
Figure 10.
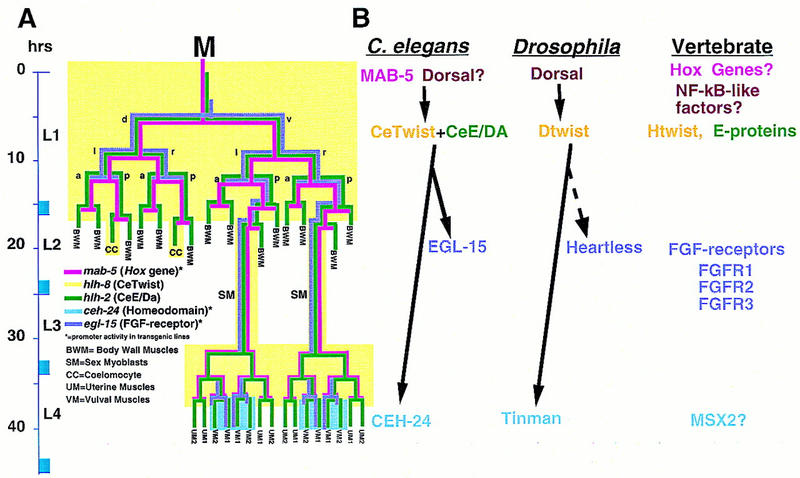
Aspects of a mesodermal patterning pathway are conserved. (A) A proposed pathway of genes active in the M lineage. Expression patterns for mab-5 and hlh-2 are derived from antibody staining (S. Salser and C. Kenyon, unpubl.; Krause et al. 1997, this work). mab-5 expression may extend to later stages in the SM lineages. Expression patterns for ceh-24 and egl-15 are predicted from reporter constructs (Harfe and Fire 1998; C. Branda and M. Stern, unpubl.). The hlh-8 pattern is inferred from both types of data (see text). The proposed pathway, inferred from genetic and interference experiments as described in the text, includes a requirement for MAB-5 in the early activation of hlh-8 and a role for the resulting CeTwist–CeE/DA heterodimer in the activation of ceh-24 and egl-15. Also as expected from this model (data not shown), MAB-5 was required for early (but not late) M lineage activity of egl-15 reporter constructs. (B) Comparative aspects of a Twist-dependent mesodermal diversification pathway. The vertical location of each protein name corresponds to the proposed time of action. In Drosophila, Dorsal can directly activate Dtwist by binding NF-κB/Dorsal sites upstream of Dtwist (Thisse et al. 1991). Dtwist can activate heartless (FGFR) expression (Shishido et al. 1997), although it is not known if this interaction is direct (dashed arrow). hlh-8 requires a consensus NF-κB/Dorsal site for promoter activity. No dorsal gene has been identified in C. elegans. Dominant mutations in human Twist (Htwist), FGFRs, and the homeodomain MSX2 can result in craniosynostosis (Jabs et al. 1993; Muenke et al. 1994; Meyers et al. 1995; Przylepa et al. 1996; Ghouzzi et al. 1997; Howard et al. 1997).