

Abstract
Cells of the craniofacial skeleton are derived from a common mesenchymal progenitor. The regulatory factors that control their differentiation into various cell lineages are unknown. To investigate the biological function of dentin matrix protein 1 (DMP1), an extracellular matrix gene involved in calcified tissue formation, stable transgenic cell lines and adenovirally infected cells overexpressing DMP1 were generated. The findings in this paper demonstrate that overexpression of DMP1 in pluripotent and mesenchyme-derived cells such as C3H10T1/2, MC3T3-E1, and RPC-C2A can induce these cells to differentiate and form functional odontoblast-like cells. Functional differentiation of odontoblasts requires unique sets of genes being turned on and off in a growth- and differentiation-specific manner. The genes studied include transcription factors like core binding factor 1 (Cbfa1), bone morphogenetic protein 2 (BMP2), and BMP4; early markers for extracellular matrix deposition like alkaline phosphatase (ALP), osteopontin, osteonectin, and osteocalcin; and late markers like DMP2 and dentin sialoprotein (DSP) that are expressed by terminally differentiated odontoblasts and are responsible for the formation of tissue-specific dentin matrix. However, this differentiation pathway was limited to mesenchyme-derived cells only. Other cell lines tested by the adenoviral expression system failed to express odontoblast-phenotypic specific genes. An in vitro mineralized nodule formation assay demonstrated that overexpressed cells could differentiate and form a mineralized matrix. Furthermore, we also demonstrate that phosphorylation of Cbfa1 (osteoblast-specific transcription factor) was not required for the expression of odontoblast-specific genes, indicating the involvement of other unidentified odontoblast-specific transcription factors or coactivators. Cell lines that differentiate into odontoblast-like cells are useful tools for studying the mechanism involved in the terminal differentiation process of these postmitotic cells.
Craniofacial skeleton is primarily derived from neural crest-derived mesenchymal cells (1). These cells are responsible for building and maintaining complex, dynamic mineralized tissues such as bone and dentin. Despite their common origin, these tissues are dramatically different, based on their phenotype-specific matrix secretion, the extent of their mineralization, and matrix resorptive properties. The proper mineralization of bones and teeth has immense importance in normal growth and development and musculoskeletal function. Problems in mineralization are evident in a number of skeletal and dental pathologies. The organic matrix appears to have a direct role in regulating the mineralization process.
Biomineralization in vertebrates as well as the invertebrate system use polyanionic proteins to direct oriented crystal growth. In bone and dentin, the nature of the hydroxyapatite crystals deposited is under the precise control of the collagen template and the noncollagenous acidic proteins. Acidic proteins like osteopontin, bone sialoprotein, osteonectin, osteocalcin, bone acidic glycoprotein 75, and dentin matrix protein 1 (DMP1), expressed by both odontoblasts and osteoblasts, have been implicated to have an important role in matrix mineralization through the regulation of crystal size and morphology (2, 3). However, there are a few additional proteins, like dentin phosphophoryn (dentin matrix protein 2, DMP2) (4, 5) and dentin sialoprotein (DSP) (6), that are virtually unique to dentin mineralization. Dentin phosphophoryns are the major acidic noncollagenous proteins in dentin and are unique proteins that have not been found in any tissue other than teeth and are therefore phenotypic markers of odontoblast activity. Dentin sialoprotein is marked by a high content of sialic acid (≈30% by weight) and low phosphorylation. The main feature of the principal noncollagenous proteins is that they are all quite acidic, and several of them are phosphorylated. Dentinogenesis imperfecta type II is a genetic disorder of dentin mineralization, and the absence and defect of noncollagenous protein synthesis have been implicated to cause mineralization defects (7).
The formation and differentiation of odontoblasts, the principal dentin-forming cells, depend on signaling molecules that move back and forth from the adjacent epithelium to the underlying mesenchyme and vice versa (8). Fully differentiated odontoblasts are tall columnar cells with a polarized distribution of their cytoplasmic organelles. Functional differentiation of odontoblasts is characterized by synthesis of type I collagen and noncollagenous proteins. Orchestration of multiple events directs these cells to synthesize and secrete a matrix conducive to the controlled growth of hydroxyapatite crystals.
DMP1 is an acidic protein that was first cloned from the mineralized dentin matrix (9). A single unique feature of DMP1 is that it is highly hydrophilic, as the principal amino acids are glutamic acid, aspartic acid, and serines. The serine residues are embedded in acidic sequences that make them good substrates for phosphorylation by casein kinase I and II. It could be postulated that DMP1, because of its high acidic nature, can bind to calcium, thereby initiating the nucleation process and, furthermore, turn on the entire cascade of regulated hydroxyapatite crystal growth. Although the precise function of DMP1 is yet to be determined, in situ hybridization experiments have demonstrated that it is localized in both osteoblasts and odontoblasts (10). At the amino acid level, DMP1 has a single RGD domain, which was shown recently to be functional in cell attachment (11). A detailed understanding of the physiological function of DMP1 is still lacking, even though this protein has been well characterized.
Our hypothesis is that DMP1 is an essential factor in the terminal differentiation of ectomesenchyme-derived neural crest cells into functional odontoblasts. Understanding the mechanisms that control specialized mesenchyme-derived cells like osteoblasts and odontoblasts in the differentiation and production of a well-ordered mineralized matrix has been the objective of this study. To explore the role of DMP1 in the differentiation of odontoblasts, we genetically overexpressed DMP1 in preosteoblastic MC3T3-E1 cells and pluripotent embryonic mesenchymal C3H10T1/2 cells. The C3H10T1/2 cells have the ability to differentiate into myoblasts, adipocytes, chondrocytes, or osteoblasts in the presence of appropriate regulatory factors in vitro (12). MC3T3-E1 cells are mouse calvaria-derived cells and have been described as relatively immature committed osteoblasts, which differentiate in response to various cellular stimulants (13). Adenovirus mediated gene expression is a well-established, highly efficient gene transfer technique. Adenovirus-mediated DMP1 overexpression was used to analyze early gene expression in both C3H10T1/2 and MC3T3-E1 cells. Thus the effect of DMP1 overexpression was tested by two experimental approaches, one by stable transfections and the other by adenovirus-mediated overexpression.
Materials and Methods
Plasmid Construction.
DMP1 cDNA was PCR amplified and cloned into pSTBlue-1 (Novagen) vector. Two colonies having both sense and antisense orientations of DMP1 were then subcloned into KpnI/XhoI sites of pcDNA3.1 vector (Invitrogen). These plasmids were named pcDNA3.1-DMP1 (sense) and pcDNA3.1-DMP1 (antisense). They were also used in the construction of the adenoviral vectors.
Construction of DMP1-Expressing Adenovirus.
An E1a-deleted, replication-deficient recombinant adenovirus carrying an expression cassette for rat DMP1 was prepared by the direct ligation method as described by Stow (14). In this procedure the DMP1 cDNA released from pcDNA 3.1-DMP1 (sense) by HindIII and XhoI was subcloned into cytomegalovirus (CMV) general vector at these sites. The pCMV general vector contains approximately the first (left side) 400 bp of the human type 5 adenovirus (Ad5) genome followed by the CMV promoter/enhancer, the DMP1 coding region, and an simian virus 40 polyadenylation sequence. The resulting pCMV-DMP1 was digested with XbaI to release a 3.4-kb fragment containing the Ad5 left-side obligate packaging sequence together with the DMP1 expression cassette. This 3.4-kb fragment was ligated to the right-side 32.7-kb of the Ad5 dl 309/356 genome. The ligated viral DNA was transfected into 293 cells (a human embryonic kidney cell line that expresses E1a). The resulting virus [Ad5/CMV-DMP1 (sense)] was harvested from infected monolayers, plaque purified (twice), and propagated in 293 cells. The viral titer after final propagation was 2 × 109 plaque-forming units (pfu)/ml. Ad5/CMV–DMP1 (antisense) was made in an identical manner, and the viral titer after final propagation was 1.8 × 109 pfu/ml. The recombinant adenoviruses (both sense and antisense) were tested by restriction analysis and by PCR, with the use of respective primers for the left-side packaging sequence and the E1a region. The control Ad5/CMV-lacZ, a recombinant adenovirus carrying an expression cassette for the Escherichia coli lacZ gene (a gift from Dr. Christopher Newgard, University of Texas Southwestern Medical Center), was prepared as described by Herz and Gerard (15). The viral titer after final propagation was 1 × 109 pfu/ml.
Cell Culture, Transfections, and Adenoviral Infections.
Embryonic undifferentiated mesenchymal C3H10T1/2 cells obtained from American Type Culture Collection were cultured in basal medium eagle (BME) medium (Celgro; Chicago, IL) supplemented with 10% FBS (GIBCO/BRL). MC3T3-E1 mouse preosteoblastic cells were cultured with DMEM supplemented with 10% heat-inactivated FBS (Celgro), 1% penicillin and streptomycin, and 1% Fungizone. Stable transfections with sense and antisense plasmid DNA were performed with Superfect (Qiagen, Chatsworth, CA) according to the manufacturer's protocol. Selections of the stable colonies were made with G418 sulfate at 0.8 μg/ml of medium. Single cell colonies were isolated by plating cells at low density, and six individual colonies were picked with the use of cloning discs (Fisher Scientific). Control cells were mock transfected with “empty” vector and selected identically. Adenoviral infections were performed on cells at 80–85% confluence in six-well plates. Typically, 100 pfu per cell viral solution was used, and infection was allowed for 1 h at 37°C and further continued in the presence of regular medium for 24 h. Cells were harvested for further analysis.
Other nonosteogenic cell lines, J774 (macrophages), RPC-C2A (rat pulp cells), C2C12 (myoblasts), and NIH 3T3 (embryonic fibroblast), were cultured in the presence of DMEM media supplemented with 10% FBS.
Morphological Observation.
Cells were routinely observed under a phase-contrast microscope to evaluate their overall appearance. Microphotographs were taken with ×10 objective and a Nikon camera.
Northern Blotting.
Total RNA was isolated from cultured cells at log phase with the use of TriZol reagent (GIBCO/BRL). RNA gel electrophoresis and Northern blotting were done as described by Sambrook et al. (16). Twenty micrograms of total RNA was resolved on a 0.8% agarose gel containing formaldehyde. The RNA was transferred to a Hybond nylon membrane (Amersham Pharmacia). The membrane was prehybridized with the use of HyperHyb (Research Genetics, Huntsville, AL) and probed with randomly labeled (Decaprime kit; Ambion, Austin, TX), appropriate probes.
Reverse Transcription–PCR (RT-PCR).
DNaseI (RNase free, RQ1; Promega)-treated RNA was used for all RT-PCR reactions. Three micrograms of total RNA was reverse transcribed for 90 min at 42°C with Superscript II (GIBCO/BRL). PCR Supermix (GIBCO/BRL) was used in all of the PCR reactions. Primers for the PCR reaction were designed from available sequences at the National Center for Biotechnology Information gene databank. The PCR products were verified either by sequencing or by restriction mapping. For DMP2 the primers were designed from the untranslated region.
In Vitro Assay for Induction of Mineralized Nodule Formation.
The mineralization microenvironment was created by treating the MC3T3 cells (80–90% confluent) with 10 mM β-glycerophosphate and 100 μg/ml ascorbic acid along with 10 nM dexamethasone, and C3H10T1/2 cells were induced with 7 mM β-glycerophosphate and 100 μg/ml ascorbic acid along with 10 nM dexamethasone.
Mitogen-Activated Protein Kinase (MAPK)/Casein Kinase II Inhibitor.
The activation of MAPK in MC3T3-E1 and C3H10T1/2 overexpressed cells was inhibited by treatment with specific mitogen-activated protein kinase kinase (MEK) inhibitor PD98059 (Sigma) or by heparan sulfate (Sigma), an inhibitor for casein kinase II, based on published experimental protocols (17). The cells were infected with the adenoviral constructs as described earlier, in the presence or absence of specific inhibitors. Protein and total RNA were isolated from these cells, and expression of care binding factor (Cbfa1), osteocalcin (OCN), DSP, and DMP2 was analyzed by RT-PCR. The proteins were immunoprecipitated with the use of affinity-purified DMP1 antibody and analyzed for DMP1 phosphorylation with the use of monoclonal phosphoserine antibody (Sigma).
Western Blot Analysis and Immunoprecipitation.
Total protein was extracted as described by Harlow and Lane (18), and the concentration was estimated with the use of Bradford microassay (Bio-Rad) reagent. Twenty-five micrograms of total protein was resolved on a 10% SDS-PAGE gel, transferred electrophoretically to nitrocellulose membrane, and subsequently probed with the appropriate antibody. Immunoprecipitation was performed by incubating the total protein with affinity-purified anti-DMP1 antibody for 2 h at 24°C. The antigen–antibody complex was precipitated with the use of protein G agarose (Sigma) and subsequently resolved on a 10% SDS-PAGE gel.
Imaging and Quantitation.
All photographs and Northern blots were scanned and labeled with the use of adobe photoshop (version 5.5) software. A Kodak Digital Imaging system was used for quantification.
Results
DMP1 Overexpression.
Northern blot analysis confirmed that the transfected cells were overexpressing (10- to 12-fold) DMP1 gene when compared with control cells (Fig. 1A). The presence of DMP1 in cell extracts was detected by Western blotting with affinity-purified DMP1 antibody (Fig. 1B). Positive results were seen only in sense constructs in both the stable cell lines and in recombinant adenovirus-transduced cells. The antisense overexpressing cells completely inhibited DMP1 translation (Fig. 1B).
Figure 1.
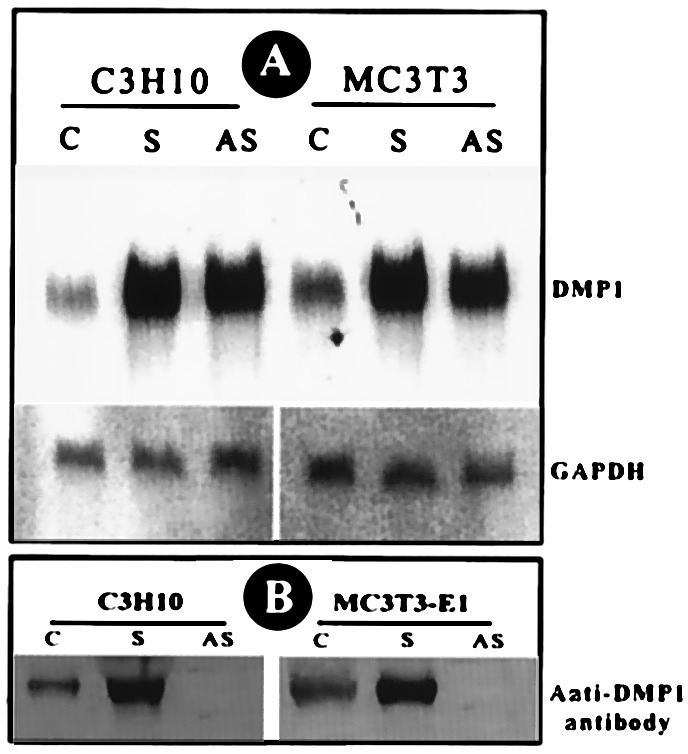
Overexpression of DMP1. The cell lines were transfected as described in Materials and Methods. Stable selection was made at 800 μg/ml of G418 sulfate. The expression level of DMP1 was analyzed by either Northern blot (A) or Western blot (B). Twenty micrograms of total RNA was resolved on a 0.8% agarose gel under denaturing conditions and transferred. The membrane was probed with random [32P]dCTP-labeled DMP1 cDNA or glyceraldehyde-3-phosphate dehydrogenase. Fifteen micrograms of total protein was resolved on a 10% SDS-PAGE gel and subsequently electroblotted. The membrane was probed with affinity-purified anti-DMP1 antibody (B).
DMP1 Overexpression and Cell Morphology.
Mock MC3T3-E1 and C3H10T1/2 cells maintained a fibroblast-like appearance with a polygonal pattern at confluence (Fig. 2A). Cells transfected with sense DNA exhibited profound changes in their morphology at both low and high density. These cells were columnar and polarized, tended to align themselves in straight parallel lines, and had long dendrite-like processes (Fig. 2B). In some instances these alignments can be traced for a distance of 4–5 mm. In contrast, cells overexpressing antisense DMP1 did not exhibit any characteristic morphological change (Fig. 2C).
Figure 2.

DMP1 overexpression and morphology. The G418-resistant cells were plated at high density (3 × 106 cells per milliliter) and low density (1 × 104 cells per milliliter). The cells were allowed to grow in the appropriate medium for 24 h, and photographs were taken with a phase-contrast microscope. “Mock” represents a stable cell line expressing the “empty” vector (A). “Sense” (B) and “antisense” (C) represent the stable cell lines overexpressing sense and antisense DMP1 cDNA. Microphotographs were taken at ×10 with a Nikon camera.
DMP1 Overexpression and Changes in Gene Expression.
We next focused on correlating morphological changes with the expression of unique sets of genes being turned on in a growth- and differentiation-specific manner. Northern blot analysis (Fig. 3) demonstrated that overexpression of DMP1 triggered the differentiation of C3H10T1/2 cells into odontoblast-like cells, with enhanced expression of alkaline phosphate (ALP), Cbfa1, and bone morphogenetic protein 4 (BMP4) corresponding to early-stage markers and bone sialoprotein (BSP), OCN, osteopontin (OPN), and osteonectin corresponding to the terminally differentiated state. As expected, the mock C3H10T1/2 cells (expressing the vector alone) and cells with DMP1 in the antisense orientation did not express any of the odontoblast-specific genes (Fig. 3). On the other hand, mock MC3T3-E1 cells did express most of the osteoblast phenotype-specific genes at different levels (Fig. 3), as they have been reported to be immature preosteoblastic cells. An interesting observation was that DMP1 in the antisense orientation inhibited expression of endogenous osteocalcin and alkaline phosphatase in MC3T3-E1 cells.
Figure 3.
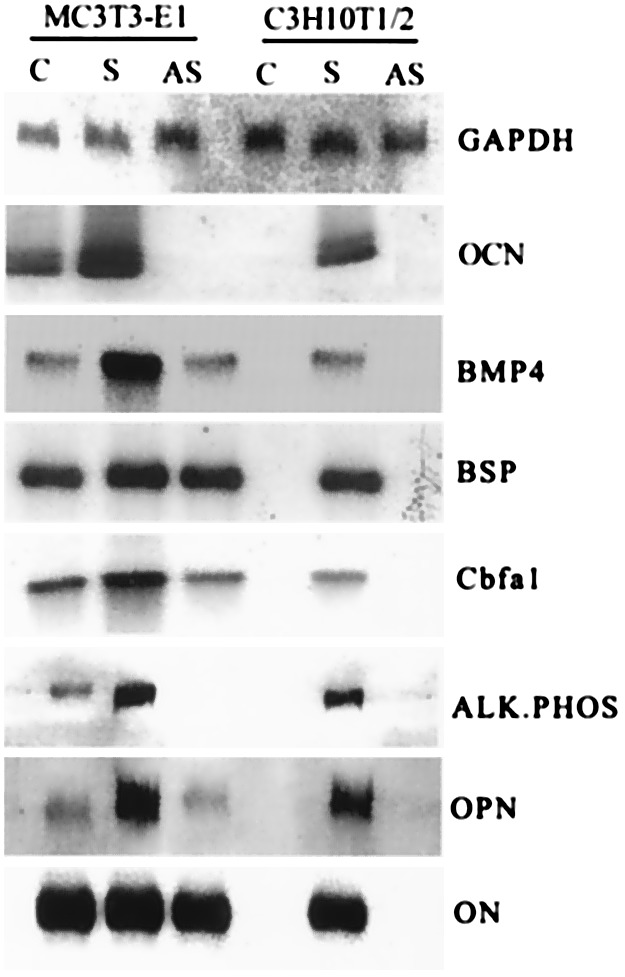
Northern analysis of various marker genes. Total RNA was isolated from G418-resistant cell lines overexpressing DMP1. Northern blot was carried out with 20 μg of RNA. The probes were randomly labeled with [32P]dCTP. C, Stable MC3T3-E1 and C3H10T1/2 cell lines expressing the “empty vector.” S, Stable MC3T3-E1 and C3H10T1/2 cell lines overexpressing DMP1 in sense orientation. AS, Stable MC3T3-E1 and C3H10T1/2 cell lines overexpressing DMP1 in the antisense orientation. Glyceraldehyde-3-phosphate dehydrogenase was used as the control.
DMP1 Overexpression Induces DMP2 and DSP Expression.
The most unexpected finding in this study was the expression of odontoblast-specific markers DMP2 and DSP during the cellular differentiation process (Fig. 4). DMP2 is the systematic name we have applied to dentin phosphophoryn (5), a dentin-specific protein that has been isolated biochemically. DSP is another apparently dentin-specific matrix protein, second only to the phosphophoryn in content in dentin. To determine whether DMP1 is responsible for the induction of these phenotypic products, cell lines harboring the antisense DMP1 construct was used to check the expression of DMP2 and DSP. Consistent with our hypothesis, the antisense construct completely inhibited the expression of DSP and DMP2 in both cell lines (Fig. 4).
Figure 4.
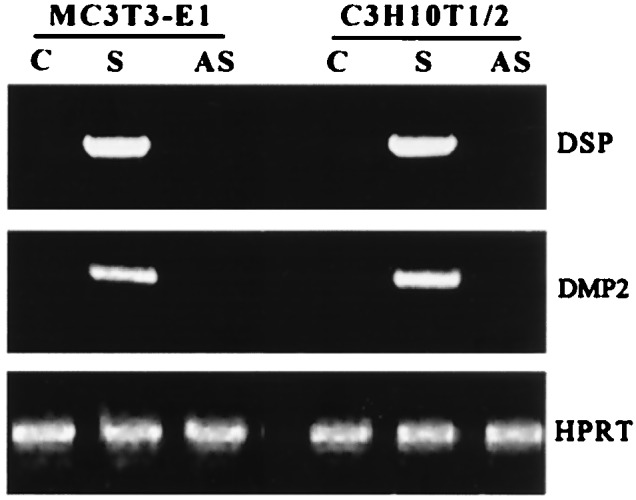
Expression of DSP and DMP2. G418-resistant cells were analyzed by RT-PCR as described in Materials and Methods. C, Mock MC3T3-E1 and C3H10T1/2. S, Stable MC3T3-E1 and C3H10T1/2 cell lines overexpressing DMP1 (sense). AS, Cell lines overexpressing the antisense sequence of DMP1. PCR products were resolved on a 2% agarose gel.
Moreover, the preosteoblastic cell line (MC3T3-E1) as well as the embryonic mesenchymal cell line (C3H10T1/2) had the potential to differentiate into either osteoblast or odontoblast among the pool of stable transgenic cells. However, six single cell colonies isolated for each of the overexpressing cell lines expressed Cbfa1, DSP, and DMP2 (data not shown). This observation was again consistent with our hypothesis that DMP1 was essential for the terminal differentiation of odontoblasts.
To confirm the direct role of DMP1 in the expression of DMP2 and DSP, in an independent study the DMP1 protein translation was inhibited specifically by the antisense DMP1 mRNA with the use of the adenoviral system. As expected, antisense DMP1 completely inhibited the expression of DMP2 and DSP (Fig. 5).
Figure 5.

Suppression of DSP and DMP2 by antisense DMP1. Cell lines overexpressing DMP1 (sense) were infected with Ad5/CMV-DMP1 (antisense) as described in Materials and Methods. Total RNA was isolated (after 24 h), and RT-PCR was performed with gene-specific primers. C, Control cells (with “empty vector”). S, cells overexpressing sense DMP1. S + AS infected with Ad5-CMV-DMP1 (antisense) on cells overexpressing DMP1 (sense). In each case DSP and DMP2 were analyzed by RT-PCR.
DMP1 Overexpression and Specificity of the Responding Cells.
To determine the cell type specificity, adenoviral construct anchoring rat DMP1 [Ad5/CMV-DMP1 (sense)] was tested on preosteoblastic and nonosteoblastic cells (Fig. 6). Of the different cell types tested, the interesting observation was that only preosteoblastic cell line MC3T3-E1 (Fig. 4), C3H10T1/2 (Fig. 4), and rat pulp cells (RPC-C2A) (Fig. 6) responded to DMP1 overexpression stimulus and expressed DMP2 and DSP. Cbfa1 expression was turned on in C2C12 cells as well as in RPC-C2A cells with a 4-fold increase in its expression. Interestingly, OCN was not expressed in RPC-C2A cells and was present in C2C12 cells. On the other hand osteonectin was expressed in a variety of cells (NIH 3T3, RPC-C2A, and C2C12), and OPN was turned on in NIH 3T3 and C2C12 cells (Fig. 6). Thus the transcriptional regulation of DMP2 and DSP by DMP1 is restricted to mesenchyme-derived cells only.
Figure 6.
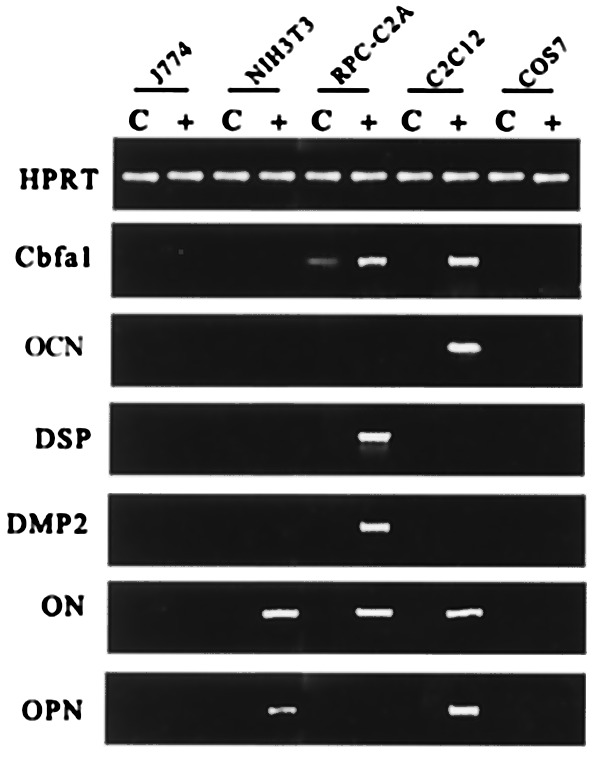
Cell specificity. To study cell specificity, the Ad5/CMV-DMP1 (sense) and Ad5/CMV-lacZ adenoviruses were used. Different cell lines were infected with adenovirus for 3 h, and fresh medium was then added. Total RNA was isolated (after 24 h), and RT-PCR performed as described in Materials and Methods. C, Control cells infected with Ad5/CMV-lacZ. +, Cells infected with Ad5/CMV-DMP1 (sense). Hypoxanthine phosphoribosyltransferase was used as the control, and gene expression levels of Cbfa1, OCN, DSP, DMP2, osteonectin, and OPN were analyzed.
Expression of DMP2 and DSP Is Independent of Cbfa1 Phosphorylation by MAPK and CK-II.
To further explore the basis of the differentiation potential of the overexpressed cells, the mechanism for the expression of the transcription factor Cbfa1 was investigated. This factor has been well studied and has all of the properties of a differentiation factor for the osteoblast lineage. During embryonic development Cbfa1 expression precedes osteoblast differentiation and is restricted to mesenchymal cells destined to become either chondrocytes or osteoblasts (19). It also regulates the expression of osteogenic-specific genes. Cbfa1 is also expressed in odontoblasts (20), but its function in odontogenesis is not well understood. Phosphorylation of Cbfa1 by the MAPK pathway was shown to play an important role in the transcriptional regulation of osteoblast-specific gene expression (21). To determine whether these signals were important in odontoblast formation and differentiation, MAPKs MEK1/MEK2 were blocked by PD98059, a selective inhibitor. Inhibition of Cbfa1 phosphorylation by PD98059 did not block expression of DSP and DMP2 as determined by RT-PCR, but there was a drastic reduction in the expression level of osteocalcin in both cell lines (Fig 7B).
Figure 7.
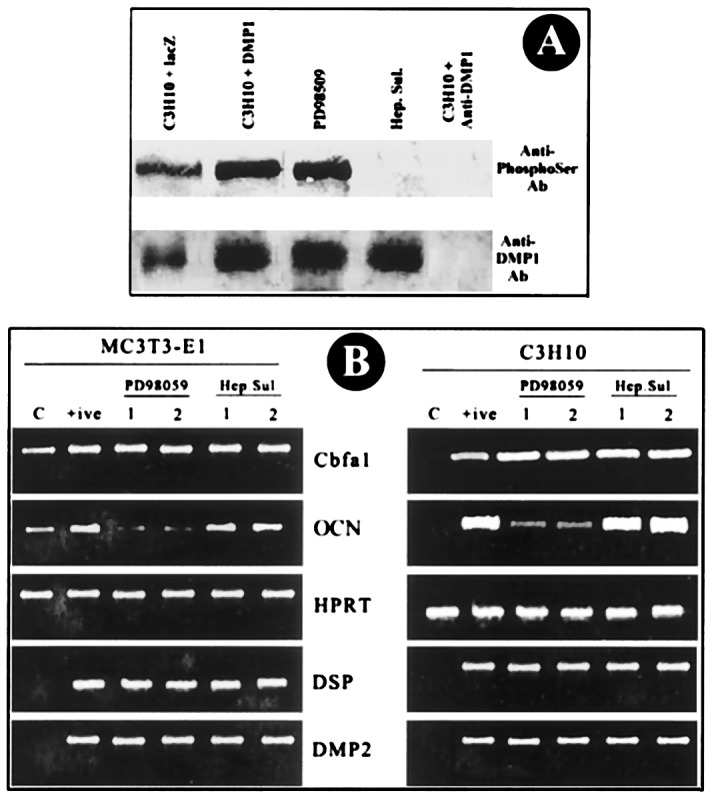
Signaling pathway in DSP/DMP2 expression. (A) DMP1 phosphorylation. C3H10T1/2 cells were infected with adenoviral constructs. Total protein extracts were prepared as described in Materials and Methods and immunoprecipitated with affinity-purified anti-DMP1 antibody. (Top) Western blot against anti-phosphoserine antibody on DMP1 immunoprecipitates. (Bottom) Western blot performed with anti-DMP1 antibody on the DMP1 immunoprecipitates. (B) DSP and DMP2 expression. Cells (2 × 106 cells per milliliter) were seeded 24 h before infection with Ad5/CMV-DMP1. Infection was carried out for 3 h, and fresh medium was added to the cells. A MAPK inhibitor (PD98059, 100 μg/ml) and a casein kinase II inhibitor (heparan sulfate, 100 μg/ml) were added along with fresh medium and incubated for another 24 h. Total RNA was isolated from cells, and RT-PCR was performed. C, Control cells infected with β-gal expression cassette. +, Positive control used for the experiment [infected with Ad5/CMV-DMP1 (sense) without inhibitors]. 1 and 2, Duplicates for the experiment. RT-PCR analysis for Cbfa1, OCN, DSP, and DMP2 was done with hypoxanthine phosphoribosyltransferase as a control.
To determine whether phosphorylated DMP1 played a significant role in the transcriptional activity of DMP2 and DSP, heparan sulfate, a known inhibitor for casein kinase II, was used in this study. RT-PCR analysis of the treated and untreated cells showed that the expression levels of DMP2 and DSP were not affected by inhibition of DMP1 phosphorylation. The expression levels of Cbfa1 and OCN also remained unaltered. The status of DMP1 phosphorylation with or without heparan sulfate and PD98509 was analyzed by immunoprecipitation by anti-DMP1 antibody and cross-probed with phosphoserine antibody. Results in Fig. 7A demonstrate that only heparan sulfate inhibited DMP1 phosphorylation and not MAPK inhibitor (Fig. 7A), confirming the results published earlier (9). Together these results confirm that DMP2 and DSP expression in the overexpressed cells is independent of phosphorylation of DMP1 by casein kinase II pathway.
DMP1 Overexpression and Induction of Mineralization.
Finally an in vitro nodule formation assay was carried out to determine whether the up-regulation of odontoblast-specific gene transcription resulted in a mineralized matrix formation. Nodule formation due to secretion of extracellular matrix proteins in the presence of phosphate ions and ascorbic acid has been considered to be an important feature for mineralization and precedes mineralization. It was observed that overexpression of DMP1 enhanced the onset of mineralization in both MC3T3-E1 (Fig. 8A) and C3H10T1/2 (Fig. 8B), the size of the nodule formed (Fig. 8 C and D) was significantly large, and the overall kinetics of nodule formation was favored by at least 4- to 5-fold in transgenic cell lines when compared with the mock cells. Consistent with our hypothesis, the cells overexpressing antisense DMP1 failed to mineralize despite prolonged treatment with ascorbic acid and β-glycerophosphate (data not shown).
Figure 8.

DMP1 overexpression and mineralization: a graphical representation of the effect of DMP1 overexpression on mineralized nodule formation in MC3T3-E1 (A) and C3H10T1/2 cells (B). (C) The size of the mineralized nodule was measured with an occulometer. Mock cells expressing the empty vector were used as a control. (D) The mineralized nodule was microphotographed (after 15 days) with a Nikon camera at ×10.
Discussion
The molecular mechanisms underlying the differentiation of neural crest-derived ectomesenchymal cells into odontoblasts are not clear. Various morphogens, cytokines, extracellular matrix molecules, and cell–cell interactions have been implicated in embryonic ectomesenchymal differentiation into highly polarized secretory odontoblasts (22). This differentiation is a multistep process that is highly orchestrated at every stage of development. In this paper on the potential of DMP1, an extracellular matrix macromolecule present in mineralized tissues, we report on the initiation of the process of odontogenic-specific cell differentiation.
A finding in this study is that stable transfection of cDNA encoding for rat DMP1 in C3H10T1/2 and MC3T3-E1 cells induced these cells to differentiate into functional odontoblast-like cells with definite changes in size and shape. Changes in cell morphology were well correlated with transcriptional up-regulation of odontoblast-specific genes DMP2 and DSP, which are virtually unique to dentin. A second line of evidence indicating that these were odontoblast-like cells derives from the observation that the extracellular matrix laid down by these cells eventually calcified and formed well-defined mineralized nodules. The ability of antisense DMP1 to inhibit induction of DSP and DMP2 suggests that DMP1 must act as an important molecule during the differentiation of neural crest-derived mesenchymal cells into terminally differentiated odontoblasts in vivo.
The transgenic cell lines clearly showed characteristic morphological and polarization changes that were not observed in the control cells. However, changes in structural morphology were restricted only to mesenchyme-derived C3H10T1/2, MC3T3-E1, and RPC-C2A cells, whereas other cells of different lineages remained identical to the control. Thus our findings suggest that the mesenchymal cell–DMP1 interactions at the extracellular level may be responsible for several cellular signaling pathways leading to their ultimate differentiation into odontoblasts, with changes in cell shape and morphology. Such changes were also reported when MC3T3-E1 was grown on discrete domains of laminin (23). Extracellular matrix components are not only known to promote attachment and differentiation of different types of cells, but are also associated with profound changes in cellular morphology related to cell–cell and cell–matrix interactions (24).
Expression of several transcription factors and differentiation-specific gene expression both in vivo and in vitro confirmed DMP1-overexpressing cells as odontoblasts. Genetically engineered C3H10T1/2 cells had the ability to express transcription factors like BMP4 and Cbfa1; early and proliferative stage markers for odontogenesis like ALP, OPN, OCN, and osteonectin; as well as odontogenic secretory stage-specific phenotypic markers DSP and DMP2, which are responsible for dentin extracellular matrix deposition and maturation. However, cells overexpressing antisense DMP1 had the same expression profile as that of the mock-transfected cells. The dentin-specific DSP and phosphophoryn genes are normally synthesized by odontoblasts only after they are terminally differentiated and enter the secretory phase. Thus a strong correlation has been unveiled between changes in cellular morphology and odontogenic-specific gene expression patterns.
In contrast, cells transfected with DMP1 in the antisense orientation were particularly interesting. Blockage of DMP1 expression abrogated the expression of ALP and OCN. The down-regulation of OCN and ALP in the absence of DMP1 (by the expression of antisense DMP1) suggests that DMP1 could turn on a specific signal for induction of these genes. ALP is an early marker for both osteoblasts and odontoblasts. This enzyme plays a vital role in calcified tissue formation, probably by regulating phosphate transport (25). OCN, on the other hand, is produced by mature osteoblasts and odontoblasts during mineralization, and the message level is elevated during both cell proliferation and differentiation. Moreover, increases in the expression levels of BMP4, Cbfa1, ALP, and OPN in sense overexpressing MC3T3-E1 cells suggests that DMP1 plays a significant role either directly or indirectly in the transcriptional mechanism of these genes as well as in the differentiation process. Similarly, Sox9, MyoD, and peroxisome-proliferator-activated receptor gamma2 (PPARγ2) have been reported as essential transcription factors that trigger the determinative switch to chondrocytes (26), skeletal muscle (27), or adipose tissue (28) formation, respectively.
In an independent study, it was demonstrated that infection of these two cell lines with the recombinant adenovirus triggered the expression of DSP and DMP2 as well as demonstrating morphological transformation. Control cells infected with lacZ or the antisense DMP1 did not express DMP2 and DSP, and no morphological changes were observed. The direct immediate effect of expression of DMP1 on the transcription of DSP and DMP2 further strengthens our hypothesis that DMP1 can act as a signaling molecule in the odontogenic induction process and thus is capable of programming multipotential cells to differentiate into odontoblastic lineage.
The ability of DMP1 to induce the signal of the DSP and DMP2 message depends on cell specificity. Interestingly, only mesenchyme-derived cells like C3H10T1/2, MC3T3-E1, and RPC-C2A responded to the differentiation signal. Cell lines from other lineages, when infected with DMP1 adenoviral construct, abrogated expression of dentin phenotypic products. Furthermore, these cells failed to express most of the other noncollagenous proteins present in the dentin matrix. Thus our results indicate that the signal from DMP1 may be an integral part of a tightly coordinated program responsible for the induction of terminal differentiation of odontoblasts.
To characterize the molecular signals that are necessary for establishing odontogenesis, the expression of Cbfa1, an essential transcription factor for the development of the osteoblastic phenotype, was studied. Induction of significant expression of Cbfa1 in overexpressed cells and consequently the expression of dentin-specific DMP2 and DSP suggests that Cbfa1 may be a downstream transcription factor necessary for odontoblast differentiation. One of the interesting observations in this study is the regulation of the OCN gene by Cbfa1 expression in the MC3T3-E1 antisense DMP1 cell line. OCN is a known target gene for the osteoblast-specific transcription factor Cbfa1, and its promoter has four Cbfa1-binding sites (OSE2) (29). Earlier studies on the OCN promoter showed that mutations incorporated at the OSE2 sites completely abolished the promoter activity (29). However, in MC3T3-E1 cells overexpressing antisense DMP1 the expression level for the OCN message is below the detectable level, whereas the Cbfa1 expression level was unaltered compared with that of the mock cells. Our findings here suggest that there are other transcription factors in addition to Cbfa1 in the control of the expression of OCN in vivo. Our data also correlate well with the published studies on Cbfa1 phosphorylation (17). Earlier studies had demonstrated that phosphorylation of Cbfa1 by the MAPK pathway is essential for its transcriptional activity. Inhibition of Cbfa1 phosphorylation by the MEK1/MEK2-specific inhibitor PD98059 was shown to completely inhibit OCN promoter activity (17). However, our data obtained with the use of the MEK1/MEK2 inhibitor demonstrated reduction in the expression level of OCN by ≈5-fold compared with the lacZ-infected cells. Results on OCN expression correlated well with the Cbfa1 knockout studies (30). Together, these observations clearly suggest the involvement of other transcription factors/coactivators/inhibitors in the regulation of OCN gene expression in addition to Cbfa1.
The exact biochemical function of DMP1 in either bone or dentin is not known, but functional motifs for phosphorylation have offered clues. Results from our present study indicate that heparan sulfate, a known inhibitor of casein kinase II, inhibits serine phosphorylation in DMP1 and simultaneously has no effect on the expression levels of DSP and DMP2. One of the potential signal transduction pathways that might regulate the differentiation of these overexpressed cells into odontoblasts is the MAPK pathway, as activation of the MAPK pathway in other cell types such as neuronal cells promotes differentiation (31). Experimental observations with the MAPK inhibitor PD98059 led to the conclusion that the phenotypic expression of DMP2 and DSP is independent of phosphorylation by MAPKs in the overexpressed cells.
Mineralization of the dentin matrix may be considered a hallmark of mature, secretory odontoblasts. In vitro mineralization assay demonstrated that the progenitor overexpressed cells differentiated to a functional matrix synthesizing odontoblasts, depositing a collagenous extracellular matrix accompanied by the activation of DMP2 and DSP. The size of the mineralized nodule formed increased with the number of days in culture, which indicated increased expression of genes, thought to contribute to the differentiated state of the odontoblasts. Thus the overexpressed cell lines have exhibited a dynamic odontogenic differentiation process.
Changes in cell differentiation and morphogenesis induced by DMP1 might also require several transcriptionally regulated events. Relatively little is known regarding the mechanisms by which DMP1 can regulate such diverse cellular responses. Transcription factors and signaling pathways that regulate odontoblast differentiation and expression of tissue-specific genes have yet to be identified. This information will be important in defining on a molecular level the gene regulatory pathways required for the growth and differentiation of odontoblasts.
Acknowledgments
We thank our colleagues Dr. Srilata Bagchi and Mr. Gen He for their valuable comments and critical reading of the manuscript. This research was supported by National Institutes of Health Grant DE 11657.
Abbreviations
- DMP1
dentin matrix protein 1
- DSP
dentin sialoprotein
- CMV
cytomegalovirus
- pfu
plaque-forming unit
- RT-PCR
reverse transcription–PCR
- MAPK
mitogen-activated protein kinase
- OCN
osteocalcin
- OPN
osteopontin
- Cbfa1
core binding factor 1
- BMP
bone morphogenetic protein
- ALP
alkaline phosphatase
- MEK
mitogen-activated protein kinase kinase
- BSP
bone sialoprotein
References
- 1.Erickson C A, Reedy M V. Curr Top Dev Biol. 1998;40:177–209. doi: 10.1016/s0070-2153(08)60367-1. [DOI] [PubMed] [Google Scholar]
- 2.Linde A, Goldberg M. Crit Rev Oral Biol Med. 1993;4:679–728. doi: 10.1177/10454411930040050301. [DOI] [PubMed] [Google Scholar]
- 3.Veis A. In: Extracellular Matrix. Comper W D, editor. London: Harwood; 1996. pp. 41–76. [Google Scholar]
- 4.Dimuzio M T, Veis A. Calcif Tissue Res. 1978;25:169–178. doi: 10.1007/BF02010765. [DOI] [PubMed] [Google Scholar]
- 5.George A, Bannon L, Sabsay B, Dillon J W, Malone J, Veis A, Jenkins A N, Gilbert D J, Copeland N G. J Biol Chem. 1996;271:32869–32873. doi: 10.1074/jbc.271.51.32869. [DOI] [PubMed] [Google Scholar]
- 6.D'Souza R N, Bronckers A L J J, Happonen R-P, Doga D A, Farach-Carson M C, Butler W T. J Histochem Cytochem. 1992;40:359–366. doi: 10.1177/40.3.1552175. [DOI] [PubMed] [Google Scholar]
- 7.Takagi Y, Sasaki S. J Oral Pathol. 1986;17:208–212. doi: 10.1111/j.1600-0714.1988.tb01526.x. [DOI] [PubMed] [Google Scholar]
- 8.Chen Y, Zhang Y, Jiang T-X, Barlow A J, Amand T R S, Hu Y, Heaney S, Francis-West P, Chuong C-M, Maas R. Proc Natl Acad Sci USA. 2000;97:10044–10049. doi: 10.1073/pnas.160245097. . (First Published August 22, 2000; 10.1073/pnas.160245097) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.George A, Sabsay B, Simonian P A L, Veis A. J Biol Chem. 1993;268:12624–12630. [PubMed] [Google Scholar]
- 10.D'Souza R N, Cavender A, Sunavala G, Alvarez J, Ohshima T, Kulkarni A B, MacDougall M. J Bone Miner Res. 1997;12:2040–2049. doi: 10.1359/jbmr.1997.12.12.2040. [DOI] [PubMed] [Google Scholar]
- 11.Kulkarni G V, Chen B, Malone J P, Narayanan A S, George A. Arch Oral Biol. 2000;45:475–484. doi: 10.1016/s0003-9969(00)00010-8. [DOI] [PubMed] [Google Scholar]
- 12.Atkinson B L, Fantle K S, Benedict J J, Huffer W E, Gutierrez-Hartmann A. J Cell Biochem. 1997;65:325–339. [PubMed] [Google Scholar]
- 13.Franceschi R T, Iyer B S. J Bone Miner Res. 1992;7:235–246. doi: 10.1002/jbmr.5650070216. [DOI] [PubMed] [Google Scholar]
- 14.Stow N D. J Virol. 1981;37:171–180. doi: 10.1128/jvi.37.1.171-180.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Herz J, Gerard R D. Proc Natl Acad Sci USA. 1993;90:2812–2816. doi: 10.1073/pnas.90.7.2812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sambrook J, Fritch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 17.Dudley D T, Pang L, Decker S J, Bridges A J, Saltiel A R. Proc Natl Acad Sci USA. 1995;92:7686–7689. doi: 10.1073/pnas.92.17.7686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Harlow E, Lane D. Antibodies: A Laboratory Manual. Plainview, NY: Cold Spring Harbor Lab. Press; 1988. [Google Scholar]
- 19.Franceschi R T. Crit Rev Oral Biol Med. 1999;10:40–57. doi: 10.1177/10454411990100010201. [DOI] [PubMed] [Google Scholar]
- 20.D'Souza R N, Aberg T, Gaikwad J, Cavender A, Owen M, Karsenty G, Thesleff I. Development (Cambridge, UK) 1999;126:2911–2920. doi: 10.1242/dev.126.13.2911. [DOI] [PubMed] [Google Scholar]
- 21.Ducy P, Michael S, Priemel M, Shen J, Pinero G, Geoffroy V, Amling M, Karsenty G. Genes Dev. 1999;13:1025–1036. doi: 10.1101/gad.13.8.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Maas R, Bei M. Crit Rev Oral Biol Med. 1997;8:4–39. doi: 10.1177/10454411970080010101. [DOI] [PubMed] [Google Scholar]
- 23.Vukicevic S, Luyten F R, Kleinman H K, Reddi A H. Cell. 1990;63:437–445. doi: 10.1016/0092-8674(90)90176-f. [DOI] [PubMed] [Google Scholar]
- 24.Vuori K, Ruoslahti E. Nat Cell Biol. 1999;1:E85–E87. doi: 10.1038/12081. [DOI] [PubMed] [Google Scholar]
- 25.Beck G R, Jr, Zerler B, Moran E. Proc Natl Acad Sci USA. 2000;97:8352–8357. doi: 10.1073/pnas.140021997. . (First Published July 11, 2000; 10.1073/pnas.140021997) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.de Crombrugghe B, Lefebvre V, Behringer R R, Bi W, Murakami S, Huang W. Matrix Biol. 2000;19:389–394. doi: 10.1016/s0945-053x(00)00094-9. [DOI] [PubMed] [Google Scholar]
- 27.Perry R L, Rudnick M A. Front Biosci. 2000;5:D750–D767. doi: 10.2741/perry. [DOI] [PubMed] [Google Scholar]
- 28.Rosen E D, Walkey C J, Puigserver P, Spiegelman B M. Genes Dev. 2000;14:1293–1307. [PubMed] [Google Scholar]
- 29.Schinke T, Karsenty G. J Biol Chem. 1999;274:30182–30189. doi: 10.1074/jbc.274.42.30182. [DOI] [PubMed] [Google Scholar]
- 30.Ducy P, Zhang R, Geoffroy V, Ridall A L, Karsenty G. Cell. 1997;89:747–754. doi: 10.1016/s0092-8674(00)80257-3. [DOI] [PubMed] [Google Scholar]
- 31.Burden S J. Cell. 2000;100:495–497. doi: 10.1016/s0092-8674(00)80685-6. [DOI] [PubMed] [Google Scholar]