

Abstract
Background
Nitrogen fixation gene expression in Sinorhizobium meliloti, the alfalfa symbiont, depends on a cascade of regulation that involves both positive and negative control. On top of the cascade, the two-component regulatory system FixLJ is activated under the microoxic conditions of the nodule. In addition, activity of the FixLJ system is inhibited by a specific anti-kinase protein, FixT. The physiological significance of this negative regulation by FixT was so far unknown.
Results
We have isolated by random Tn5 mutagenesis a S. meliloti mutant strain that escapes repression by FixT. Complementation test and DNA analysis revealed that inactivation of an asparagine synthetase-like gene was responsible for the phenotype of the mutant. This gene, that was named asnO, encodes a protein homologous to glutamine-dependent asparagine synthetases. The asnO gene did not appear to affect asparagine biosynthesis and may instead serve a regulatory function in S. meliloti. We provide evidence that asnO is active during symbiosis .
Conclusions
Isolation of the asnO mutant argues for the existence of a physiological regulation associated with fixT and makes it unlikely that fixT serves a mere homeostatic function in S. meliloti. Our data suggest that asnO might control activity of the FixT protein, in a way that remains to be elucidated. A proposed role for asnO might be to couple nitrogen fixation gene expression in S. meliloti to the nitrogen needs of the cells.
Background
Sinorhizobium meliloti forms N2-fixing nodules on the roots of alfalfa (Medicago sativa) and closely related plants. Expression of nitrogen fixation genes is under both positive and negative control. This regulation depends on a regulatory cascade, on top of which the two-component regulatory system FixLJ activates expression of nitrogen fixation genes in response to microoxic conditions [1], such as those that prevail inside the nodule [2]. Under microoxic conditions, the sensor histidine kinase FixL autophosphorylates and transfers its phosphate to the FixJ transcriptional regulator protein [3]. Phosphorylated FixJ then activates transcription of two intermediate regulatory genes, nifA and fixK, that both encode transcriptional regulators [3]. NifA mediates activation of nif genes involved in nitrogenase biosynthesis whereas FixK, a member of the Crp/Fnr family, activates expression of genes involved in the synthesis of a respiratory oxidase complex [4, 5]. fixK is also indirectly responsible for negative regulation of the cascade since it controls expression of a gene, fixT, that negatively affects expression of FixLJ dependent genes (see Figure 6). We have shown recently that the FixT protein negatively affects the expression of nifA and fixK by inhibiting phosphorylation of the sensor hemoprotein kinase FixL and, by consequence, phosphorylation of FixJ [6]. Whether FixT serves a mere homeostatic function in S. meliloti (the level of FixT protein feed-back controlling activity of the FixLJ system) or whether FixT allows integration of a physiological signal by the FixLJ system was so far unknown. We addressed this question by looking for S. meliloti mutants in which the FixT protein would not be active in repression.
Figure 6.
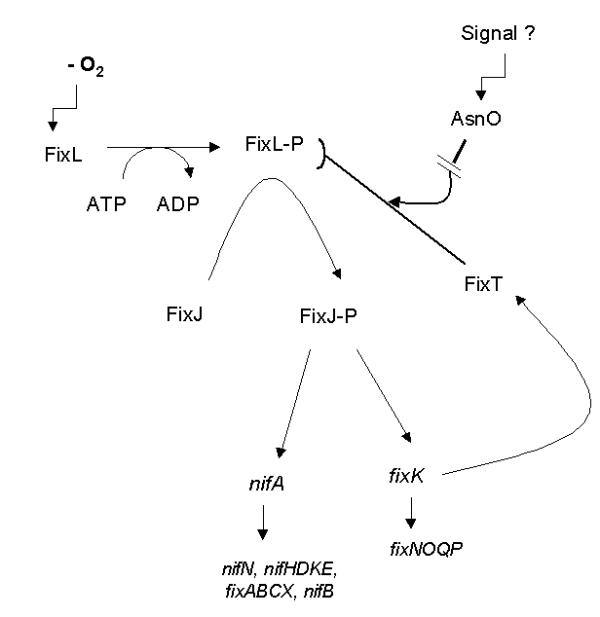
Updated model for the regulation of respiratory and nitrogen fixation gene expression in S. meliloti.
Here we report the isolation of a S. meliloti mutant strain that phenotypically escapes the repressor activity exerted by FixT. The mutation lies in a gene named asnO encoding a protein homologous to glutamine-amidotranferases. We discuss the significance of this finding with respect to the regulation of symbiotic nitrogen fixation.
Results and Discussion
Isolation of a S. meliloti mutant strain escaping repression by FixT
We previously observed that in a S. meliloti wild-type strain, constitutive expression of fixT, driven by the strong neomycin promoter of plasmid pMF10 (Table 1), resulted in strong inhibition of the expression of a fixK-lacZ reporter fusion (pMF457 plasmid; Table 1), thus leading to white colonies on X-gal containing plates. We used this observation to screen for S. meliloti mutants that would escape repression by FixT. After random Tn5 mutagenesis of a strain overexpressing fixT, we isolated blue colonies on X-gal containing plates. These putative mutant colonies were subsequently assayed for β-galactosidase activity in liquid cultures under microoxic conditions. Four independent mutants were isolated, that were characterized by an unrepressed level of expression of the fixK gene despite the constitutive expression of fixT (Figure 1A; compare lanes 2 and 3). Two of the mutants possessed a Tn5 insertion in the same gene. One of these two mutants, GMI401, was further characterized. Southern-Blot analysis of genomic DNA digested by different restriction enzymes, revealed a single Tn5 insertion in GMI401 (data not shown). Transduction experiments using the N3 phage, showed genetic linkage between the mutant phenotype and the Tn5 insertion since reintroduction of the pMF10 plasmid in the transduced strain confirmed the lack of repression of fixK by overexpressed fixT (Figure 1A; lanes 5 and 6). This excluded the possibility that a mutation on one of the plasmids, pMF457 or pMF10, could have been responsible for the phenotype observed.
Table 1.
Bacterial strains and plasmids used
| Material | Characteristics | References |
| Strains | Genotype | |
| S. meliloti | ||
| GMI211 | SmR Nod+ Fix+ | [20] |
| GMI401 | GMI211 asnO::Tn5 SmR NmR | This work |
| GMI5704 | GMI211 fixJ2.3::Tn5 SmR NmR BleoR | [18] |
| E. coli | ||
| ER | asnA31 asnB32 thi | [15] |
| MM294 | Pro-82 thi-1 hsdR17 supE44 endA1 | [21] |
| DH5 α | endA1 hsdR17 (rk-mk+) supE44 thi-1 recA1 gyrA relA1Δ (lacIZYA- | [29] |
| argF)U169 deoR (φ 80dlacΔ (lacZ)M15) | ||
| Plasmids | ||
| pMF457 | pGD926 (IncP broad host range vector) derivative carrying a fixK- | |
| lacZ fusion with a mutation in the fixT promoter. TcR | ||
| pCHK57 | pGMI41211 (Inc-P1 broad host range vector) derivative carrying a | [31] |
| nifA-lacZ fusion. TcR | ||
| pMF11 | Same as pMF10 but with fixT in reverse orientation. GmR | |
| pBH1 | pBBR1-MCS3 derivative, carrying an HindIII fragment containing | This work |
| the asnO gene and the SMb20482 orf. | ||
| pBasn2 | pBBR1-MCS3 derivative, expressing asnO under the control of the | This work |
| pLac promoter | ||
| pUC23 | pUC18 digested with EcoRI, carrying the S. meliloti EcoRI | This work |
| fragment containing the asnO region with the Tn5 insert. ApR, | ||
| KmR. | ||
| BAC37 | pBeloBAC11 containing a 100 kb S. meliloti genome fragment | [26] |
| carrying the asn-like region of pSymB. | ||
| pRK602 | PRK600::Tn5, Cmr, Nm-Kmr | [21] |
Figure 1.
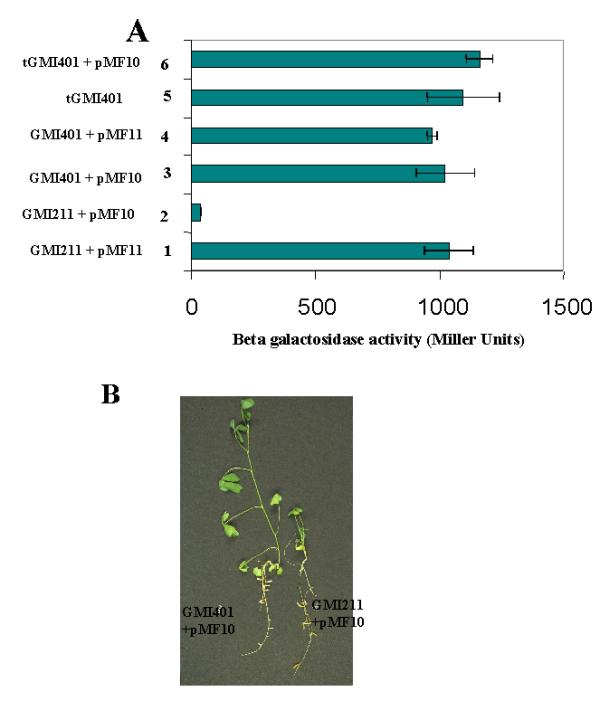
Characterization of the S. meliloti mutant strain (GMI401) escaping FixT repressor activity. Panel A : We monitored microoxic expression of a fixK-lacZ fusion carried by the reporter plasmid pMF457 in S. meliloti. pMF10 allows constitutive expression of fixT, pMF11 is a negative control (fixT cloned in the antisense orientation). 1 : wild-type strain GMI211(pMF457)(pMF11); 2 : wild-type strain GMI211(pMF457)(pMF10); 3 : mutant strain GMI401(pMF457)(pMF10); 4 : mutant strain GMI401 (pMF457) (pMF11); 5 : transductant strain GMI401(pMF457); 6 : transductant strain GMI401(pMF457)(pMF10). Panel B : In planta phenotype of the wild-type GMI211 (pMF10) and mutant strain GMI401 (pMF10). Medicago sativa seedlings were inoculated with the bacterial strains and grown for 3 weeks on medium lacking any nitrogen source.
In the GMI401 mutant strain, the level of fixK gene expression in the absence of pMF10 was the same as in the wild-type strain (Figure 1A; compare lane 1 to lane 4 and lane 5). This result demonstrated that activation of fixK gene expression by the FixLJ two-component system was not affected in the mutant strain, but rather that the phenotype was genuinely due to decreased repression by fixT. Western-Blot analysis using an antibody directed against the FixT protein indicated that the level of FixT protein was the same in the GMI211(pMF10) wild-type strain and in the GMI401(pMF10) mutant strain (data not shown).
Similar results have been obtained using a nifA-lacZ fusion (pCHK57 plasmid). The inhibition of nifA-lacZ expression by overexpressed fixT that was observed in a GMI211 wild-type strain was not observed anymore in the GMI401 mutant strain (data not shown).
Altogether, these results tend to indicate that, in the GMI401 mutant strain, the absence of repression of fixK and nifA by fixT was due to a reduced inhibition of the FixLJ system by the FixT protein.
The GMI401 mutant strain produced nitrogen fixing nodules on M. sativa, as did the GMI211 wild-type strain (data not shown). Whereas the GMI211 parent strain expressing fixT constitutively produced non N2-fixing nodules (Figure 1B), the GMI401 mutant strain was able to induce effective N2-fixing nodules on alfalfa (Medicago sativa cv. gemini) thus enabling the plants to grow in the absence of combined nitrogen. Hence, the gene that modulates repression by FixT activity ex planta is also active in planta.
The Tn5-insertion maps in an asparagine synthetase-like gene
The Tn5 insertion in GMI401 was positioned on the pSymB megaplasmid of S. meliloti (see Materials and Methods), whereas the fixLJ, fixT and fixK genes are located on pSymA megaplasmid. The genomic DNA flanking the Tn5 insertion was sequenced (Figure 2 and Materials and Methods). Prediction of coding regions around the region of interest was performed using the FrameD program [7]. As shown in Figure 2, this analysis revealed a putative open reading frame encompassing the Tn5 insertion site and another putative orf located just dowstream. On the basis of the sequence analysis, these two genes may belong to the same operon, and the Tn5 insertion may thus affect expression of both genes. During the annotation phase of the S. meliloti genome sequencing project (http://sequence.toulouse.inra.fr/meliloti.html) that was run concomitantly to this work, the upstream gene was named asnO, for its homology to the Bacillus subtilis asnO gene. We have thus adopted this annotation in this paper. The deduced amino sequence of the second orf, that was named SMb20482, is similar to an acetyl-transferase in the amino-terminal part of the protein and to a cyanophycin synthetase gene in the carboxy-terminal part of SMb20482.
Figure 2.
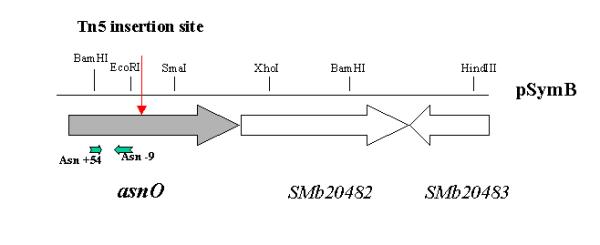
Genetic organisation of the asnO region. The genes are shown as thick arrows. The red small arrow indicates the position of the Tn5 insertion in the GMI401 mutant strain. The position and orientation of the specific primers are shown in small green arrows.
The deduced amino acid sequence of the gene carrying the Tn5 insertion is similar to that of glutamine-dependent asparagine synthetases of various bacteria including B. subtilis and Escherichia coli (see Figure 3). These proteins are members of the PurF family of glutamine-dependent amidotransferases. All PurF enzymes possess a conserved amino-terminal cysteine, which is essential for glutamine-dependent amidotransferase activity [8, 9]. This Cys2 residue is also conserved in the S. meliloti asnO product. The PurF-type amidotransferases possess 14 additional conserved residues in the amino-terminal glutamase domain [8, 9]. As shown by the alignment of Figure 3, all these residues are present in the asnO product of S. meliloti. We conclude that the AsnO protein belongs to the PurF family of glutamine amidotransferases. The carboxy-terminal domian of the proteins, carrying the synthetase activity, is less conserved.
Figure 3.
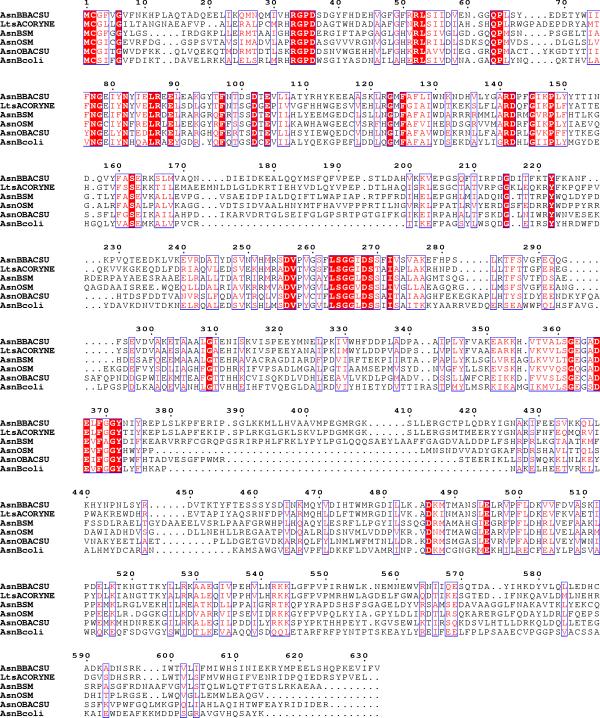
AsnO of S. meliloti is related to glutamine-dependent amido transferases. Alignment of amino acid sequences of AsnO and AsnB from S. meliloti, AsnO (swissprot accession number: Sp 005272) and AsnB (Sp P54420) from B. subtilis, AsnB from E. coli (Sp P22106), and LtsA from C. glutamicum (Sp BAA89484). The alignement was done by using ClustalW programme [31].
The asnO gene complements the mutant phenotype of GMI401
In order to perform complementation trials, we cloned the asnO gene in the pBBRI-MCS3 plasmid [10], a shuttle vector that replicates in both E. coli and S. meliloti. A DNA fragment containing the entire asnO coding region and its 5' flanking promoter region, was cloned downstream of the lac promoter of the plasmid pBBRI-MCS3. We introduced the corresponding pBasn2 plasmid in a strain expressing fixT constitutively and monitored expression of fixK under microoxic conditions by RT-PCR experiments. Data indicated complementation of the GMI401 mutant strain by the asnO transgene (Figure 4). The restoration of the wild-type phenotype (ie inhibition of fixK expression in the presence of pMF10), demonstrated the implication of the asnO gene in the characteristic phenotype observed in the GMI401 mutant strain. However, restoration of the wild-type phenotype was not complete, for a reason that we do not understand.
Figure 4.
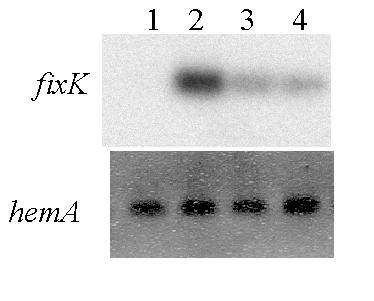
Complementation of the mutant phenotype by the asnO gene. RNAs isolated from free-living microoxic cultures (2% O2) of S. meliloti strains grown in M9 minimal medium were amplified by RT-PCR using specific primers. RT-PCR products were separated on agarose gels, blotted onto a nylon membrane and hybridized with the 32P labelled probe of the expected product. Upper panel : fixK gene (see Materials and Methods). Lower panel : hemA gene (control). Lane 1 = GMI211 (pMF10), Lane 2 = GMI401 (pMF10), Lanes 3 and 4 = GMI401 (pMF10) (pBasn2) of two independent transconjugants.
asnO inactivation in GMI401 does not lead to auxotrophy for asparagine
Because of the homology of the S. meliloti asnO gene product with known asparagine synthetases, we tested whether the asnO gene was involved in asparagine biosynthesis, by three complementary approaches (data not shown).
First, we observed that the mutant strain GMI401 and the isogenic GMI211 wild-type strain, grew at equal and similar rates in minimal medium with or without asparagine.
Second, we tested the ability of pBasn2 to restore prototrophy of a E. coli asparagine auxotroph, a double asnB/asnA mutant strain (Table 1). In asparagine-supplemented minimal medium, all strains grew at equal rates. However, in the absence of added asparagine, the S. meliloti asnO gene under the control of the plac promoter was unable to complement the asparagine auxotrophy of the asnA/asnB E. coli mutant strain.
Third, addition of asparagine at different concentrations in minimal medium, did not lead to complementation of the S. meliloti GMI401 mutant phenotype, i.e did not restore repression of fixK gene expression by overexpressed fixT. This observation suggested that the mutant phenotype is not dependent on the level of asparagine, and hence, that the asnO gene does not mediate asparagine synthesis.
The reactions catalyzed by asparagine synthetases involve two different family of proteins depending on whether glutamine or ammonia is used as a nitrogen source. Members of the AsnA family, that are found in procaryotes, only use ammonia as the amino group donor [11, 12]. The AsnB family, which is found in both procaryotes and eucaryotes, preferentially uses glutamine as nitrogen source [13, 14]. E. coli possesses an asnA and an asnB gene [15] whereas three asparagine synthetase genes, asnB, asnH and asnO, which all belong to the AsnB family, have been characterized in B. subtilis. No member of the AsnA family has been found in B. subtilis [16]. Complete genome analysis indicated that S. meliloti carries no asnA-like gene and two asnB-type genes (see http://sequence.toulouse.inra.fr/meliloti.html) one of which is asnO and a second gene that was named asnB. Sequence analysis is consistent with the possibility that the asnB gene of S. meliloti might be the ortholog of the biosynthetic asnB gene of E. coli and B. subtilis whereas the asnO gene mutated in GMI401 may play a regulatory function, in relationship with nitrogen fixation and microoxic respiration (Figure 3). Similarly, the asnO gene of B. subtilis has a regulatory role in sporulation [16], whereas the ltsA gene of Corynebacterium glutamicum is involved in cell wall formation [17].
asnO gene expression is induced in microoxic conditions
We have monitored by RT-PCR experiments expression of the asnO gene in a wild-type strain grown in minimal medium, in either oxic or microoxic conditions. The results showed that asnO gene expression was slightly enhanced in microoxic conditions as compared to oxic conditions (Figure 5). No effect of a fixJ mutation was observed on asnO expression using a GMI5704 fixJ mutant strain [18].
Figure 5.

asnO gene expression RT-PCR analysis of RNAs isolated from S. meliloti GMI211 wild-type strain (wt) or GMI5704 fixJ mutant strain (fixJ) grown in minimal medium M9 in either oxic (+) or microoxic (2% O2) conditions (-). RT-PCR were performed with either asnO or hemA specific primers and the products were separated on agarose gels, blotted on a nylon membrane and hybridized with the corresponding 32P labelled PCR product and analysed on a Phosphorimager.
Conclusions
FixT is an intriguing protein as it has not been described so far in any other bacterium besides S. meliloti. Furthermore its mode of action is original, as it has the capacity to block phosphorylation, and hence activity, of the FixL sensor histidine kinase. They are only a few examples of such anti-kinase proteins in the literature. Lastly, FixT primary sequence did not provide clues to its function. There is thus a great deal of interest in determining the biological role of fixT in S. meliloti.
The present work argues in favor of a physiological function associated with fixT, by showing that mutation of the asnO gene impairs repression by the FixT protein. This finding brings support to the previous suggestion that FixT may allow integration of an additional signal by the FixLJ two-component regulatory system whose activity is primarily regulated by oxygen (Figure 6). Multiple signal integration by a single two-component regulatory system is well documented for instance in B. subtilis [19].
Further work is required to elucidate the relationship between fixT, fixL and asnO. We propose as a working model that the absence of AsnO may result in an imbalance in the pool of a metabolite (e.g a substrate or a product of AsnO), that would affect the intrinsic repressing activity of FixT or, equally, the interaction between FixT and FixL. Identification of the reaction catalyzed by AsnO and further elucidation of the mode of action of the FixT protein should shed light to this model. Because glutamine, a likely by-product of nitrogen fixation in symbiotic rhizobia, is a predicted substrate of the AsnO protein, it is tempting to speculate that asnO and fixT may provide a link between the nitrogen status of bacteria -or of the plant cell- and nitrogen fixation activity and reducing power generation. Possibly, such a genetic device may connect the nitrogen needs of the plant to the nitrogen fixation activity of the microsymbiont.
Materials and Methods
Bacterial strains, plasmids and growth conditions
The bacterial strains and plasmids used in this study are listed in Table 1. S. meliloti GMI211, the symbiotically effective parent strain used in this study is a lac Smr, Nod+ Fix+ derivative of S meliloti RCR2011 [20]. Tn5 transposon mutagenesis of GMI211 was performed using the pRK602 plasmid (Table 1, [21]). In order to isolate random Tn5 insertions in the S. meliloti genome, we conjugated E. coli MM294(pRK602) donor strain with a S. meliloti GMI211(pMF10) recipient strains, and screened for streptomycin (100 μg/ml) and neomycin (100 μg/ml) resistant transconjugants.
S. meliloti strains were grown at 30°C in TY complex medium or in defined M9 medium [22] supplemented with 0.3 mM CaCl2, 1 mM MgSO4 and 2 μM biotin. Sterilised carbon sources were added at 20 mM final concentrations. Microoxic conditions were achieved as described by de Philip et al [23] (2% oxygen for 4 h). For testing induction of fixK-LacZ expression by microoxic conditions, S. meliloti strains were previously grown to OD600 = 0.3 in M9 medium. β-Galactosidase assays were performed as described by de Philip et al. [23].
Southern Blots
DNA techniques were carried out as described by Sambrook et al. [24]. S. meliloti genomic DNA was prepared as described by Chen and Kuo [25]. For Southern-Blot analysis, 1 μg of genomic DNA was digested at 37°C for 4 hours with the appropriate enzymes. DNA fragments were separated on a 0.8% agarose gel, transferred onto a nylon membrane (BiodyneA transfer membrane, Pall, East Hills, NY), and hybridized with a Tn5 32P-labelled probe.
asnO cloning and sequencing
A genomic fragment of S. meliloti GMI401 carrying the Tn5 insertion was cloned from a EcoRI-digested genome fragment into a pUC18 plasmid, resulting in the pUC23 plasmid (see Table 1). However, the presence of a EcoRI restriction site in the asnO gene (see Figure 2) did not permit to obtain the entire sequence of the gene from this construct. Therefore, a library of 96 BACs covering the S. meliloti genome [26], was screened to isolate the full-length asnO gene. PCR screening of the BAC library was performed using the oligonucleotide primers ASN+54 and ASN-9 shown in Figure 2. By this method, we could detect the presence of the asnO gene on BAC37, that was assigned to the pSymB megaplasmid [26]. Complete sequence of the asnO region was determined by a primer-walking approach on the BAC37 and pBH1 plasmid (see Table 1), which was obtained by subcloning a HindIII fragment of BAC37 in pBBR1-MCS3 [10]. Sequencing was performed on a ABI373 automated sequencer (ABI, Columbia, MD), using the ABI PRISM Dye terminator cycle sequencing ready kit (Perkin-Elmer, Oak, Brook, IL).
Database searches
Putative open reading frame were predicted using the FrameD program [7] used for S. meliloti whole genome analysis (http://sequence.toulouse.inra.fr/meliloti.html). Protein analysis was done with the NCBI web page using the BLAST2 package program [27] against the NCBI-nr and Swiss-Prot databases. Homologous domains searches were driven using the ProDom database [28].
Plant methods
Medicago sativa cv. Gemini seedlings were aseptically grown on agar slants made up with nitrogen-free Fahraeus medium. Three-day-old plants were inoculated with the different S. meliloti strains and grown for 3 weeks before observation.
RNA preparation
The bacteria from a 25 ml culture at 0.4 OD600nm were harvested and RNA prepared with the Qiagen RNeasy kit as described by the manufacturer. DNA was eliminated by addition of 7.5 Units of FPLC-Pure RNase-free DNaseI (Amersham-Pharmacia Biotech). RNA was further extracted with phenol-chloroform and then precipitated with ethanol. After washing with 70% ethanol, the pellet was resuspended in nuclease-free water. RNA was quantified by absorbance measurements at 260 nm. Absence of DNA contamination was verified by PCR amplification.
RT-PCR analysis
RT-PCR reactions were performed according to the manufacturer, using the SuperScript™ One-Step RT-PCR System (GibcoBRL).
The following oligonucleotides were used for reverse transcription of the messenger RNA and amplification of the product to evaluate gene expression :
For the hemA gene: hemAr (reverse primer) : 5'-GTCGATCGCGTTCTT-3'; hemAf (foward primer) : 5'-TGGATGGGCTGCATCA-3'
For the asnO gene: RTA1 (reverse primer) : 5'-TGCGTATTCTCGACCTG-3'; RTA2 (foward primer) : 5'-TCGCGAAAATTGTAGATG-3'
For the fixK gene: KPR (reverse primer) : 5'- CCGATTACCAGAAGATGC-3'; KPF (foward primer) : 5'-TATCTACCGCCTCCTTTC-3'
RT-PCR products were electrophoresed on a 2% agarose gel, blotted onto a nylon membrane and probed with a 32P-labeled DNA probe prepared from the gene of interest. Washing was done with 0.1XSSC, 0.1%SDS at 42°C during 30 min.
Acknowledgments
Acknowledgements
We thank Dr T. Hirasawa for generous gift of E. coli strain ER, Dr Delphine Capela for access to the S. meliloti BAC genomic library, Laurent Sauviac for automatic cycle sequencing, Isabelle Gairin for informatic technical assistance, Jérôme Gouzy and Frédéric Ampe for help in sequence analysis. We are grateful to Dr Julie Cullimore and Dr Eliane Meilhoc for critical reading of the manuscript. HB and part of this work were supported by a grant from the European Union in the frame of the BIOTECH programme (FIXNET, BIOT4-CT97-2319).
Contributor Information
Hélène Bergès, Email: hberges@toulouse.inra.fr.
Claire Checroun, Email: checroun@ibcg.biotoul.fr.
Sébastien Guiral, Email: guiral@ibcg.biotoul.fr.
Anne-Marie Garnerone, Email: garneron@toulouse.inra.fr.
Pierre Boistard, Email: boistard@toulouse.inra.fr.
Jacques Batut, Email: jbatut@toulouse.inra.fr.
References
- David M, Daveran ML, Batut J, Dedieu A, Domergue O, Ghai J, Hertig C, Boistard P, Kahn D. Cascade Regulation of nif Gene Expression in Rhizobium meliloti. Cell. 1988;54:671–683. doi: 10.1016/s0092-8674(88)80012-6. [DOI] [PubMed] [Google Scholar]
- Soupene E, Foussard M, Boistard P, Truchet G, Batut J. Oxygen as a key developmental regulator of Rhizobium meliloti N2- fixation gene expression within the alfalfa root nodule. Proc Natl Acad Sci USA. 1995;92:3759–3763. doi: 10.1073/pnas.92.9.3759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilles-Gonzalez MA, Ditta GS, Helinski DR. A haemoprotein with kinase activity encoded by the oxygen sensor of Rhizobium meliloti. Nature. 1991;350:170–2. doi: 10.1038/350170a0. [DOI] [PubMed] [Google Scholar]
- Reyrat JM, David M, Blonski C, Boistard P, Batut J. Oxygen regulated in vitro transcription of Rhizobium meliloti nifA and fixK genes. J Bacteriol. 1993;175:6687–6872. doi: 10.1128/jb.175.21.6867-6872.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Batut J, Daveran-Mingot ML, David M, Jacobs J, Garnerone AM, Kahn D. fixK, a gene homologous with fnr and crp from Escherichia coli, regulates nitrogen fixation genes both positively and negatively in Rhizobium meliloti. EMBO J. 1989;8:1279–1286. doi: 10.1002/j.1460-2075.1989.tb03502.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foussard M, Garnerone AM, Ni F, Soupene E, Boistard P, Batut J. Negative autoregulation of the Rhizobium meliloti fixK gene is indirect and requires a newly identified regulator, FixT. Mol Microbiol. 1997;25:27–37. doi: 10.1046/j.1365-2958.1997.4501814.x. [DOI] [PubMed] [Google Scholar]
- Garnerone AM, Cabanes D, Foussard M, Boistard P, Batut J. Inhibition of the FixL sensor kinase by te FixT protein in Sinorhizobium meliloti. J Biol Chem. 1999;274:32500–32506. doi: 10.1074/jbc.274.45.32500. [DOI] [PubMed] [Google Scholar]
- Schiex T, Thébault P, Kahn D. Recherche des gènes et des erreurs de séquençage dans les génomes bactériens GC-riches (et autres...) Proc of JOBIM', Montpellier France. 2000:321–328. [Google Scholar]
- Boehlein SK, Richards NG, Walworth ES, Schuster SM. Arginine 30 and asparagine 74 have functional roles in the glutamine dependent activities of Escherichia coli asparagine synthetase B. J Biol Chem. 1994;269:26789–95. [PubMed] [Google Scholar]
- Boehlein SK, Richards NG, Schuster SM. Glutamine-dependent nitrogen transfer in Escherichia coli asparagine synthetase B. Searching for the catalytic triad. J Biol Chem. 1994;269:7450–7. [PubMed] [Google Scholar]
- Kovach ME, Elzer PH, Hill DS, Robertson GT, Farris MA, RoopII RM, Peterson KM. Four new derivatives of the broad-host-range cloning vector pBBR1-MCS, carrying different antibiotic-resistance cassettes. Gene. 1995;1666:175–176. doi: 10.1016/0378-1119(95)00584-1. [DOI] [PubMed] [Google Scholar]
- Humbert R, Simoni RD. Genetic and biochemical studies demonstrating a second gene coding for asparagine synthetase in Escherichia coli. J Bacteriol. 1980;142:212–220. doi: 10.1128/jb.142.1.212-220.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reitzer LJ, Magasanik B. Asparagine synthetase of Klebsiella aerogenes : properties and regulation of synthesis. J Bacteriol. 1982;151:1299–1313. doi: 10.1128/jb.151.3.1299-1313.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes CA, Beard HS, Matthews BF. Molecular cloning and expression of two cDNAS encoding asparagine synthetase in soybean. Plant Mol Biol. 1997;33:301–311. doi: 10.1023/a:1005784202450. [DOI] [PubMed] [Google Scholar]
- Scofield MA, Lewis WS, Schuster SM. Nucleotide sequence of Escherichia coli asnB and deduced amino acid sequence of asparagine synthetase B. J Biol Chem. 1990;265:12895–902. [PubMed] [Google Scholar]
- Cedar H, Schwartz JH. The asparagine synthetase of Escherichia coli. Biosynthetic role of the enzyme, purification, and characterization of the reaction products. J Biol Chem. 1963;244:4112–4121. [PubMed] [Google Scholar]
- Yoshida KI, Fujita Y, Ehrlich SD. Three asparagine synthetase genes of Bacillus subtilis. J Bacteriol. 1999;181:6081–6091. doi: 10.1128/jb.181.19.6081-6091.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirasawa T, Wachi M, Nagai K. A mutation in the Corynebacterium glutamicum ltsA gene causes susceptibility to lysozyme, temperature-sensitive growth, and L-glutamate production. J Bacteriol. 2000;182:2696–26701. doi: 10.1128/jb.182.10.2696-2701.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- David M, Daveran ML, Batut J, Dedieu A, Domergue O, Ghai J, Hertig C, Boistard P, Kahn D. Cascade regulation of nif gene expression in Rhizobium meliloti. Cell. 1988;54:671–683. doi: 10.1016/s0092-8674(88)80012-6. [DOI] [PubMed] [Google Scholar]
- Perego M, Hanstein C, Welsh K, Djavakhishvili T, Glaser P, Hoch JA. Multiple protein-aspartate phosphatases provide a mechanism for the integration of diverse signals in the control of development in B. subtilis. Cell. 1994;79:1047–1055. doi: 10.1016/0092-8674(94)90035-3. [DOI] [PubMed] [Google Scholar]
- Niel C, Guillaume B, Bechet M. Mise en évidence de deux enzymes présentant une activité β-galactosidase chez Rhizobium meliloti. Can J Microbiol. 1977;23:1178–1181. [PubMed] [Google Scholar]
- Finan TM, Hirsch AM, Leigh JA, Johansen E, Kuldau GA, Deegan S, Walker GC, Signer ER. Symbiotic mutants of Rhizobium meliloti that uncouple plant from bacterial differentiation. Cell. 1985; 40:869–877. doi: 10.1016/0092-8674(85)90346-0. [DOI] [PubMed] [Google Scholar]
- Miller J.H. Experiments in molecular genetics. Cold Spring Harbor, New York. 1972.
- de Philip P, Batut J, Boistard P. FixL is an oxygen sensor and regulates R. meliloti nifA and fixK genes differently in Escherichia coli. J Bacteriol. 1990;172:4255–62. doi: 10.1128/jb.172.8.4255-4262.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J, Fristsch EF, Maniatis T. Molecular cloning. A Laboratory Manual Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. 1989.
- Chen WP, Kuo TT. A simple and rapid method for the preparation of Gram-negative bacterial genomic DNA. Nucleic Acids Res. 1993;21:2260–2260. doi: 10.1093/nar/21.9.2260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Capela D, BarloyHubler F, Gatius MT, Gouzy J, Galibert F. A high-density physical map of Sinorhizobium meliloti 1021 chromosome derived from bacterial artificial chromosome library. Proc Nat Acad Sci Usa. 1999;96:9357–9362. doi: 10.1073/pnas.96.16.9357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corpet F, Gouzy J, Kahn D. Recent improvements of the ProDom database of protein domain families. Nucleic Acids Res. 1999;27:263–267. doi: 10.1093/nar/27.1.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodcock DM, Crowther PJ, Doherty J, Jefferson S, DeCruz E, Noyer-Weidner M, Smith SS, Michael MZ, Graham MW. Quantitative evaluation of Escherichia coli host strains for tolerance to cytosine methylation in plasmid and phage recombinants. Nucleic Acids Res. 1989;17:3469–78. doi: 10.1093/nar/17.9.3469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson JD, Higgins1 DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Research. 1994;22:4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ditta G, Schmidhauser T, Yakobson E, Lu P, Liang XW, Finlay DR, Guiney D, Helinski DR. Plasmids related to the broad host range vector, pRK290, useful for gene cloning and for monitoring gene expression. Plasmid. 1985;13:149–153. doi: 10.1016/0147-619x(85)90068-x. [DOI] [PubMed] [Google Scholar]