

Abstract
DNA was extracted from three fecal samples, more than 2,000 years old, from Hinds Cave, Texas. Amplification of human mtDNA sequences showed their affiliation with contemporary Native Americans, while sequences from pronghorn antelope, bighorn sheep, and cottontail rabbit allowed these animals to be identified as part of the diet of these individuals. Furthermore, amplification of chloroplast DNA sequences identified eight different plants as dietary elements. These archaic humans consumed 2–4 different animal species and 4–8 different plant species during a short time period. The success rate for retrieval of DNA from paleofeces is in strong contrast to that from skeletal remains where the success rate is generally low. Thus, human paleofecal remains represent a source of ancient DNA that significantly complements and may in some cases be superior to that from skeletal tissue.
DNA retrieved from excrement left by now extinct animals allows their identification and genetic study, and reveals aspects of their diet (1). Large amounts of ancient fecal material of putative human origin are also found during archaeological excavations, especially in dry caves and rock shelters. One such site is Hinds Cave, located on the eastern margin of the Chihuahuan Desert in southwestern Texas, where more than 1,000 putative human fecal deposits were found during an excavation in 1974 (2).
To investigate whether such material can be used to study DNA sequences from ancient humans and from the foods they ingested, we analyzed molecular composition and DNA preservation for three paleofecal samples (numbered I-III here) (Fig. 1). The results show that DNA sequences from the defecating individuals as well as those from plants and animals consumed by them can be retrieved. The analyses of these sequences allow the mtDNA population affiliations of the humans to be determined and also provides information about their diets.
Figure 1.
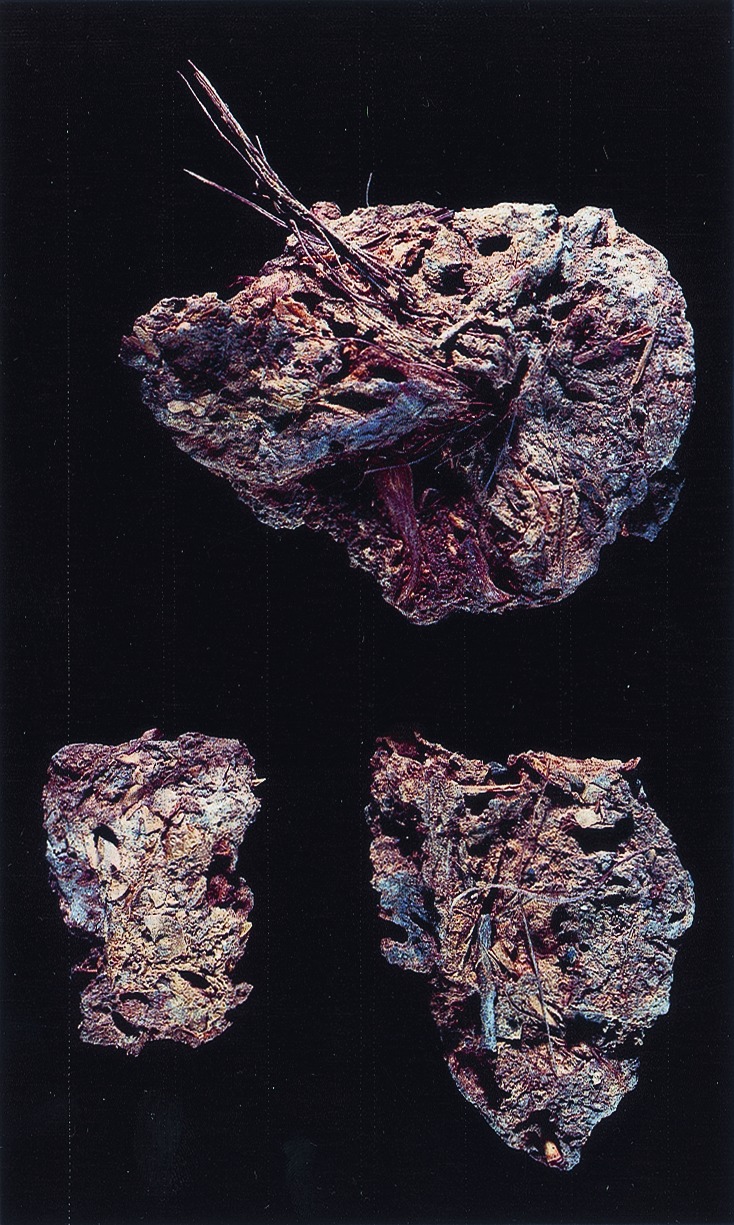
Human paleofeces from Hinds Cave, Texas. [Scale, 1 cm in the photo is equal to 2.78 cm real (slide).]
Materials and Methods
Experimental Procedures.
To assess the level of biomolecular preservation before DNA analysis, we analyzed the samples by pyrolysis gas chromatography/MS, as has been performed on previous coprolite samples (1). Approximately 15 mg of ground paleofecal samples was pyrolyzed and analyzed by pyrolysis-gas chromatography/MS) as described for bone samples (3).
DNA was extracted once from all samples as described (1). In addition, one extraction each from samples I and II was performed with the following modifications. Two to three grams of dried paleofecal matter was rehydrated for 5 days in the dark in a glass desiccator containing an open 500-ml bottle of double-distilled water, which was allowed to evaporate aided by a strip of Whatman filter paper with one end in the water and one end on the outside of the bottle. The relative humidity within the desiccator reached a plateau of 92% by the second day. After rehydration, samples were placed into 50-ml Falcon tubes containing 10 ml of lithium chloride extraction buffer (0.1 M Tris⋅Cl, pH 7.2/10 mM EDTA, pH 8.0/0.5 M LiCl/1% lithium dodecyl sulfate/50 mM DTT/200 μg/ml Proteinase K), which was rotated overnight at 37°C. Five milliliters of a 4% cetyltrimethylammonium bromide, 2% polyvinylpyrrolidone was added and again rotated overnight at 37°C. From this mixture, 1 ml was extracted as described (1) while the rest was frozen at −20°C for later extraction. The additional DNA extraction of sample III for independent replication was performed in Oxford as described (1).
PCR amplifications were performed as described (4) by using the primers listed below for three restriction sites (HaeIII, HincII, and AluI), the 9-bp repeat, hypervariable region I, 12S and 16S rRNA genes, and the chlorplast rbcL gene: L00635 5′-TGAAAATGTTTAGACGGCCTCACATC-3′; H00708, 5′-TAGAGGGTGAACTCACTGGAAC-3′; L13259, 5′-AATCGTAGCCTTCTCCACTTCA-3′; H13377, 5′-TATCTTGTTCATTGTTAACGTTGTGG-3′; L05054, 5′-TAGGATGAATAATAGCAGCTCTACCG-3′; H05184, 5′-GGGTGGATGGAATTAAGGGTGT-3′; L09158, 5′-ATACTACGGTCAATGCTCTG-3′; H09297, 5′-ATGCTAAGTTAGCTTTACAG-3′; L16131, 5′-CACCATGAATATTGTACGGT-3′; H16218, 5′-ATGTGTGATAGTTGAGGGTTG-3′; L16209, 5′-CCCCATGCTTACAAGCAAGT-3′; H16303, 5′-TGGCTTTATGTACTATGTAC-3′; L16287, 5′-CACTAGGATACCAACAAACC-3′; H16379, 5′-CAAGGGACCCCTATCTGAG-3′; 12Sa′, 5′-CTGGGATTAGATACCCCACTAT-3′; 12So, 5′-GTCGATTATAGGACAGGTTCCTCTA-3′; 16S6, 5′-TTTCGGTTGGGGCGACCTCGGAG-3′; 16S7, 5′-TTGCGCTGTTATCCCTAGGGTAACT-3′; rbcLZ1, 5′ATGTCACCACAAACAGAGACTAAAGCAAGT-3′; rbcL19b, 5′CTTCTTCAGGTGGAACTCCAG-3′, and rbcL19, 5′-AGATTCCGCAGCCACTGCAGCCCCTGCTTC-3′.
Amplifications of portions of the hypervariable region were performed twice for all samples to detect substitutions due to nucleotide misincorporations that can be present in all clones when amplifications that start from a few or from single template molecules (5). All amplification products were cloned into TA cloning vectors (Invitrogen) as per manufacturer's instructions as described (5). Colony PCR was performed as described (6). Sequencing was performed with a cycle sequencing kit (Amersham Pharmacia) as per manufacturer's instructions.
Plant Identification.
A total of 111 rbcL clones were sequenced. Because nucleotide differences present in only one clone are likely to be the result of nucleotide misincorporations during the PCR, the consensus sequences of each group of related sequences were taken to represent the sequence of a particular plant present in the paleofecal specimen. These sequences were taxonomically identified as described (1) by using the program blastn (January 20, 2000). Families and orders matching a 0 and/or 1 mismatch were noted. Where only one family matched the sequence, that family was assumed to be correct, where two or more families from the same order matched, the order was deemed the correct order. Five clones that could not be associated with any sequence cluster and matched database entries at more than two differences were deemed nonidentifiable. Finally, two identical clones from sample II (see Fig. 3, indicated by *) carried one difference to two families from the order Zingiberales. Members of this order are neither arid-adapted, nor adapted to continental climates, hence, the simplest explanation for this finding is a misidentification, probably because no representative of the correct family is present in the database.
Figure 3.

DNA sequences of clones from two fragments of the chloroplast rbcL gene amplified from the three paleofecal samples. Letters and numbers to the left indicate clone number, sample number [given as A(I), B(II), C(III)], extraction number, and PCR number. G indicates samples that were ground under liquid nitrogen and extracted, and R indicates samples that were rehydrated and extracted. X indicates clones that match data bank sequences with two or more differences. j indicates clones that represent putative jumping PCR events. Sequence clusters were identified as: A, Rhamnaceae; B, Ulmaceae; C, Fagaceae; D, Asteracea; E, Liliales; F, Fabaceae; G, Solonaceae; and H, Fouquieriaceae. * indicates two clones similar to the order Zingiberales.
Animal Identification.
To identify nonhuman vertebrate sequences in the 12S and 16S rRNA-encoding DNA (rDNA) amplicons, we cloned the PCR products and screened the clones via a colony PCR using the M13 forward and reverse primers with the addition of a third, human-specific primer (12SA′H 5′-GCCCTAAACCTCAACAGTTAAATC-3′ and 16S6H 5′-ACCAGTCAAAGCGAACTACTATAC-3′, respectively). All PCR products showing an amplification product of the expected length for a relevant insertion, but failing to show the shorter human amplification product were sequenced. The screening of 68 12S rDNA clones from sample I resulted in three nonhuman clones; whereas no nonhuman clones were found among 64 12S rDNA clones from sample II, nor among 33 and 28 16S rDNA clones from samples I and II, respectively. Sequences were compared with GenBank sequences by using blastn (January 20, 2000). Families matching at 0 and 1 mismatches were noted along with the next closest match, and identifications were made when one (and only one) family matched at 0 and 1 mismatch.
Results
Context and Dating.
Hinds Cave is a rock shelter located on the Pecos River of Val Verde County, Texas (2), which contains evidence of 10,000 years of intermittent occupation by prehistoric hunter-gatherers. The three paleofecal samples used in this study come from lens 13 of the undisturbed latrine block B. Lens 13 is 1.5–2 m below the current cave surface. This area consisted mainly of a large number of superimposed paleofecal deposits. While lithic debitage and stone tools were not common in area B, carbon dating of the lens places its deposition within period C, the Archaic. Nevertheless pieces of each sample were ground to a fine power and an aliquot of the powder from each sample was carbon-dated by accelerator MS to 2,165 ± 60 (Ua-15512), 2,370 ± 60 (Ua-15511), and 2,280 ± 90 (Ua-15386) years B.P.
Pyrolysis-Gas Chromatography/MS.
Other aliquots of the three samples were pyrolysed and the pyrolysis products were analyzed by gas chromatography/MS. All three samples revealed an abundance of polysaccharide derivatives and relatively unaltered lignin compounds, which indicate excellent preservation of plant tissues, whereas guaiacol and syringol derivatives indicate the presence of both di- and monocotyledonous plants (7). Furthermore, products derived from amino acids and peptides (diketopiperazines) as well as aliphatic compounds (fatty acids, alkenes, and steroids) suggest the presence of meat remains. Finally, abundant pyrroles, cyanobenzenes, and indoles, likely to derive from the Maillard reaction (8), i.e., the cross-linking of reducing sugars to primary amines (9), were seen in all samples. Because the resolution of Maillard products has been shown to make DNA from ancient remains available for enzymatic amplification (1), PTB (N-phenacyl thiazolium bromide), a chemical that breaks Maillard cross-links (10), was added to the DNA extractions.
mtDNA Analyses.
To determine whether the paleofeces were derived from humans, we performed four different amplifications of mtDNA fragments from the DNA extracts. These amplifications encompass three restriction sites and a 9-bp direct repeat of variable length, which together define the haplogroups A–D of phylogenetically related mtDNA types. These haplogroups together account for 95–100% of contemporary Native American mtDNA (11–13). The amplification products were cloned and multiple clones were sequenced (Fig. 2). In the case of sample I, all clones lacked a HaeIII site at position 663, carried a HincII site at position 13,259, an AluI site at position 5,176 as well as one copy of the 9-bp repeat (Table 1). This is indicative of haplogroup B. Because the assignment of haplogroups based on restriction sites correlates with DNA sequence motifs in the mtDNA control region (15), a portion of the control region was amplified to verify that the mtDNA was of Native American origin. In addition to substitutions in individual clones, probably PCR errors, all clones carried transitions at positions 16,189 and 16,217 and a transversion at position 16,183. The former two substitutions are typical of the Native American haplogroup B, whereas the latter substitution, when compared with 5,846 contemporary Hypervariable Region I (HVR I) sequences from all over the world (16), is seen in combination with the other two substitutions in only 16 individuals of Asian and American origin.
Figure 2.

DNA sequences of clones derived from sample I. (A) Clones from the three restriction fragments (brackets indicate restriction site) and 9-bp repeat, (B) a segment of hypervariable region I, (reference sequence from ref. 14), and (C) three clones from a fragment of the 12S rRNA gene matching to the genera Antilocapra and Sylvilagus, whose sequences appear as the reference sequence at top. Ambiguous bases are indicated by standard abbreviations and dashes indicate deletions.
Table 1.
Dates and mtDNA sequence data for three fecal samples
| Sample no. | 14C (yrs) | HG | Rest. sites
|
9-bp del | HVR I position*
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HaeIII 663 | HincII 13,259 | AluI 5,176 | 183 | 189 | 217 | 223 | 298 | 325 | 327 | 357 | ||||
| Ref. | − | + | + | − | A | T | T | C | T | T | C | T | ||
| I | 2,165 | B | − | + | + | + | C | C | C | na | na | na | na | na |
| II | 2,370 | C | − | − | + | − | na | na | T | T | C | C | T | C |
| III | 2,280 | C | − | − | + | − | na | na | T | T | C | C | T | T |
Carbon date, haplogroup designation (HG), restrictions site presence or absence, 9-bp deletion, and hypervariable region I (HVRI) positions are given for all three samples. na, not analyzed; ref, reference sequence (14).
Nucleotide positions are given after subtraction of 16,000.
Samples II and III were similarly analyzed. For sample II (see Fig. 4, which is published as supplemental data on the PNAS web site, www.pnas.org), both restriction sites and control region sequences unequivocally indicate that the DNA present belonged to Native American haplogroup C. For sample III (Fig. 5, which is published as supplemental data), the amplifications of the restriction sites revealed two different types of mtDNA in each of three amplifications. A majority of clones (9 of 10, 6 of 10, 5 of 7) suggested the presence of mtDNA belonging to haplogroup C, whereas the remaining clones carried another sequence that could stem from either haplogroup B or other contemporary DNA types. However, because two copies of the 9-bp repeat were present (data not shown), haplogroup B DNA is an unlikely source for the second DNA sequence. The analysis of the control region similarly indicated that the DNA of at least two individuals was present, one belonging to haplogroup C (5 of 9 clones) and the other representing a mtDNA type not seen in contemporary Native Americans, however, common among Europeans and therefore a likely contaminant. To investigate whether the results obtained were reproducible, and to clarify which of the two DNA types present in the extract from sample III were endogenous, a sample was sent to Oxford, where an independent extraction and amplification of the mtDNA control region was performed. Upon direct sequencing of this amplification product, only one sequence, carrying three substitutions assigning it to haplogroup C, was detected (Fig. 5). Sequencing of 10 clones of the amplification product confirmed this result. Thus, this sequence was considered endogenous to sample III. It is furthermore noteworthy that the mtDNA sequence determined from this sample differs at one nucleotide position (C at 16357) from the sample II sequence. Thus, while all paleofecal samples stem from the same approximate time period, they derive from three different individuals.
Plant Diet.
To analyze the plant diet of these individuals, we amplified a 157-bp fragment (including primers) of the chloroplast rbcL gene (1) from all samples, and, in addition, a 183-bp fragment (including primers) from samples I and II. The products were cloned, and clones were sequenced until the same groups of related DNA sequences were repeatedly found (Fig. 3). The consensus sequences of each group were compared with the approximately 4,000 rbcL sequences present in GenBank. This allowed the taxonomic identification at the order, and in some cases the family level (1). Sample I contained DNA sequences from Liliales (an order including locally common plants such as Agave and Yucca), Asteraceae (the sunflower family, also common locally), and Ulmaceae (the elm family, in this case, probably the hackberry Celtis). Sample II contained DNA from Liliales, Asteraceae, Fagaceae (the oak family, present in the canyons nearby and possibly ingested in the form of acorns), Solanaceae (the nightshade family, probably the locally present, edible Physalis), and Ulmaceae. Sample III contained DNA from Asteraceae, Fabaceae (Legumes), Fouquieriaceae (the ocotillo family, present locally), Rhamnaceae (the buckthorn family, probably the locally common shrub Condalia), and Ulmaceae (Table 2).
Table 2.
Plant and animal remains identified from the paleofecal samples
| Plant order/family genera (common name) | Molecular
|
Macro
|
Animal family/genera (common name) | Molecular
|
Macro
|
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | I | II | III | I | II | III | I | II | III | ||
| Asterales/asteraceae (sunflower) Helianthus, many genera | 7 | 3 | 4 | Antilocapridae Antilocapra (pronghorn) | 2 | ||||||||
| Caryophyllales/cactaceae (cacti) Opuntia | √b | √ | Bovidae (sheep)gOvis | 11 | |||||||||
| Ericales/fouquieriaceae (ocotillo) Foquieria | 1 | Cricetidaee (packrats, cotton rats) Sigmodon, Neotoma | √ | √ | √ | ||||||||
| Fabales/fabaceae (legumes) Acacia, Prosopis, Sophora, Mimosa | 7 | √c | Leporidae (cottontail)hSylvilagus | 1 | |||||||||
| Fagales/fagaceae (oak) Quercus | 7 | 1 | Osteichthyes (bony fish) | √ | |||||||||
| Liliales/lilliaceae (lily) Yucca, Allium, Dasylirion, Nolina | 10 | 4 | √a | √ | √ | Sciuridae (squirrels)fCitellus | √ | ||||||
| Rhamnales/rhamnaceae (buckthorn) Karwinskia, Condalia, Colubrina | 10 | ||||||||||||
| Rosales/ulmaceae (elm) Celtis | 8 | 12 | 5 | √d | |||||||||
| Solonales/solanaceae (nightshade) Nicotiana, Physalis, Lycium, Datura | 6 | ||||||||||||
Plant (families/orders) and animal (families/genera) remains found by molecular and microscopic analysis for the three paleofecal samples (I, II, III). The assignment of possible plant genera is based on contemporary flora of the lower Pecos region, Texas, and comparison with previous work (17). The number of clones assigned to each taxon are given. Taxa identified morphologically were:
Allium sp., Agave sp., and Yucca sp.;
Opuntia sp.;
Prosopis sp.;
Celtis sp.;
Neotoma and Sigmodon sp.;
Citellus sp.;
not found near the site at present, but previously present in the Guadalupe and Chisos mountains;
no macroremains in these paleofecal specimens, although commonly found in other Hinds Cave paleofeces (17).
The three paleofecal samples also were analyzed microscopically. This confirmed the presence of Lilliceae (lillies), Fabaceae, and Ulmaceae. The other six plants identified from the DNA sequences were not found, but Cactaceae (cactus family, common locally), not identified among the clones sequenced, was observed microscopically.
Animal Diet.
To analyze the meat diet of the three individuals, we amplified a 151-bp fragment of the mitochondrial 12S rDNA gene and a 141-bp fragment (including primers) of the mitochondrial 16S rDNA gene from each sample. In all cases, amplification products were obtained and cloned, and about 10 clones were sequenced. Not surprisingly, the vast majority of the clones carried human sequences, presumably derived from the defecating individuals. Thus, from a total of 40 clones sequenced from samples I and II, all were of human origin. For sample III, 6 of 8 12S rDNA sequences, and 5 of 14 16S rDNA sequences were of nonhuman origin and identical to each other. When the 12S rDNA sequence was compared with the approximately 2,000 vertebrate 12S rDNA sequences present in GenBank, it was found to be identical to one species in the genus Ovis (O. aries, sheep), while the next closest sequences carried three differences and belonged to members of the genera Capra (goats) and Cephalophus (African duikers). When the 16S rDNA sequence was similarly compared with 400 GenBank sequences, it was found to be identical to two members of the genus Ovis (O. dalli, O. aries), to differ at one position from bighorn sheep (O. canadensis), and at five positions from the next closest genus, Oreamus (American mountain goats). Although there is no archeological or paleontological record of big horn sheep in the Lower Pecos region, bighorn sheep (O. canadensis) have historically inhabited the desert mountain ranges of western Texas (18, 19). Although it is possible that domestic sheep (O. aries) could have contaminated this sample after their introduction into North America, we find this unlikely because the samples stem from 2 m below the current surface of the cave. Further sequences would have to be obtained to show this unambiguously, however.
To determine whether a small proportion of the molecules amplified from samples I and II might derive from food remnants, we screened the clones from the rDNA amplification products for sequences of nonhuman origin. No such clones were found in the case of sample II, but three nonhuman 12S rDNA sequences were found for sample I. Two of these are identical to pronghorn antelope (Antilocapra americana), while the next closest sequence differed at seven positions from two members of the family Bovidae. The third sequence is identical to cottontail rabbit (Sylvilagus audobonii), while showing nine differences to the next closest match in the family Muridae. Both pronghorn antelope and cottontail rabbit are known to have been present in the area, and a tooth tentatively identified as pronghorn antelope (20), as well as cottontail remains (21), have been found in the cave. Hence, the sequences were identified as stemming from pronghorn antelope and cottontail rabbit, respectively.
In contrast to the molecular analysis, microscopic examination identified only small mammals and fish in the samples (Table 2). A total of three teeth and one bone of packrat (Neotoma sp.) were found in the three samples, while sample I also contained a single squirrel bone, sample II five scales and a spine from a bony fish, and sample III two teeth and a femur of a cotton rat (Sigmodon sp.).
Discussion
Usually, microscopic analysis are used to investigate the animal and plant diet from paleofeces. For animals, this approach relies on the presence of tissues such as hair, fur, bone, and scales that pass the intestinal tract relatively intact. In contrast, molecular analyses can identify pure meat. Thus, the failure of earlier analyses to identify big game as a part of the diet of these individuals is easily explainable. That the molecular analysis failed to identify small mammals and fish may not be surprising either, in view of the fact that only a few remains of these species were observed, and they may have been comparatively minor components in the diets of these individuals over the time period covered by the samples. In view of these findings, it is tempting to suggest that emphasis on small mammals in hunter-gatherer diets (e.g., ref. 22) sometimes may be mistaken because big game would tend to be ingested as pure meat and not together with more resistant tissues that can be morphologically identified in feces. It also may have been the case that large animals were butchered at the kill site and only the meat carried back to the occupation site, hence only a few remains of such large animals would be found at excavations of rock shelters like Hinds Cave (20).
The diversity of both meat and plants ingested by the Archaic inhabitants of Hinds Cave is noteworthy. Sample I contained evidence of four animals (pronghorn antelope, cottontail rabbit, packrat, and squirrel) and four plants (hackberry, sunflower family, yucca or agave, and opuntia), sample II contained two animals (packrats and fish) and six plants (hackberry, oak, sunflower family, yucca or agave, nightshade family, and legume family), and sample III contained three animals (bighorn sheep, packrat, and cotton rat) and eight different plants (Buckthorn family, hackberry, oak, sunflower family, yucca or agave, legume family, ocotillo, and opuntia). This represents a remarkably rich diet. Other evidence also indicates that the diet of these prehistoric hunter-gatherers included a considerable diversity of plants and animals (23). Thus, as compared with individuals dependent on agriculture, the diet of the Hinds Cave hunter-gatherers seems to have been more varied and nutritionally sound (17).
Conclusions
Human mtDNA sequences affiliated with current Native American groups could be retrieved from all three fecal samples analyzed, a similarly high success rate has been achieved in studies of fecal matter from now extinct animals (24). This is in marked contrast to most skeletal and mummified human tissue where the success rate of DNA retrieval is low (25). Although skeletal remains obviously have advantages, for example, for the study of social contexts revealed by burial practices, these findings show that paleofeces from dry cave and rock shelter sites represent a source of ancient DNA that is relatively abundant, and from which the DNA is more reliably retrievable than is the case for human skeletal remains. In addition, paleofecal DNA offers valuable information on the meat and plant components of ancient diets that complements and extends information obtainable by morphological and biochemical (26) analyses. Finally, paleofeces may be more accessible to scientific study than other remains, because they do not represent objects of spiritual value (27).
Supplementary Material
Acknowledgments
We thank D. Poinar for help and constant support; M. Hofreiter, P. Martin, S. Meyer, I. Nasidze, G. Poinar, D. Serre, M. Stoneking, and L. Vigilant for constructive discussions; G. Possnert for carbon dating; S. Bicknell for photographs, and the Max Planck Society and the Deutsche Forschungsgemeinschaft for financial support.
Abbreviation
- rDNA
rRNA-encoding DNA
Footnotes
References
- 1.Poinar H N, Hofreiter M, Spaulding G S, Martin P S, Stankiewicz A B, Bland H, Evershed R P, Possnert G, Pääbo S. Science. 1998;281:402–406. doi: 10.1126/science.281.5375.402. [DOI] [PubMed] [Google Scholar]
- 2.Shafer H J, Bryant V M. Archeological and Botanical Studies at Hinds Cave, Val Verde County, Texas. College Station: Texas A&M University Anthropology Laboratory; 1977. [Google Scholar]
- 3.Poinar H N, Stankiewicz B A. Proc Natl Acad Sci USA. 1999;96:8426–8431. doi: 10.1073/pnas.96.15.8426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Höss M, Pääbo S. Nucleic Acids Res. 1993;21:3913–3914. doi: 10.1093/nar/21.16.3913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Krings M, Stone A, Schmitz R W, Krainitzki H, Stoneking M, Pääbo S. Cell. 1997;90:19–30. doi: 10.1016/s0092-8674(00)80310-4. [DOI] [PubMed] [Google Scholar]
- 6.Kilger C, Krings M, Poinar H N, Pääbo S. BioTechniques. 1997;22:412–414. doi: 10.2144/97223bm08. [DOI] [PubMed] [Google Scholar]
- 7.Van Bergen P F, Goni M, Collinson M, Boon J J, Goth K, de Leeuw J W. Geochim Cosmochim Acta. 1994;58:3823–3844. [Google Scholar]
- 8.Yaylayan V A, Keyhani A, Huygues-Despointes A. Adv Exp Med Biol. 1998;443:237–244. doi: 10.1007/978-1-4899-1925-0_19. [DOI] [PubMed] [Google Scholar]
- 9.Mauron J. Prog Food Nutr Sci. 1981;5:5–35. [PubMed] [Google Scholar]
- 10.Vasan S, Zhang X, Kapurmitou A, Bernhagen J, Teichberg S, Basgen J, Wagle D, Shih D, Terlecky I, Bucala R, et al. Nature (London) 1996;382:275–278. doi: 10.1038/382275a0. [DOI] [PubMed] [Google Scholar]
- 11.Smith D G, Malhi R S, Eshleman J, Lorenz J G, Kaestle F A. Am J Phys Anthropol. 1999;110:271–284. doi: 10.1002/(SICI)1096-8644(199911)110:3<271::AID-AJPA2>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- 12.Schurr T G, Ballinger S W, Gan Y Y, Hodge J A, Merriwether D A, Lawrence D N, Knowler W C, Weiss K M, Wallace D C. Am J Hum Genet. 1990;46:613–623. [PMC free article] [PubMed] [Google Scholar]
- 13.Brown M D, Hosseini S H, Torroni A, Bandelt H J, Allen J C, Schurr T G, Scozzari R, Cruciani F, Wallace D C. Am J Hum Genet. 1998;63:1852–1861. doi: 10.1086/302155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Anderson S, Bankier A T, Barrell B G, de Bruijn M H L, Coulson A R, Drouin J, Eperon I C, Nierlich D P, Roe B A, Sanger F, et al. Nature (London) 1981;290:457–465. doi: 10.1038/290457a0. [DOI] [PubMed] [Google Scholar]
- 15.Ward R H, Frazier B L, Dew-Jager K, Pääbo S. Proc Natl Acad Sci USA. 1991;88:8720–8724. doi: 10.1073/pnas.88.19.8720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Burckhardt F, von Haeseler A, Meyer S. Nucleic Acids Res. 1998;27:138–142. doi: 10.1093/nar/27.1.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sobolik K D. Prehistoric Diet and Subsistence in the Lower Pecos as Reflected in Coprolites from Baker Cave, Val Verde County, Texas. University of Texas, Austin: Texas Archeological Research Laboratory; 1991. [Google Scholar]
- 18.Davis W B. The Mammals of Texas. Texas A&M University, College Station: Texas Parks and Wildlife Department; 1974. , Bulletin 41. [Google Scholar]
- 19.Schmidly D J. The Mammals of Trans-Pecos Texas: Including Big Bend National Park and Guadalupe Mountains National Park. College Station: Texas A&M Univ. Press; 1977. [Google Scholar]
- 20.Lord K J. Ph.D. thesis. College Station: Texas A&M University; 1984. [Google Scholar]
- 21.Sobolik K D. N Am Archeol. 1993;14:227–244. [Google Scholar]
- 22.Stiner M C, Munro N D, Surovell T A. Curr Anthropol. 2000;41:39–74. [PubMed] [Google Scholar]
- 23.Sobolik K D. Tex J Sci. 1990;44:23–32. [Google Scholar]
- 24.Hofreiter M, Poinar H N, Spaulding G S, Martin P S, Possnert G, Pääbo S. Mol Ecol. 2000;9:1975–1984. doi: 10.1046/j.1365-294x.2000.01106.x. [DOI] [PubMed] [Google Scholar]
- 25.Handt O, Krings M, Ward R, Pääbo S. Am J Hum Genet. 1996;59:368–376. [PMC free article] [PubMed] [Google Scholar]
- 26.Marlar R A, Leonard B L, Billman B R, Lambert P M, Marlar J E. Nature (London) 2000;407:74–78. doi: 10.1038/35024064. [DOI] [PubMed] [Google Scholar]
- 27.Morell V. Science. 1998;280:190–192. doi: 10.1126/science.280.5361.190. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.