunofluorescence microscopy
Testicular paraffin sections were prepared after torsion. A DNA marker and antibodies were used for immunostaining: 4,6-diamidino-2-phenylindole (DAPI), Rhodamine Red-X goat anti-rabbit immunoglobulin G (Jackson ImmunoResearch laboratories, Inc., West Grove, PA, USA), SGK1, FOXO3a and p-FOXO3a Thr32 (Abcam plc, Cambridge, UK). The sections were mounted using Vectashield mounting medium (Vector Laboratories, Burlingame, CA, USA). Immunofluorescence was observed using a Leica TCS SP2 confocal microscope (Leica Microsystems, Heidelberg, Germany).
Statistical analysis
Each result is represented as the mean ± SEM of at least three independent experiments. Data were processed by one-way analysis of variance (anova). Multiple comparisons were performed by Student–Newman–Keuls test whenever one-way anova was significant (p<0.05). SigmaStat (Systat Software Inc., Chicago, IL, USA) was used for data analysis.
Results
Torsion-induced pathological changes
The ipsilateral testes showed congestion and haemorrhage after 1 h of torsion, and this became more obvious after 2 and 4 h of torsion (Fig. 1A). Microscopic images of the architecture of ipsilateral testicular seminiferous tubules are shown in Fig. 1B: a–d. The interstitial spaces displayed oedema after 2 and 4 h of torsion. In addition, the MSTD of the ipsilateral testes at 1, 2 and 4 h of torsion were 5, 13.5 and 22.5% lower than that of the sham group, respectively. Compared with the contralateral testis, 2 and 4 h of torsion significantly reduced the MSTD of the ipsilateral testis. The MSTD of ipsilateral testis at 4 h was also less than that of the 1-h group (Fig. 1B: e).
Figure 1.
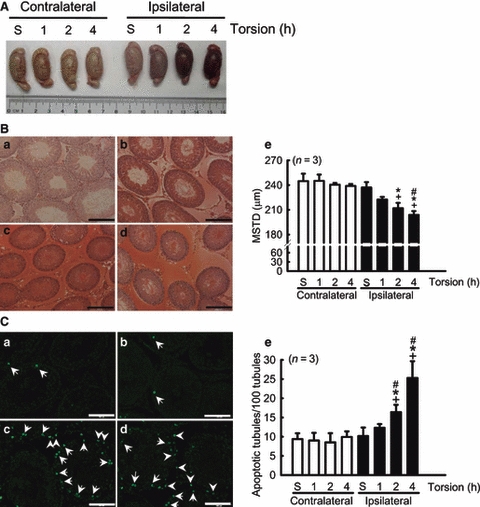
Apparent histological changes and apoptotic cells. (A) After 1 h of torsion, the ipsilateral testis showed congestion and haemorrhage and this was aggravated after 2 and 4 h of testicular ischaemia. (B) Oedema and haemorrhage were observed in the interstitial spaces, after (c) 2 h and (d) 4 h of torsion. Histological sections of the rats from the (a) sham and (b) 1-h groups show normal seminiferous tubular architecture (stain: haematoxylin and eosin; scale bar, 200 μm). (e) The quantifications of mean seminiferous tubular diameter of bilateral testes. (C) Testis sections stained by the terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick-end labelling (TUNEL) method demonstrate the presence of apoptotic cells in testes (a–d). Prominent apoptotic spermatogonia (a–d, arrows) and interstitial cells (c and d, arrow head) in the ipsilateral testes are indicated. (e) Quantitative comparisons of TUNEL-stained cells after different periods of torsion (scale bar, 100 μm). Values are mean ± SEM. *p<0.05, vs. respectively contralateral testes; +p<0.05, vs. sham (S) group; #p<0.05, vs. ipsilateral 1 h group.
Torsion-induced apoptosis
The percentage of apoptotic seminiferous tubules in the testes was detected by TUNEL assay (Fig. 1C: a–d) followed by a quantitative comparison (Fig. 1C: e). Torsion-induced apoptotic cells were most obvious in intra-tubular elements, but a few interstitial components were also observed. Interestingly, the apoptotic germ cells were found to be located at the periphery of each tubule (Fig. 1C: b–d), which are the spermatogonia. The percentages of TUNEL-positive seminiferous tubules of the ipsilateral sham, 1, 2 and 4 h groups were 10.2 ± 1.4, 12.3 ± 0.9, 16.4 ± 1.8 and 25.3 ± 4.4%, respectively. Compared with the contralateral testis in the 2 and 4 h groups, torsion significantly induced apoptosis in the ipsilateral testis. There were no statistically significant differences among the contralateral groups.
Torsion impaired the testosterone secretion capacity of TICs and LCs
The hCG-evoked testosterone secretion of the TICs and LCs was significantly reduced after 2 and 4 h torsion. In the LCs, there was approximately 29, 79 and 76% suppression of the hCG-evoked testosterone levels noted in the 1, 2 and 4 h groups as compared with the ipsilateral sham group. The hCG-evoked testosterone levels were 27.4-, 8.9-, 3.9- and 4.1-fold greater than the respective vehicle levels of the ipsilateral testes (Fig. 2A). In the TICs, the hCG-evoked testosterone levels of the 1, 2 and 4 h groups were approximately 32, 81 and 77% lower than the ipsilateral sham group. The hCG-evoked testosterone levels were 2.6-, 3.3-, 1.0- and 1.2-fold greater than the respective vehicle levels of the ipsilateral testes (Fig. 2B). However, the basal testosterone levels of either the LCs or the TICs were not significantly different across all groups.
Figure 2.
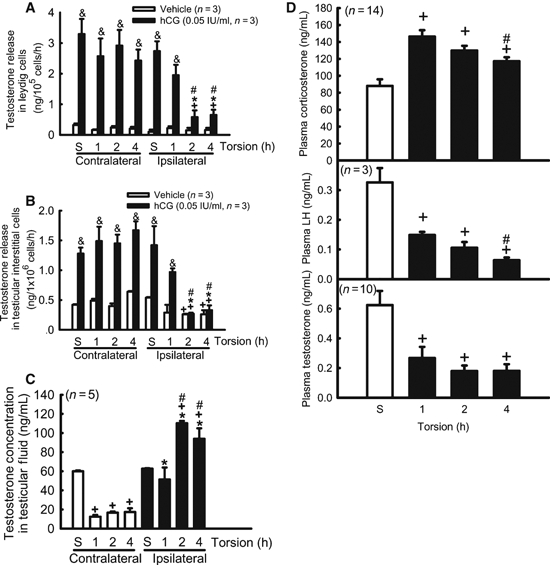
The effects of torsion on testosterone, corticosteroid and luteinizing hormone (LH) levels. Torsion impaired testosterone secretion capacity of (A) Leydig cells and (B) testicular interstitial cells. (C) The testosterone levels of testicular interstitial fluid were enhanced in the ipsilateral testes, but suppressed in the contralateral testes after unilateral torsion. (D) Experimental torsion increased plasma corticosterone levels and reduced plasma LH levels, which contributed to the decline in plasma testosterone levels. Values are mean ± SEM. *p<0.05, vs. respectively contralateral testes; +p<0.05, vs. sham (S) group; #p<0.05, vs. ipsilateral 1 h group; &p<0.05, vs. counterpart vehicle treatment.
Torsion altered the testosterone levels of the TIF
The TIF testosterone concentration was examined by RIA (Fig. 2C). The TIF testosterone levels of the ipsilateral testes were 1.7- and 1.5-fold increased after 2 and 4 h of torsion. The TIF testosterone levels of the contralateral testes at 1, 2 and 4 h of torsion were approximately 76, 68 and 67% lower than that in the sham group.
Estimation of hormone levels
Torsion increased the plasma corticosterone concentration. The corticosterone plasma levels after sham, 1, 2 and 4 h of torsion were 88 ± 7.8, 146.3 ± 7.6, 129.8 ± 5.6 and 117.2 ± 4.7 ng/mL, respectively (Fig. 2D). Torsion suppressed plasma LH and testosterone levels. The plasma LH concentrations after sham, 1, 2 and 4 h of torsion were 326 ± 48, 148 ± 10, 105 ± 19 and 63 ± 9 pg/mL, respectively. The plasma testosterone levels after sham, 1, 2 and 4 h of torsion were 0.63 ± 0.10, 0.26 ± 0.08, 0.18 ± 0.02 and 0.17 ± 0.03 ng/mL, respectively (Fig. 2D).
Torsion increased the mRNA and protein expression of SGK1
The expressions of sgk1 mRNA were determined by RT-PCR and real-time PCR. With RT-PCR using agarose gel quantitative comparison, the sgk1 mRNA expression of the 1, 2 and 4 h ipsilateral testes showed a 3.6–6.7-fold increase compared with the sham group (Fig. 3A,B). By real-tme PCR, the sgk1 mRNA expression of the 1, 2 and 4 h ipsilateral testes showed a 1.6–3.5-fold increase compared with the sham group (Fig. 3C). Across all the groups, there were no significant differences in the amount of sgk1 mRNA expression by the contralateral testes using either RT-PCR or real-time PCR.
Figure 3.
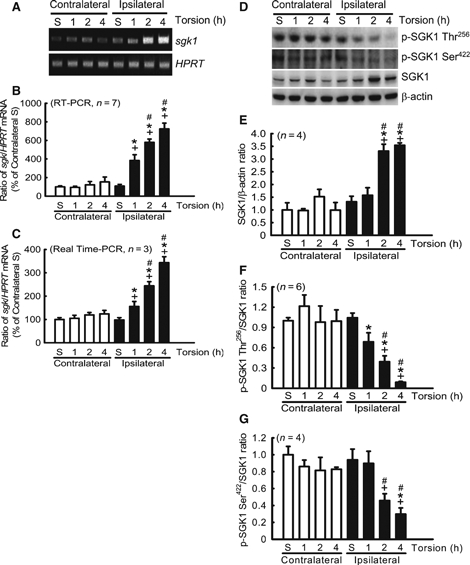
Torsion increased the expressions of serum- and glucocorticoid-inducible kinase-1 (SGK1) mRNA and protein, but suppressed SGK1 phosphorylation. Expression of sgk1 mRNA was increased by 2 and 4 h of torsion. (A) A representative gel pattern shows the expression of sgk1 mRNA for the bilateral testes after different periods of torsion. Quantitative comparisons of sgk1 mRNA were carried out by (B) reverse transcription-polymerase chain reaction (PCR) and (C) real-time PCR. Hypoxanthine phosphoribosyltransferase (HPRT) gene was used as an internal control. The values show the ratio of sgk1/HPRT as mean ± SEM. (D) A representative gel pattern shows expressions of SGK1, p-SGK1 Thr256 and p-SGK1 Ser422 in the bilateral testes following torsion. SGK1 protein expression was increased in the 2- and 4-h torsion groups compared with the sham group and in the ipsilateral testis (B), but expression of (C) p-SGK1 Thr256and (D) p-SGK1 Ser422 was decreased in the ipsilateral testes after torsion. Blotting shows SGK1 at 50 kDa, p-SGK1 Thr256 and Ser422 at 48 kDa and β-actin at 43 kDa. β-actin was used as an internal control. Values are mean ± SEM. *p<0.05, vs. respectively contralateral testes; +p<0.05, vs. sham (S) group; #p<0.05, vs. ipsilateral 1 h group.
A representative gel pattern showing the protein expression of SGK1 after torsion is shown in Fig. 3D. SGK1 protein expression was enhanced in the 2 and 4 h torsion groups, showing a 2.5-fold increase in the ipsilateral testes compared with the sham group (Fig. 3E). Thus, the expression of SGK1 at both the mRNA and protein levels is significantly increased after torsion.
Torsion decreased SGK1 phosphorylation at Thr256 and Ser422
The expressions of p-SGK1 Thr256 and Ser422 were investigated by immunoblotting. In the ipsilateral testes, the p-SGK1 Thr256 levels of the 1, 2 and 4 h torsion groups were approximately 34, 62 and 91% lower than those in the sham group (Fig. 3F), respectively. The p-SGK1 Ser422 levels of the ipsilateral testes in the 1, 2 and 4 h torsion groups were approximately 5, 69 and 75% lower than those in the sham group (Fig. 3G), respectively. Taken together, it was found that torsion suppressed the phosphorylation of SGK1.
Torsion decreased the phosphorylation of PDK1
A representative gel pattern displaying the expressions of PDK1 and the p-PDK1 Ser241 is shown in Fig. 4A. The ratios of p-PDK1 Ser241/PDK1 in the ipsilateral testes of the 1, 2 and 4 h groups were approximately 25, 41 and 58% lower than those in sham group, respectively. The p-PDK1 Ser241/PDK1 ratio of the ipsilateral testis was suppressed compared with the contralateral testis after 4 h of torsion (Fig. 4B). There were no differences among any of the groups in terms of the contralateral testes. Thus, torsion significantly decreased the phosphorylation of PDK1.
Figure 4.
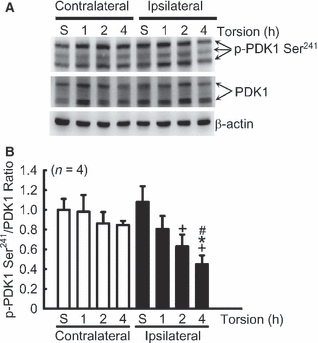
Torsion decreased the phosphorylation of phosphoinositide-dependent protein kinase-1 (PDK1). (A) A representative gel pattern shows the expression of p-PDK1 Ser241 and PDK1 after different periods of torsion. Blotting shows PDK1 (two bands) and p-PDK1 Ser241 (three bands) at 58–68 kDa, and β-actin at 43 kDa. β-actin was used as an internal control. (B) The ratio of p-PDK1 Ser241/PDK1 was decreased after 2 and 4 h of torsion in the ipsilateral testis. *p<0.05, vs. respectively contralateral testes; +p<0.05, vs. sham (S) group; #p<0.05, vs. ipsilateral 1 h group.
Torsion decreased the FOXO3a phosphorylation
Immunoblotting was performed for p-FOXO3a Thr32, the preferential phosphorylation site of SGK1 (Fig. 5A). The p-FOXO3a Thr32/FOXO3a ratio in the ipsilateral testes of the 1, 2 and 4 h groups was approximately 7, 42 and 74% lower than that in sham group, respectively. Thus, torsion suppressed the expression of p-FOXO3a Thr32 in the ipsilateral testis over a 2-h period compared with the contralateral testis (Fig. 5B).
Figure 5.

Torsion decreased the phosphorylation of FOXO3a. (A) A representative gel shows the expression of FOXO3a and p-FOXO3a Thr32 after torsion. Blotting shows FOXO3a at 90 kDa, p-FOXO3a Thr32 at 95 kDa and β-actin at 43 kDa. β-actin was used as an internal control. (B) The ratio of p-FOXO3a Thr32/FOXO3a in the ipsilateral testis was decreased after 2 and 4 h of torsion. *p<0.05, vs. respectively contralateral testes; +p<0.05, vs. sham (S) group; #p<0.05, vs. ipsilateral 1 h group.
Decrease in SGK1 and p-FOXO3a Thr32 expression in apoptotic cells
The colocalization of SGK1/TUNEL, p-FOXO3a Thr32/TUNEL and FOXO3a/TUNEL was detected by immunofluorescence microscopy in paraffin testicular sections (Fig. 6). Torsion-induced apoptotic cells can be identified by TUNEL stain in 4-h ipsilateral testis (Fig. 6A,E,I), whereas the expression of SGK1and p-FOXO3a Thr32 was barely detectable in the apoptotic cells (Fig. 6B,F). In addition, expression of FOXO3a (Fig. 6J) cannot be directly correlated with cells undergoing apoptosis.
Figure 6.
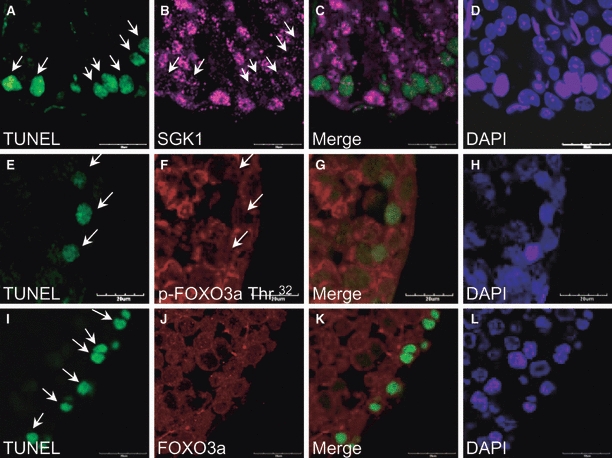
Immunofluorescent terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick-end labelling (TUNEL) staining of serum- and glucocorticoid-inducible kinase-1 (SGK1)/TUNEL, p-FOXO3a Thr32/TUNEL and FOXO3a/TUNEL in the ipsilateral 4-h testis sections. The apoptosis cells (arrows) are pinpointed by TUNEL stain (green; A, E, and I), whereas the expression of SGK1 (purple; B) and p-FOXO3a Thr32 (red; F) is barely detectable. The expression of FOXO3a (red; J) is not directly relatable to identifiable apoptotic cells. The colocalization images of SGK1/TUNEL, p-FOXO3a Thr32/TUNEL and FOXO3a/TUNEL are shown in C, G and K. The sections are counterstained with DAPI (blue; D, H and I). Scale bar: 20 μm.
Discussion
The results from this in vivo study support our hypothesis that testicular ischaemia-induced apoptosis is associated with the PDK1, SGK1 and FOXO3a signalling cascade. Our results suggest, after torsion stimulation, that the PDK1/SGK1/FOXO3a signalling cascade was downregulated, that the plasma corticosteroid level was increased, that plasma LH and testosterone levels were decreased and that the testosterone-secreting capacity of LCs and TICs was impaired.
The in vivo testicular ischaemic model in this study successfully led to testicular oedema, haemorrhage, congestion and cyanosis in the ipsilateral testis. Moreover, we noted that the MSTD was significantly decreased after 2 h of torsion, which is known to be an early sensitive indicator (Karaguzel et al., 1995; Zhang et al., 2002) of the testicular size, which has a positive correlation with testicular function in torsion (Takihara et al., 1987). It is well known that venous obstruction leads to hypoxia, poor nutrient supply and metabolic product accumulation in local testicular elements; the capillary permeability and hydrostatic pressure may be changed in testes (Altay et al., 2001). Therefore, torsion-induced interstitial oedema might alter the MSTD.
It is well known that the testicular torsion-induced damage severity depends on the duration and degree of the spermatic cord twisting. A previous study indicated that necrosis was only found in one rat immediately after 4 h of 1080° torsion (Romero et al., 2009). This group of rats underwent more severe torsion than the rats in our study group. Although we did not evaluate the extent of necrosis in the present experimental setting, it is reasonable to speculate that the extent of torsion-induced necrosis may well be less with 720° torsion than with 1080° torsion. Therefore, it is likely that the level of necrosis in our study will be small, if detectable at all.
In addition, torsion induced a progressive loss of testicular components, depending on cell type and ischaemic duration. We found that the TUNEL-positive staining mainly occurred in spermatogonia, although a few interstitial cells were also observed to be apoptotic after 2 h of torsion. This result is consistent with that of the previous reports in which LCs were less sensitive to ischaemic stimuli than spermatogenic cells (Anderson & Williamson, 1990; Arena et al., 2006). Turner et al. (2005) reported that torsion following by detorsion resulted in no LC apoptosis and that the twisted testis maintained intra-testicular testosterone levels with considerable steroidogenic capacity (Baker & Turner, 1995). However, our results revealed that hCG-induced testosterone secretion, one measure of the functional capacity of testosterone production, was significantly reduced in the LCs and TICs of the ipsilateral testes immediately after 2 h of torsion without detorsion, but that no effect on the basal testosterone secretion of LCs and TICs was detected between the bilateral testes. Although blood flow to the testis is occluded completely by the 720° spermatic cord twisting during the time of torsion (Turner, 1985), testosterone production was accompanied by basal testosterone release, which results in increased levels of TIF testosterone during mechanical stimuli in the ipsilateral testis following torsion.
In addition, an increase in plasma corticosterone level and a dramatic decrease in LH concentration were observed after torsion. This LH reduction might be caused by the stress-suppressed hypothalamus–pituitary–gonadal axis via increased Rfamide-related peptides and corticosterone production (Rivier & Rivest, 1991; Kirby et al., 2009) that inhibit secretion of gonadotropin-releasing hormone. It is reasonable to speculate that the reduction in LH may contribute to the decline to 50% of the sham group's contralateral TIF testosterone and plasma testosterone concentrations after 1 h of torsion.
Previous studied have revealed that SGK1 expression is altered by hypoxia/ischaemia stimulations. Aoyama et al. (2005) report that expressions of SGK1 and p-SGK1 Thr256 are elevated under hypoxia treatment, but lowered by serum deprivation in cardiomyocytes in vitro. We therefore speculated that SGK1 expression is altered by testicular ischaemia in vivo. However, previous reports have indicated that sgk1 gene expression in testicular tissue is very low and barely detectable when examined by Northern blotting (Webster et al., 1993; Waldegger et al., 1998). Using RT-PCR and real-time PCR, we found that sgk1 mRNA was abundantly present in rat testes. We also noted that the levels of SGK1 mRNA and protein were significantly increased after torsion. A previous study has revealed that androgen treatment increased SGK1 protein expression (Shanmugam et al., 2007). In this in vivo study, the expression of SGK1 in the ipsilateral testis is induced by torsion. This induction might relate to the increase in TIF testosterone levels after torsion, which has been described before. Our results provide new evidence for ischaemia-induced damage in vivo. In addition, previous studies have indicated that SGK1 expression can also be induced by ischaemia/reperfusion in vivo. Expression of the sgk1 gene is induced by ischaemia/reperfusion in brain (Lu et al., 2003; Nishida et al., 2004). Moreover, Rusai et al. (2009) indicated that SGK1 is upregulated after ischaemia and reaches its plateau after 6 h of reperfusion in kidney. In addition, it has been found that both phospho-SGK1 and total SGK1 increased 2–7 days after ischaemia/reperfusion in the cardiomyocytes (Aoyama et al., 2005).
In contrast, our results show that levels of p-SGK1 Thr256 and p-SGK1 Ser422 were suppressed after testicular ischaemia. Related to these differences in SGK1 and p-SGK1 levels after testicular ischaemia, it was noted that the expression of p-PDK1 Ser241 was decreased after 2 h of torsion. PDK1 is the kinase mainly responsible for SGK1 activation (Kobayashi & Cohen, 1999). Budas et al. (2006) found that inhibition of the ability to phosphorylate PDK1 results in a larger heart infarcted area after ischaemia/reperfusion. The mediation of cell survival by Ser241 phosphorylation needs the activation of PDK1 (Casamayor et al., 1999), and is altered under ischaemic stress affecting the kidney (Zheng et al., 2008) and the myocardium (Kis et al., 2003); this occurs via the regulation of phosphoinositide-3-kinase (PI3K) activity. Moreover, a previous report has revealed that testosterone suppresses PI3K activity (Papadopoulou et al., 2008), which results in a decrease in PDK1 phosphorylation. We therefore speculate that torsion-induced apoptosis may work together with PDK1 and SGK1 dephosphorylation in rats via increasing levels of TIF testosterone.
FOXO3a, a pro-apoptotic factor, is negatively regulated by SGK1. It has been shown that SGK1 phosphorylates the Thr32 of FOXO3a selectively (Brunet et al., 2001). When mutated at the SGK1 phosphorylation site, activation of FOXO3a leads to apoptosis, which is mediated by a decrease in SGK1 phosphorylation (Brunet et al., 2001). A recent report has shown that decreased phosphorylation of FOXO3a induces transcription of genes that promote cell-cycle arrest (p21 and p27) and apoptosis (Bim, FasL and TRAIL); this occurs by inhibiting ubiquitination, which inhibits degradation by the 26S proteasome (Yang et al., 2008). In this in vivo study, torsion increased SGK1 at both the mRNA and protein levels, but p-SGK1 Thr256 and p-SGK1 Ser422 were simultaneously reduced. This phenomenon may amplify the effects of SGK1 dephosphorylation and lead to FOXO3a dephosphorylation, then apoptosis. The expression of p-FOXO3a Thr32 was significantly decreased by 2 and 4 h of torsion; this is similar to the findings of a previous report, where chemical anoxic exposure inhibited p-FOXO3a Thr32 in LLC-PK1 cells (renal epithelial cell line from porcine kidneys) (Andreucci et al., 2003) in vitro. Moreover, using immunofluorescence microscopy, we found that apoptotic spermatogonia show a directly related decrease in expression of SGK1 and p-FOXO3a Thr32, but that there is no significant increase in the expression of FOXO3a. This decrease in p-FOXO3a Thr32 would seem to be related to the dephosphorylation of PDK1 and SGK1, clearly highlighting the importance of decreased phosphorylation of FOXO3a during torsion.
The PDK1, SGK1 and FOXO3a signalling cascade seems to be affected by various stimuli (Kobayashi & Cohen, 1999; Brunet et al., 2001; Leong et al., 2003), including ischaemic stimulation following apoptosis. Necrosis is no longer considered the sole mechanism for ischaemia-induced cell death, and it seems likely that apoptosis is an important mode of cell death, en route to ultimate cell death, in this situation. We therefore suggest that testicular ischaemia downregulates the phosphorylation of the PDK1/SGK1/FKHRL1 signalling pathway and this induces cell apoptosis, as shown in Fig. 7.
Figure 7.
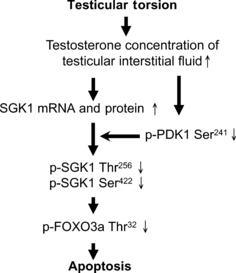
Schematic signalling model of the phosphoinositide-dependent protein kinase-1 (PDK1)/serum- and glucocorticoid-inducible kinase-1 (SGK1)/FOXO3a pathway during testicular torsion. We propose that testicular torsion decreases SGK1 phosphorylation via a decrease in PDK1 phosphorylation, which then appears to downregulate FOXO3a phosphorylation and this leads to cell apoptosis.
In a clinical context, the optimal time for torsion/detorsion surgery has been suggested to be 4–6 h after torsion has incurred. However, the results of this animal study suggest that torsion-induced spermatogonia apoptosis occurs at least partially via the PDK1/SGK1/FOXO3a signalling cascade and impairs the testosterone secretion capacity of the testes after only 2 h of torsion. In this context, we suggest that medical treatment, such as drug administration and reperfusion surgery, should be performed as early as 2 h after testicular ischaemia has occurred to prevent greater damage.
Acknowledgments
The authors appreciate Dr Yung-Chiong Chow (Department of Urology, Mackay Memorial Hospital) and Dr Chun-Jen Huang (Department of Anesthesiology, Mackay Memorial Hospital) for assistance and various suggestions with respect to the testicular animal model. They thank Dr Juliann G. Kiang (Armed Forces Radiobiology Research Institute, Uniformed Services University) for critical evaluation of the manuscript and helpful experimental suggestions. They also thank Dr Tzong-Shyuan Lee (Department and Institute of Physiology, National Yang-Ming University), Dr An-Hang Yang (Division of Ultrastructural and Molecular Pathology, Department of Pathology, Taipei Veterans General Hospital) and Dr Hwai-Shi Wang (Institute of Anatomy and Cell Biology, National Yang-Ming University) for their kind technical support and helpful discussions with respect to H&E staining. The authors acknowledge the immunofluorescence microscopy technical services provided by Imaging Core Facility of Nanotechnology of the University System of Taiwan & Yang-Ming University (UST-YMU). This research was supported by grants from the Veterans General Hospitals University System of Taiwan (VGHUST96-P7-32 and VGHUST99-P6-36).
References
- Altay B, Hekimgil M, KEFi A, Cikili N. Histopathological examination of both ipsilateral and contralateral testes with different obstructive models in prepubertal and adult rats. BJU Int. 2001;88:84–89. doi: 10.1046/j.1464-410x.2001.02256.x. [DOI] [PubMed] [Google Scholar]
- Anderson JB, Williamson RC. Fertility after torsion of the spermatic cord. Br J Urol. 1990;65:225–230. doi: 10.1111/j.1464-410x.1990.tb14715.x. [DOI] [PubMed] [Google Scholar]
- Andreucci M, Michael A, Kramers C, Park KM, Chen A, Matthaeus T, Alessandrini A, Haq S, Force T, Bonventre JV. Renal ischemia/reperfusion and ATP depletion/repletion in LLC-PK(1) cells result in phosphorylation of FKHR and FKHRL1. Kidney Int. 2003;64:1189–1198. doi: 10.1046/j.1523-1755.2003.00204.x. [DOI] [PubMed] [Google Scholar]
- Aoyama T, Matsui T, Novikov M, Park J, Hemmings B, Rosenzweig A. Serum and glucocorticoid-responsive kinase-1 regulates cardiomyocyte survival and hypertrophic response. Circulation. 2005;111:1652–1659. doi: 10.1161/01.CIR.0000160352.58142.06. [DOI] [PubMed] [Google Scholar]
- Arena F, Nicotina PA, Romeo C, Zimbaro G, Arena S, Zuccarello B, Romeo G. Prenatal testicular torsion: ultrasonographic features, management and histopathological findings. Int J Urol. 2006;13:135–141. doi: 10.1111/j.1442-2042.2006.01247.x. [DOI] [PubMed] [Google Scholar]
- Baker LA, Turner TT. Leydig cell function after experimental testicular torsion despite loss of spermatogenesis. J Androl. 1995;16:12–17. [PubMed] [Google Scholar]
- Biondi RM, Kieloch A, Currie RA, Deak M, Alessi DR. The PIF-binding pocket in PDK1 is essential for activation of S6K and SGK, but not PKB. EMBO J. 2001;20:4380–4390. doi: 10.1093/emboj/20.16.4380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brunet A, Park J, Tran H, Hu LS, Hemmings BA, Greenberg ME. Protein kinase SGK mediates survival signals by phosphorylating the forkhead transcription factor FKHRL1 (FOXO3a) Mol Cell Biol. 2001;21:952–965. doi: 10.1128/MCB.21.3.952-965.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Budas GR, Sukhodub A, Alessi DR, Jovanovic A. 3-phosphoinositide-dependent kinase-1 is essential for ischemic preconditioning of the myocardium. FASEB J. 2006;20:2556–2558. doi: 10.1096/fj.06-6252fje. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casamayor A, Morrice NA, Alessi DR. Phosphorylation of Ser-241 is essential for the activity of 3-phosphoinositide-dependent protein kinase-1: identification of five sites of phosphorylation in vivo. Biochem J. 1999;342(Pt 2):287–292. [PMC free article] [PubMed] [Google Scholar]
- Chang LL, Wun WS, Wang PS. Mechanisms of inhibition of dehydroepiandrosterone upon corticosterone release from rat zona fasciculata-reticularis cells. J Cell Biochem. 2008;104:359–368. doi: 10.1002/jcb.21627. [DOI] [PubMed] [Google Scholar]
- Chiao YC, Cho WL, Wang PS. Inhibition of testosterone production by propylthiouracil in rat Leydig cells. Biol Reprod. 2002;67:416–422. doi: 10.1095/biolreprod67.2.416. [DOI] [PubMed] [Google Scholar]
- Chong ZZ, Maiese K. Erythropoietin involves the phosphatidylinositol 3-kinase pathway, 14-3-3 protein and FOXO3a nuclear trafficking to preserve endothelial cell integrity. Br J Pharmacol. 2007;150:839–850. doi: 10.1038/sj.bjp.0707161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cosentino MJ, Sheinfeld J, Erturk E, Cockett AT. The effect of graded unilateral testicular biopsy on the reproductive capacity of male rats. J Urol. 1986;135:155–158. doi: 10.1016/s0022-5347(17)45552-2. [DOI] [PubMed] [Google Scholar]
- Filho DW, Torres MA, Bordin AL, Crezcynski-Pasa TB, Boveris A. Spermatic cord torsion, reactive oxygen and nitrogen species and ischemia-reperfusion injury. Mol Aspects Med. 2004;25:199–210. doi: 10.1016/j.mam.2004.02.020. [DOI] [PubMed] [Google Scholar]
- Firestone GL, Giampaolo JR, O'Keeffe BA. Stimulus-dependent regulation of serum and glucocorticoid inducible protein kinase (SGK) transcription, subcellular localization and enzymatic activity. Cell Physiol Biochem. 2003;13:1–12. doi: 10.1159/000070244. [DOI] [PubMed] [Google Scholar]
- Hsia SM, Chiang W, Kuo YH, Wang PS. Downregulation of progesterone biosynthesis in rat granulosa cells by adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) bran extracts. Int J Impot Res. 2006;18:264–274. doi: 10.1038/sj.ijir.3901405. [DOI] [PubMed] [Google Scholar]
- Hsu WL, Chiu TH, Tai DJ, Ma YL, Lee EH. A novel defense mechanism that is activated on amyloid-beta insult to mediate cell survival: role of SGK1-STAT1/STAT2 signaling. Cell Death Differ. 2009;16:1515–1529. doi: 10.1038/cdd.2009.91. [DOI] [PubMed] [Google Scholar]
- Huang WJ, Yeh JY, Kan SF, Chang LS, Wang PS. Effects of hyperprolactinemia on testosterone production in rat Leydig cells. J Cell Biochem. 2001;80:313–320. doi: 10.1002/1097-4644(20010301)80:3<313::aid-jcb30>3.0.co;2-e. [DOI] [PubMed] [Google Scholar]
- Karaguzel G, Gedikoglu G, Tanyel FC, Buyukpamukcu N, Hicsonmez A. Subsequent biological effects of chemical sympathectomy in rats undergoing unilateral testicular torsion. Eur Urol. 1995;28:147–151. doi: 10.1159/000475040. [DOI] [PubMed] [Google Scholar]
- Kirby ED, Geraghty AC, Ubuka T, Bentley GE, Kaufer D. Stress increases putative gonadotropin inhibitory hormone and decreases luteinizing hormone in male rats. Proc Natl Acad Sci USA. 2009;106:11324–11329. doi: 10.1073/pnas.0901176106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kis A, Yellon DM, Baxter GF. Second window of protection following myocardial preconditioning: an essential role for PI3 kinase and p70S6 kinase. J Mol Cell Cardiol. 2003;35:1063–1071. doi: 10.1016/s0022-2828(03)00208-6. [DOI] [PubMed] [Google Scholar]
- Kobayashi T, Cohen P. Activation of serum- and glucocorticoid-regulated protein kinase by agonists that activate phosphatidylinositide 3-kinase is mediated by 3-phosphoinositide-dependent protein kinase-1 (PDK1) and PDK2. Biochem J. 1999;339(Pt 2):319–328. [PMC free article] [PubMed] [Google Scholar]
- Lang F, Bohmer C, Palmada M, Seebohm G, Strutz-Seebohm N, Vallon V. (Patho)physiological significance of the serum- and glucocorticoid-inducible kinase isoforms. Physiol Rev. 2006;86:1151–1178. doi: 10.1152/physrev.00050.2005. [DOI] [PubMed] [Google Scholar]
- Leong ML, Maiyar AC, Kim B, O'Keeffe BA, Firestone GL. Expression of the serum- and glucocorticoid-inducible protein kinase, Sgk, is a cell survival response to multiple types of environmental stress stimuli in mammary epithelial cells. J Biol Chem. 2003;278:5871–5882. doi: 10.1074/jbc.M211649200. [DOI] [PubMed] [Google Scholar]
- Lin H, Wang SW, Tsai SC, Chen JJ, Chiao YC, Lu CC, Huang WJ, Wang GJ, Chen CF, Wang PS. Inhibitory effect of digoxin on testosterone secretion through mechanisms involving decreases of cyclic AMP production and cytochrome P450scc activity in rat testicular interstitial cells. Br J Pharmacol. 1998;125:1635–1640. doi: 10.1038/sj.bjp.0702229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu A, Tang Y, Ran R, Clark JF, Aronow BJ, Sharp FR. Genomics of the periinfarction cortex after focal cerebral ischemia. J Cereb Blood Flow Metab. 2003;23:786–810. doi: 10.1097/01.WCB.0000062340.80057.06. [DOI] [PubMed] [Google Scholar]
- Nishida Y, Nagata T, Takahashi Y, Sugahara-Kobayashi M, Murata A, Asai S. Alteration of serum/glucocorticoid regulated kinase-1 (sgk-1) gene expression in rat hippocampus after transient global ischemia. Brain Res Mol Brain Res. 2004;123:121–125. doi: 10.1016/j.molbrainres.2004.01.008. [DOI] [PubMed] [Google Scholar]
- Papadopoulou N, Charalampopoulos I, Alevizopoulos K, Gravanis A, Stournaras C. Rho/ROCK/actin signaling regulates membrane androgen receptor induced apoptosis in prostate cancer cells. Exp Cell Res. 2008;314:3162–3174. doi: 10.1016/j.yexcr.2008.07.012. [DOI] [PubMed] [Google Scholar]
- Park J, Leong ML, Buse P, Maiyar AC, Firestone GL, Hemmings BA. Serum and glucocorticoid-inducible kinase (SGK) is a target of the PI 3-kinase-stimulated signaling pathway. EMBO J. 1999;18:3024–3033. doi: 10.1093/emboj/18.11.3024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pu HF, Wang SW, Tseng CI, Huang HL, Lin CW, Hsu JM, Chen MJ, Chow YC, Wang PS. Mechanisms of digoxin and digitoxin on the production of corticosterone in zona fasciculata-reticularis cells of ovariectomized rats. J Cell Biochem. 2006;97:303–313. doi: 10.1002/jcb.20603. [DOI] [PubMed] [Google Scholar]
- Richards JS. New signaling pathways for hormones and cyclic adenosine 3′,5′-monophosphate action in endocrine cells. Mol Endocrinol. 2001;15:209–218. doi: 10.1210/mend.15.2.0606. [DOI] [PubMed] [Google Scholar]
- Rivier C, Rivest S. Effect of stress on the activity of the hypothalamic–pituitary–gonadal axis: peripheral and central mechanisms. Biol Reprod. 1991;45:523–532. doi: 10.1095/biolreprod45.4.523. [DOI] [PubMed] [Google Scholar]
- Romero FR, Gomes RP, Lorenzini F, Erdmann TR, Tambara FR. Ipsilateral testicular necrosis and atrophy after 1,080-degree torsion of the spermatic cord in rats. Acta Cir Bras. 2009;24:118–123. doi: 10.1590/s0102-86502009000200008. [DOI] [PubMed] [Google Scholar]
- Rusai K, Wagner B, Roos M, Schmaderer C, Strobl M, Boini KM, et al. The serum and glucocorticoid-regulated kinase 1 in hypoxic renal injury. Cell Physiol Biochem. 2009;24:577–584. doi: 10.1159/000257527. [DOI] [PubMed] [Google Scholar]
- Shanmugam I, Cheng G, Terranova PF, Thrasher JB, Thomas CP, Li B. Serum/glucocorticoid-induced protein kinase-1 facilitates androgen receptor-dependent cell survival. Cell Death Differ. 2007;14:2085–2094. doi: 10.1038/sj.cdd.4402227. [DOI] [PubMed] [Google Scholar]
- Shioda N, Ishigami T, Han F, Moriguchi S, Shibuya M, Iwabuchi Y, Fukunaga K. Activation of phosphatidylinositol 3-kinase/protein kinase B pathway by a vanadyl compound mediates its neuroprotective effect in mouse brain ischemia. Neuroscience. 2007;148:221–229. doi: 10.1016/j.neuroscience.2007.05.040. [DOI] [PubMed] [Google Scholar]
- Shiraishi K, Naito K, Yoshida K. Inhibition of calpain but not caspase protects the testis against injury after experimental testicular torsion of rat. Biol Reprod. 2000;63:1538–1548. doi: 10.1095/biolreprod63.5.1538. [DOI] [PubMed] [Google Scholar]
- Shiraishi K, Naito K, Yoshida K. Nitric oxide promotes germ cell necrosis in the delayed phase after experimental testicular torsion of rat. Biol Reprod. 2001;65:514–521. doi: 10.1095/biolreprod65.2.514. [DOI] [PubMed] [Google Scholar]
- Takihara H, Cosentino MJ, Sakatoku J, Cockett AT. Significance of testicular size measurement in andrology: II. Correlation of testicular size with testicular function. J Urol. 1987;137:416–419. doi: 10.1016/s0022-5347(17)44053-5. [DOI] [PubMed] [Google Scholar]
- Tsai KJ, Chen SK, Ma YL, Hsu WL, Lee EH. sgk, a primary glucocorticoid-induced gene, facilitates memory consolidation of spatial learning in rats. Proc Natl Acad Sci USA. 2002;99:3990–3995. doi: 10.1073/pnas.062405399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner TT. Acute experimental testicular torsion. No effect on the contralateral testis. J Androl. 1985;6:65–72. doi: 10.1002/j.1939-4640.1985.tb00817.x. [DOI] [PubMed] [Google Scholar]
- Turner TT, Brown KJ. Spermatic cord torsion: loss of spermatogenesis despite return of blood flow. Biol Reprod. 1993;49:401–407. doi: 10.1095/biolreprod49.2.401. [DOI] [PubMed] [Google Scholar]
- Turner TT, Jones CE, Howards SS, Ewing LL, Zegeye B, Gunsalus GL. On the androgen microenvironment of maturing spermatozoa. Endocrinology. 1984;115:1925–1932. doi: 10.1210/endo-115-5-1925. [DOI] [PubMed] [Google Scholar]
- Turner TT, Bang HJ, Lysiak JJ. Experimental testicular torsion: reperfusion blood flow and subsequent testicular venous plasma testosterone concentrations. Urology. 2005;65:390–394. doi: 10.1016/j.urology.2004.09.033. [DOI] [PubMed] [Google Scholar]
- Waldegger S, Barth P, Forrest JN, Jr, Greger R, Lang F. Cloning of sgk serine-threonine protein kinase from shark rectal gland – a gene induced by hypertonicity and secretagogues. Pflugers Arch. 1998;436:575–580. doi: 10.1007/s004240050674. [DOI] [PubMed] [Google Scholar]
- Wang SW, Lin H, Hwang JJ, Wang PS. Inhibition of testosterone secretion by digitoxin in rat testicular interstitial cells. J Cell Biochem. 1999;74:74–80. [PubMed] [Google Scholar]
- Webster MK, Goya L, Ge Y, Maiyar AC, Firestone GL. Characterization of sgk, a novel member of the serine/threonine protein kinase gene family which is transcriptionally induced by glucocorticoids and serum. Mol Cell Biol. 1993;13:2031–2040. doi: 10.1128/mcb.13.4.2031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang S, Shih HJ, Chow YC, Tsai PS, Wang TY, Wang PS, Huang CJ. The protective role of heme oxygenase-1 induction on testicular tissues after testicular torsion and detorsion. J Urol. 2007;177:1928–1933. doi: 10.1016/j.juro.2007.01.015. [DOI] [PubMed] [Google Scholar]
- Yang W, Dolloff NG, El-Deiry WS. ERK and MDM2 prey on FOXO3a. Nat Cell Biol. 2008;10:125–126. doi: 10.1038/ncb0208-125. [DOI] [PubMed] [Google Scholar]
- Yazawa H, Sasagawa I, Suzuki Y, Nakada T. Glucocorticoid hormone can suppress apoptosis of rat testicular germ cells induced by testicular ischemia. Fertil Steril. 2001;75:980–985. doi: 10.1016/s0015-0282(01)01705-8. [DOI] [PubMed] [Google Scholar]
- Zhang RD, Wen XH, Kong LS, Deng XZ, Peng B, Huang AP, Wan Y, Yang ZW. A quantitative (stereological) study of the effects of experimental unilateral cryptorchidism and subsequent orchiopexy on spermatogenesis in adult rabbit testis. Reproduction. 2002;124:95–105. doi: 10.1530/rep.0.1240095. [DOI] [PubMed] [Google Scholar]
- Zheng X, Xie L, Qin J, Shen H, Chen Z, Jin Y. Effects of wortmannin on phosphorylation of PDK1, GSK3-beta, PTEN and expression of Skp2 mRNA after ischemia/reperfusion injury in the mouse kidney. Int Urol Nephrol. 2008;40:185–192. doi: 10.1007/s11255-007-9215-9. [DOI] [PubMed] [Google Scholar]