

Abstract
In earlier studies, the assimilation of selenate by plants appeared to be limited by its reduction, a step that is thought to be mediated by ATP sulfurylase. Here, the Arabidopsis APS1 gene, encoding a plastidic ATP sulfurylase, was constitutively overexpressed in Indian mustard (Brassica juncea). Compared with that in untransformed plants, the ATP sulfurylase activity was 2- to 2.5-fold higher in shoots and roots of transgenic seedlings, and 1.5- to 2-fold higher in shoots but not roots of selenate-supplied mature ATP-sulfurylase-overexpressing (APS) plants. The APS plants showed increased selenate reduction: x-ray absorption spectroscopy showed that root and shoot tissues of mature APS plants contained mostly organic Se (possibly selenomethionine), whereas wild-type plants accumulated selenate. The APS plants were not able to reduce selenate when shoots were removed immediately before selenate was supplied. In addition, Se accumulation in APS plants was 2- to 3-fold higher in shoots and 1.5-fold higher in roots compared with wild-type plants, and Se tolerance was higher in both seedlings and mature APS plants. These studies show that ATP sulfurylase not only mediates selenate reduction in plants, but is also rate limiting for selenate uptake and assimilation.
Se is an essential trace element for animals and bacteria, but it is also toxic at higher concentrations (Wilber, 1980). Se is naturally present in soils derived from shale (up to approximately 100 ppm [Wilber, 1980]), and when these soils are irrigated, selenate (SeO42−) leaches into the drainage water (McNeal and Balisteri, 1989). In addition, selenite (SeO32−) is a common contaminant in oil-refinery wastewater (Hansen et al., 1998). As a result, Se has become a serious environmental pollutant in the western United States and other areas worldwide, causing death and deformities in wildlife (Ohlendorf et al., 1986).
A promising new technology for the remediation of Se-polluted water and soil is phytoremediation. Plants can take up Se from water, soil, or sediment, accumulate it in their tissues (Wu et al., 1988; Banuelos et al., 1992, 1997; Terry et al., 1992), and volatilize it (Lewis et al., 1966; Zieve and Peterson, 1984; Velinsky and Cutter, 1991; Duckart et al., 1992; Terry et al., 1992, Terry and Zayed, 1994, 1998; Zayed and Terry, 1994). Volatile forms of Se such as dimethylselenide have been reported to be 500 to 600 times less toxic than inorganic forms (McConnell and Portman, 1952; Ganther et al., 1966; Wilber, 1980). For Se phytoremediation, terrestrial plants can be grown in Se-contaminated soils, and aquatic plants can be grown in constructed wetlands used for the treatment of Se-contaminated wastewater. Both strategies have already been shown to be quite efficient (Banuelos and Meek, 1990; Banuelos et al., 1995; Hansen et al., 1998; Terry and Zayed, 1998). Indian mustard (Brassica juncea) has proved to be a particularly suitable species for Se remediation, with high rates of Se accumulation and volatilization, a fast growth rate, and high productivity (Banuelos and Schrale, 1989; Banuelos and Meek, 1990; Terry et al., 1992; Wu et al., 1996).
Genetic engineering offers a powerful new means to improve the capacity of plants to remedy environmental pollutants. Once it is known which pathway is involved, the overexpression of rate-limiting enzymes may accelerate the flux through the entire pathway. The uptake and assimilation of selenate and sulfate are generally assumed to follow the same pathway (Ng and Anderson, 1979; Zayed and Terry, 1992; Anderson, 1993; Lauchli, 1993). Sulfate is actively transported into plant cells by sulfate permease (Leggett and Epstein, 1956). In Brassica napus a single, low-affinity sulfate transporter was detected, which was induced by S starvation and repressed by sulfate (Hawkesford et al., 1993). Sulfate uptake by this transporter was competitively inhibited by selenate. Similar results were found for sulfate permease from the legume Stylosanthes hamata (Smith et al., 1995). Part of the sulfate entering the cell is covalently linked to many different secondary metabolites and sulfolipids (Leustek, 1996), and part is reduced and assimilated. Sulfate reduction and assimilation occur primarily in leaves, and most enzymes are localized within plastids (Schwenn, 1994; Leustek, 1996). For reduction, sulfate is first activated by ATP sulfurylase to form adenosine phosphosulfate, which is subsequently reduced to free sulfite by adenosine phosphosulfate reductase (Setya et al., 1996). ATP sulfurylase, similar to sulfate permease, was induced by S starvation and repressed by feeding sulfate or reduced forms of S (Chen and Leustek, 1995; Logan et al., 1996).
Several in vitro studies have proposed that ATP sulfurylase mediates the reduction of selenate as well as sulfate in plants (Shaw and Anderson, 1972; Dilworth and Bandurski, 1977; Burnell, 1981). ATP sulfurylase is likely to be the pivotal rate-limiting enzyme controlling the pathway of S assimilation (Leustek, 1996), because it has a high substrate/product ratio, a Km for sulfate and ATP in the millimolar range, and it is subject to a powerful inhibition by its product adenosine phosphosulfate (Ki = 0.04 μm [Schwenn, 1994]). Similarly, the reduction of selenate was proposed to be rate limiting for the selenate assimilation pathway (de Souza et al., 1998; Zayed et al., 1998): when plants of several species, including Indian mustard, were supplied with selenate, they accumulated selenate, whereas when they were supplied with selenite, they accumulated an organic Se compound resembling SeMet.
Thus, ATP sulfurylase may be rate limiting for Se assimilation, and overexpression of this enzyme may increase the flux of the pathway. To test this hypothesis, the APS1 gene from Arabidopsis encoding a plastid-localized ATP sulfurylase (Leustek et al., 1994) was overexpressed in Indian mustard. The transgenic APS plants were compared with untransformed Indian mustard plants with respect to their selenate reduction, Se accumulation, and Se tolerance.
MATERIALS AND METHODS
Plant Transformation and Characterization
Indian mustard (Brassica juncea) seeds (accession no. 173874) were obtained from the North Central Regional Plant Introduction Station (Ames, IA). The DNA construct used to transform the plants contained the Arabidopsis APS1 cDNA, including its own chloroplast transit sequence, under the control of the cauliflower mosaic virus 35S promoter (Chen et al., 1997). A 1490-bp XhoI-Psp1406I fragment from pYES-APS1 (Leustek et al., 1994) was cloned into the BamHI-AccI sites of pBluescript SK(+). The polylinker restriction sites from pBluescript, SpeI and KpnI, were then used to clone APS1 into pFF20, which is a modified form of pFF19 (Timmermans et al., 1990). pFF19 was modified by replacing the HindIII site with a SalI site and by eliminating the SalI site from the polylinker of the plasmid. The expression cassette from pFF20 carrying APS1 was cloned as an EcoRI-SalI fragment into the EcoRI and SalI sites of pBI101.
This construct was used to transform Agrobacterium tumefaciens strain C53C1 as follows. All in vitro plant tissue cultures were grown at 25°C under continuous light. For transformation, Indian mustard hypocotyl segments were isolated from 3-d-old axenically grown seedlings (200–300 seedlings per transformation). The segments were immersed for 1 h in a suspension of the APS1-containing A. tumefaciens strain (A600 = 0.6, suspended in Murashige and Skoog medium); the bacteria were previously grown for 3 d at 28°C in liquid Luria-Bertani medium in the presence of 200 μm 3,5-dimethoxy-4-hydroxy-acetophenone (Fluka). After immersion in the bacteria suspension, the hypocotyls were blotted dry and transferred to modified Murashige and Skoog medium containing Murashige and Skoog salts and vitamins (Sigma, M5519), 4 g L−1 agarose, 10 g L−1 Suc, Glc, and mannitol, 200 μm 3,5-dimethoxy-4-hydroxy-acetophenone, 2 mg L−1 6-benzylaminopurine, and 0.1 mg L−1 naphthalene acetic acid. After 2 d of co-cultivation the hypocotyls were washed for 45 min in standard liquid Murashige and Skoog medium, blotted dry, and transferred to medium containing Murashige and Skoog salts and vitamins, 4 g L−1 agarose, 10 g L−1 Suc, Glc, and mannitol, 200 mg L−1 cefotaxime, 100 mg L−1 vancomycin, 20 mg L−1 kanamycin, 2 mg L−1 6-benzylaminopurine, 0.1 mg L−1 naphthalene acetic acid, and 30 μm AgNO3. After 11 d the hypocotyls were transferred to the same medium containing 10% coconut water (Sigma). Established shoots were transferred to standard Murashige and Skoog medium containing 30 g L−1 Suc, 100 mg L−1 cefotaxime, and 1 mg L−1 indole butyric acid to induce root formation.
PCR was used to identify APS transgenic lines among the kanamycin-resistant lines obtained. The PCR primers used were as follows: the forward primer was directed against the 35S promoter, with the sequence 5′ CCT TCG CAA GAC CCT TCC TC 3′. The reverse primer was directed against the APS gene and had the sequence 5′ CCG GAT CGA GAA CAC CAT CC 3′.
Total RNA was isolated from 7-d-old seedling shoots using the RNeasy Plant Mini Kit, according to the manufacturer's instructions (Qiagen, Chatsworth, CA). RNA electrophoresis, northern-blot hybridization, and washing of blots were performed as described by Hwang and Herrin (1994). The RNA blots were stained with methylene blue to ensure equal loading and transfer (Herrin and Schmidt, 1988). The APS1 DNA probe was generated by PCR using the primers described above. The PCR product was purified from the agarose gel and labeled with [32P]dCTP using random priming (Feinberg and Vogelstein, 1983).
Leaf and root samples for ATP sulfurylase enzyme analysis were collected from 9-d-old seedlings and from 6-week-old mature plants, all grown under greenhouse conditions. The samples, which consisted of total shoots/roots from seedlings or pooled samples from all leaves of a plant, were immediately stored on dry ice, ground in liquid nitrogen, and extracted with 1 mL g−1 fresh weight of a buffer containing 50 mm Tris, pH 8.0, 20% glycerol, 2 mm EDTA, and 0.1 mm PMSF. ATP sulfurylase enzyme activity was assayed in the reverse reaction, according to the method of Renosto et al. (1991).
Se Tolerance and Accumulation Experiments
To determine the Se tolerance of seedlings, T2 seeds from APS plants and wild-type Indian mustard seeds were sterilized by rinsing in 96% ethanol for 30 s, then in 0.65% hypochlorite solution for 30 min, and subsequently in sterile deionized water for 5 × 10 min, all on a rocking platform. Fifty sterilized seeds were sown in a grid pattern in Magenta boxes (Sigma) on one-half-strength Murashige and Skoog medium with 10 g L−1 Suc and 5 g L−1 Phytagar (Sigma), with or without added selenate (400 μm). After 7 d at 25°C under continuous light, individual seedlings were harvested, washed, and weighed, and the root length was measured.
For analysis of Se accumulation and tolerance in mature plants, APS and wild-type Indian mustard plants were grown in 4-inch pots containing coarse sand. The pots were maintained in a greenhouse with a controlled temperature (24°C) and a short-day (9 h) photoperiod to prevent them from flowering. The plants were watered twice a day, once with tap water and once with one-half-strength Hoagland solution (Hoagland and Arnon, 1938).
One week before the Se treatment, when the plants were 4 to 6 weeks old (for exact age, see Results), the plants were gently washed in water to remove the sand that had adhered to the roots and transferred into plastic boxes containing 3.5 L of aerated hydroponic solution (one-eighth-strength Hoagland solution). After 1 week in hydroponic conditions the nutrient solution was replaced by fresh solution containing various concentrations of Se. After 8 d of Se treatment, the plants were harvested and weighed. For elemental analysis, the plants were thoroughly washed in running deionized water to remove any Se that was bound to the outside of the roots, dried at 70°C, and the roots and shoots were ground separately using a mortar and pestle.
Biochemical Analysis of Transgenic Plants
For elemental analysis, powdered plant tissues (100-mg [dry weight] samples) were acid digested according to the method of Martin (1975). Se concentrations were analyzed in the acid digests using atomic absorption spectroscopy in combination with hydride generation (Mikkelsen, 1987). S concentrations in the acid digests were analyzed by inductively coupled plasma atomic emission spectroscopy according to the method of Fassel (1978).
The nonprotein thiol content of plant extracts was measured photospectrometrically according to the method of Galli et al. (1996). Extracts were prepared from 100-mg homogenized shoot samples by adding 300 μL of a solution containing 1 m NaOH and 1 mg L−1 NaBH4. The homogenate was centrifuged at 13,000g for 3 min at 4°C. Three hundred microliters of the supernatant was acidified by addition of 50 μL of 37% HCl, and 20 μL of this solution was added to 1 mL of 5,5′-dithiobis(2-nitrobenzoic acid) (Ellman's reagent [Ellman, 1959]), and the absorption was measured at 412 nm.
Total glutathione was measured according to a modification of the method described by Hermsen et al. (1997). Plant samples were ground in liquid nitrogen, and 100 mg of plant tissue was extracted with 0.3 mL of a solution containing 0.1 m HCl and 1 mm EDTA. One hundred fifty microliters of extract was then mixed with 300 μL of 0.1 m phosphate buffer (pH 8.0) containing 2.4 mm dithioerythritol and 45 μL of 0.28 m NaOH. This mixture was incubated for 1 h at room temperature. After 1 min of centrifugation 400 μL of supernatant was transferred to a new tube and 533 μL of phosphate buffer (pH 6.2) was added, followed by 26.7 μL of 1-chloro-2,4-dinitrobenzene. The A340 of this solution was set at 0, after which 0.72 unit of glutathione S-transferase was added and the change in A340 was measured continuously for 4 min. Statistical analyses were performed using the JMP IN statistical package (SAS Institute, Cary, NC).
XAS Analysis
Shoot and root tissues were collected from 5-week-old Indian mustard wild-type and APS plants supplied with 20 μm selenate for 8 d. The samples were frozen in liquid nitrogen, ground to a fine texture, and stored at −80°C. Comprehensive XAS analysis of frozen plant tissues was completed at the Stanford Synchrotron Radiation Laboratory on beam line 4-1. The electron energy was 3.0 GeV, with a current of approximately 50 to 100 mA. X-rays were monochromatized with a Si (111) double-crystal spectrometer, detuned 50% for harmonic rejection, with a 1-mm entrance slit that produced a beam of approximately 1 eV bandwidth at the Se K-edge. Frozen tissues were placed in a sample chamber at a 45° angle to the x-ray beam. Fluorescent x-ray spectra of Se in plant tissues and model Se reference compounds were collected with a series of replicate scans. The energy positions of all spectra were calibrated against a Se reference foil.
RESULTS
Production and Characterization of Transgenic APS Plants
Four kanamycin-resistant Indian mustard lines, designated APS1, APS8, APS9, and APS11, were obtained after transformation with the APS1 construct. When PCR was performed on these lines using primers directed against the 35S promoter (forward) and the Arabidopsis APS1 gene (reverse), the lines APS1, APS8, and APS9 showed PCR products of the expected size, whereas APS11 and wild-type plants did not (results not shown). Therefore, further experiments were performed with lines APS1, APS8, and APS9. None of these APS lines showed any phenotypic differences compared with untransformed Indian mustard plants.
To analyze the expression levels of the newly introduced APS1 gene in the transgenic plants, northern blotting was performed on RNA isolated from shoots of 7-d-old seedlings using the 35S/APS1 PCR product as a probe. One band of the expected size was observed in shoot samples from APS1, APS8, and APS9 seedlings; wild type (WT) did not give any signal (Fig. 1). The APS1 mRNA transcript levels in shoots of the APS1 and APS8 seedlings were similar, whereas APS9 seedlings appeared to have a somewhat lower expression level.
Figure 1.
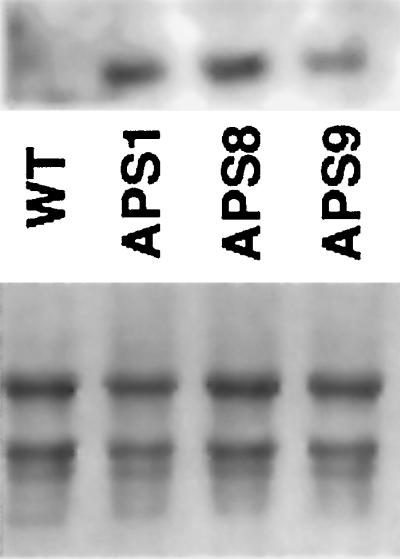
APS1 mRNA levels in shoot tissue of wild-type (WT) and APS1, APS8, and APS9 Indian mustard seedlings (7 d old). The mRNA data were obtained from pooled seedling samples (n = 50). Top, Northern blot using the 35S/APS1 PCR product as a probe. Bottom, Total RNA staining, showing equal RNA loading.
The ATP-sulfurylase enzyme activity was measured in leaf and root tissues of 9-d-old seedlings and from 6-week-old mature plants. The APS seedlings showed 1.7- to 2.7-fold higher ATP-sulfurylase activity in their shoots compared with wild-type seedlings, and 2.1- to 2.4-fold higher activity in their roots (P < 0.01; Fig. 2A). In mature APS plants, the leaf ATP sulfurylase activity levels were 2-fold higher than wild type in APS8 and APS9 plants (P < 0.01; Fig. 2B), and 1.5-fold higher in APS1 leaves (not significant); in roots of APS plants the enzyme activity levels were up to 30% higher than in wild-type roots, but these differences were not significant.
Figure 2.

ATP-sulfurylase activity in shoots and roots of 9-d-old seedlings (A) and 6-week-old plants (B) of wild-type (WT) and APS1, APS8, and APS9 Indian mustard. Values shown are the average and se of three samples, each representing 25 seedlings (A) or five mature plants (B).
APS Plants Show Increased Se Tolerance
When grown on agar medium containing 400 μm selenate, both the APS seedlings and the wild-type seedlings showed a dramatic reduction in growth, but this effect was less pronounced in the APS seedlings. After 7 d the APS seedlings had longer roots (Fig. 3A) and greater biomass (Fig. 3B). Root length is considered to be a sensitive and reliable parameter for trace-element tolerance (Murphy and Taiz, 1995). Se-treated APS seedlings of all three transgenic lines had about 50% longer roots than wild-type seedlings (P < 0.001). The fresh weights of Se-treated APS8 and APS9 seedlings were 29% and 43% higher, respectively, than those of wild-type seedlings (P < 0.001). For unknown reasons the APS1 seedlings did not attain higher fresh weights than the wild-type plants in this experiment, despite their better root growth. In the absence of Se there were no differences in seedling growth among the plant lines; the average root length and fresh weight of all control seedlings combined were 94.8 ± 1.3 mm and 146.1 ± 6.0 mg, respectively.
Figure 3.

Root length (A) and fresh weight (B) of wild-type (WT) and APS1, APS8, and APS9 Indian mustard seedlings grown for 7 d on agar medium containing 400 μm selenate. Values shown are the average and se of 50 seedlings.
Mature APS plants also showed increased Se tolerance when 4-week-old APS8 and wild-type plants were exposed to 50 μm selenate for 8 d. The wild-type plants were severely affected and nearly died under these conditions, whereas the APS8 plants were much less affected (Fig. 4). Under control conditions there was no difference in growth (not shown).
Figure 4.

Phenotypes of 4-week-old wild-type (WT) and APS8 Indian mustard plants after exposure to 50 μm selenate for 8 d. There were no visible differences between untreated WT and APS8 plants.
APS Plants Show Increased Se and S Accumulation
In the APS8 plants treated with 50 μm selenate, the shoot Se concentrations were 2-fold higher than in the wild-type plants (P < 0.01; Fig. 5). The root Se concentrations were 26% higher in APS8 plants than in wild-type plants (not significant; Table I).
Figure 5.

Shoot-tissue concentrations of Se (top) and S (bottom) in wild-type (WT) and APS8 Indian mustard plants supplied for 8 d with 20 μm selenate (left) or 50 μm selenate (right). At the start of the experiment, the plants treated with 20 μm selenate were 6 weeks old, and the plants treated with 50 μm selenate were 4 weeks old. Values shown are the average and se of eight replicates. The root Se and S concentrations are shown in Table I.
Table I.
Root-tissue concentrations of Se and S in wild-type and APS8 Indian mustard plants treated with two different concentrations of selenate
| 20 μm
|
50
μm
|
|||
|---|---|---|---|---|
| Plant | Se | S | Se | S |
| mg g−1 dry wt | ||||
| Wild type | 0.35 ± 0.02 | 3.84 ± 0.26 | 0.87 ± 0.13 | 4.35 ± 0.20 |
| APS8 | 0.62 ± 0.16 | 4.05 ± 0.35 | 1.10 ± 0.17 | 4.31 ± 0.46 |
Values shown are the average and se of eight replicate plants. The corresponding shoot data are shown in Figure 5.
Similar results were obtained when 6-week-old APS8 and wild-type plants were treated with 20 μm selenate for 8 d. The APS8 plants showed 2-fold higher shoot Se concentrations than wild-type plants (P < 0.01; Fig. 5), and 1.8-fold higher root Se concentrations (P = 0.10; Table I). At this Se concentration the plants did not show any visible symptoms of stress. When supplied with 20 μm selenite instead of selenate, the APS8 and APS9 plants showed no significant differences in Se accumulation compared with the wild-type plants (data not shown), confirming the idea that ATP sulfurylase is involved in the assimilation of selenate but not selenite.
To analyze the effect of ATP sulfurylase overexpression on S accumulation, the tissue S levels were measured in the selenate-treated plants. The APS8 plants treated with 50 or 20 μm selenate had about 1.5-fold higher S concentrations in their shoot tissues than comparable wild-type plants (P < 0.05; Fig. 5). No significant differences were found between S levels in root tissues of APS and wild-type plants (Table I).
The concentration-dependent kinetics of Se accumulation were studied by treating 5-week-old APS8, APS9, and wild-type plants with 5, 10, 20, or 40 μm selenate for 8 d, followed by the measurement of shoot and root Se concentrations. Both APS lines showed higher Se concentrations in their shoot tissues than wild-type plants at all concentrations tested (Fig. 6) with one exception: there was no difference between APS9 and wild type at 5 μm selenate. The difference in shoot Se concentration was most pronounced at the higher external selenate concentrations used; when treated with 20 to 40 μm selenate, the APS plants had 2- to 3-fold higher Se concentrations in their shoots than wild-type plants. The root-tissue Se levels in APS plants treated with 40 μm were 1.5-fold higher than those of wild-type plants (Table II ). The S levels were again higher in shoots of the APS plants than in wild-type shoots, both in Se-treated and untreated plants (P < 0.10), but not in roots. The average S concentrations in shoots of Se-treated wild-type, APS8, and APS9 plants were 7.5 ± 0.6, 9.9 ± 1.2, and 14.4 ± 1.1 mg g−1 dry weight, respectively. When grown in the absence of Se, the shoot S concentrations of wild-type, APS8, and APS9 plants were 6.6 ± 0.5, 9.1 ± 1.1, and 13.1 ± 3.7 mg g−1 dry weight, respectively. The average S levels in roots of Se-treated wild-type, APS8, and APS9 plants were 3.9 ± 0.3, 4.4 ± 0.4, and 4.2 ± 0.3 mg g−1 dry weight, respectively.
Figure 6.

Tissue Se concentrations in shoots of 5-week-old wild-type (WT) and APS8 and APS9 Indian mustard plants supplied for 8 d with different concentrations (5–40 μm) of selenate. Values shown are the average and se of three replicates. The root Se levels are shown in Table II. DW, Dry weight.
Table II.
Root-tissue concentrations of Se in wild-type and APS Indian mustard plants treated with four different concentrations of selenate
| Plant | 5 μm | 10 μm | 20 μm | 40 μm |
|---|---|---|---|---|
| mg g−1 dry wt | ||||
| Wild type | 0.19 ± 0.02 | 0.24 ± 0.01 | 0.59 ± 0.08 | 0.96 ± 0.17 |
| APS8 | 0.48 ± 0.21 | 0.38 ± 0.05 | 0.51 ± 0.28 | 1.59 ± 0.16 |
| APS9 | 0.10 ± 0.01 | 0.28 ± 0.07 | 1.12 ± 0.24 | 1.45 ± 0.12 |
Values shown are the average and se of three replicate plants. The corresponding shoot data are shown in Figure 6.
To investigate the effect of ATP-sulfurylase overexpression on sulfate reduction, we measured the levels of the reduced S compounds glutathione and total thiol in APS and wild-type plants that were not treated with Se. The APS8 and APS9 plants contained about 2-fold higher GSH levels than wild-type plants in their shoots (P < 0.05), and 1.6- and 2.2-fold higher levels in their roots (P < 0.01; Fig. 7A), respectively. In addition, the APS8 plants contained 75% higher thiol levels than wild-type plants in their shoots, and 35% higher thiol levels in roots (P < 0.005; Fig. 7B); the APS9 plants contained 41% and 22% higher thiol levels in shoots and roots, respectively, than wild-type plants (P < 0.05; Fig. 7B).
Figure 7.

Shoot- and root-tissue concentrations of glutathione (A) and thiol (B) in 5-week-old wild-type (white bars) and APS8 (hatched bars) and APS9 (shaded bars) Indian mustard plants. Values shown are the average and se of five replicates. FW, Fresh weight.
APS Plants Show Increased Selenate Reduction
XAS was used to determine which chemical species of Se was accumulated inside the APS and wild-type plants and, thus, the effect of ATP-sulfurylase overexpression on selenate reduction in vivo. The K-edge x-ray absorption near-edge spectra of Se were collected from shoots and roots of APS8, APS9, and wild-type plants treated with 20 μm selenate. The Se spectra from shoots and roots of wild-type plants were almost identical to the selenate reference (Figs. 8A and 9A). In contrast, the K-edge x-ray absorption near-edge spectra from the APS plants appear to show a combination of Se species, with characteristics similar to both the selenate and SeMet references (Figs. 8, B and C, and 9, B and C). These results show that a substantial fraction of the selenate taken up by the APS plants was reduced to organic Se, whereas in wild-type plants all Se was accumulated as selenate. Both APS8 and APS9 plants showed relatively more organic Se and less selenate in roots than in shoots (compare Figs. 8 and 9). Using a conservative estimation, the percentage of Se accumulated as organic Se was at least 70% in APS8 and APS9 root tissues (Fig. 8, B and C) and 50% and 20% in APS8 and APS9 shoot tissues, respectively (Fig. 9, B and C).
Figure 8.

XAS Se spectra from shoots of 5-week-old wild-type and APS8 and APS9 Indian mustard plants supplied with 20 μm selenate for 8 d. A through C show two Se standard reference spectra, i.e. selenate and SeMet, and one sample spectrum. A, Wild-type shoot; B, APS8 shoot; C, APS9 shoot. The spectra shown are from pooled samples from three plants each.
Figure 9.

XAS Se spectra from roots of 5-week-old wild-type (WT) and APS8 and APS9 Indian mustard plants supplied with 20 μm selenate for 8 d. A through D show two Se standard reference spectra, i.e. selenate and SeMet, and one or three sample spectra. A, Wild-type root; B, APS8 root; C, APS9 root; D, wild-type, APS8, and APS9 detopped plants (shoots were removed before Se was added). The spectra shown are from pooled samples from three plants each.
To determine the importance of the shoot for the conversion of selenate to organic Se, K-edge x-ray absorption near-edge spectra of Se were collected from plants that had their shoots removed just before selenate was supplied. These detopped roots were apparently healthy at the time of harvest and had taken up ample selenate for K-edge x-ray absorption near-edge spectra analysis. The K-edge x-ray absorption near-edge spectra of Se in roots of these detopped APS8, APS9, and wild-type plants were almost identical to the selenate reference for all plant lines (Fig. 9D). Apparently, without their shoots, APS plants were not able to reduce selenate.
DISCUSSION
The main finding of this study is that the overexpression of ATP sulfurylase in Indian mustard facilitated the increased reduction of supplied selenate, showing that ATP sulfurylase mediates selenate reduction in vivo and that this enzyme is rate limiting for the assimilation of selenate to organic Se. This was concluded from XAS showing that wild-type Indian mustard plants supplied with selenate accumulated Se in both roots and shoots as selenate, whereas in the transgenic APS plants most of the selenate taken up was reduced and accumulated as an organic form of Se, with a XAS spectrum very similar to that of SeMet (Figs. 8 and 9).
The overexpression of the Arabidopsis APS1 gene in Indian mustard resulted in transgenic plants that exhibited 1.5- to 2-fold higher ATP-sulfurylase activity in their shoots but no increase in activity in roots of mature plants, despite the use of the constitutive 35S promoter; only at the seedling stage was the root ATP sulfurylase activity 2-fold higher in the APS plants (Fig. 2). The lack of increase in ATP sulfurylase in roots of mature plants may have been caused by the degradation of precursor protein, possibly as a result of insufficient plastid-uptake capacity, as was suggested from studies with APS Arabidopsis plants (Y. Chen and T. Leustek, unpublished results).
The observation that the roots of the APS plants were not able to reduce selenate when the shoots were removed (Fig. 9D) suggests that selenate reduction in the APS plants was carried out almost exclusively in the shoots, which is in agreement with the ATP-sulfurylase activity levels (Fig. 2). Alternatively, selenate reduction in roots of APS plants may be dependent on the supply of metabolites or signal molecules by the shoot. Sulfate reduction is thought to occur predominantly in the shoot (Leustek, 1996). The expression patterns of the three Arabidopsis APS genes APS1, APS2, and APS3 in both leaves and roots, however, suggest that sulfate may also be assimilated in roots, perhaps under conditions of high S demand (Leustek, 1996).
When the Arabidopsis APS2 gene was overexpressed in tobacco cells, the transgenic cells showed no difference in growth under standard conditions or in the presence of selenate (Hatzfeld et al., 1998). In the study presented here, overexpression of the APS1 gene in Indian mustard also did not affect growth under standard conditions, but did lead to improved growth in the presence of selenate. Not only did the Indian mustard APS plants reduce selenate at increased rates, they also grew better in the presence of toxic levels of selenate (Figs. 3 and 4), and accumulated up to three times more Se per plant (Figs. 5 and 6). Thus, even though the APS plants accumulated more Se in their tissues, they were more tolerant to high levels of selenate. One possible explanation is that the form of Se accumulated in the APS plants (organic Se, possibly SeMet) was less toxic than the form accumulated in wild-type plants (selenate). However, SeMet was reported to be at least as toxic to animals and plants as selenate (Wilber, 1980; Smith and Watkinson, 1984). An alternative explanation is that the APS plants suffered less from Se-induced S deficiency. Selenate competes with sulfate for uptake by the sulfate transporter, resulting in lower sulfate uptake (Zayed and Terry, 1994). Our experiments were done at suboptimal sulfate concentrations (0.25 mm). Therefore, the 50% higher S levels in the shoots of selenate-treated APS plants compared with wild-type plants (Fig. 5) would be expected to result in increased plant growth.
The better growth of the APS plants in the presence of Se (Figs. 3 and 4) may have led to increased transpiration and thus to increased translocation of selenate and sulfate from root to shoot. This may explain why the Se and S levels were more increased in the shoots of APS plants than in the roots compared with wild type. The increased uptake of selenate and sulfate by the APS plants may have been facilitated by increased levels of sulfate/selenate permease. This sulfate/selenate transporter is known to be repressed by sulfate and selenate (Hawkesford et al., 1993), and this repression may be diminished in the APS plants. Certainly, APS roots had much lower concentrations of selenate (<30% of the root Se consisted of selenate) compared with wild-type roots (close to 100% selenate), as judged from XAS spectra (Fig. 9). We did not measure sulfate ions directly; however, there may have been less sulfate in APS roots because the levels of reduced S compounds (thiol and GSH) were 2-fold higher in APS roots (Fig. 7), whereas total S concentrations were the same in APS and wild-type roots (Table I).
Earlier research has shown that when plants are supplied with selenate, most of it is translocated to the shoot (Asher et al., 1967; Arvy, 1993; Zayed and Terry, 1994; de Souza et al., 1998), but most of the volatile Se is released from the root (Zayed and Terry, 1994). If the assimilation of selenate to organic Se was slow or absent in the roots of the APS plants, as suggested by the XAS results (Fig. 9D), then the large amount of organic Se in APS roots (>70% of the root Se; Fig. 9, B and C) must have come from the shoot, which in turn must have derived it as selenate from the root. These results show that the sequence of events after selenate absorption by APS plants is as follows: (a) the transport of selenate from the root (via the xylem) to the chloroplasts in the shoot, (b) the reduction of selenate to organic Se in chloroplasts, and (c) the translocation of organic Se from the shoot (via the phloem) to the root.
These results were used to develop a model for Se uptake, assimilation, and volatilization in plants, which is shown in Figure 10. Selenate is taken up by a sulfate transporter and quickly transported to the shoot via the xylem. The reduction of selenate by ATP sulfurylase probably occurs predominantly in the shoot, and is a slow and rate-limiting step. After the reduction of selenate, the resulting selenite appears to be converted to an organic form of Se relatively quickly, as shown by XAS. When wild-type plants were supplied with selenite for 7 d and subsequently analyzed by XAS, an organic form of Se was present in both roots and shoots, with an XAS spectrum similar to that of SeMet (de Souza et al., 1998). After the conversion of selenite to organic Se, this organic compound is transported to the root, where it is volatilized to dimethylselenide. Judging from the compounds accumulating in the wild-type and APS plants, overexpression of ATP sulfurylase shifts the rate-limiting step in the Se-volatilization pathway from the reduction of selenate to the volatilization of organic Se. It is not yet clear whether the Se-volatilization rates are any different in the APS plants than in the wild-type plants; this will be the topic of further study.
Figure 10.

A proposed model for the Se flow in Indian mustard plants. The compounds shown in boxes are the Se forms that accumulate in selenate-supplied plants of different lines and under different conditions. Selenate is translocated rapidly from root to shoot, and is accumulated in shoots and roots of wild-type plants because ATP-sulfurylase activity is limiting. When ATP sulfurylase is overexpressed (in APS plants), an organic form of Se (possibly SeMet) is accumulated in shoots and roots. Because detopped roots of APS plants do not accumulate organic Se, selenate assimilation appears to be a predominantly shoot-specific process, and there must be a flow of organic Se from shoot to root. SP, Sulfate permease; ATP-S, ATP sulfurylase; OrgSe, organic selenium.
The higher GSH levels in the roots and shoots of APS plants compared with wild-type plants (Fig. 7A) indicate that ATP sulfurylase is limiting for GSH synthesis. APS Arabidopsis plants showed a similar increase in GSH levels (Y. Chen and T. Leustek, unpublished results). It has been suggested that the availability of Cys is rate limiting for GSH biosynthesis because Cys fed to poplar leaf discs resulted in higher GSH levels (Strohm et al., 1995; Noctor et al., 1996). Because ATP sulfurylase is the key enzyme for Cys biosynthesis, overexpression of this enzyme may lead to higher Cys production and therefore to higher GSH levels. Because GSH plays an important role in plant resistance to oxidative stress and is the precursor of phytochelatins (Rauser, 1995; Zenk, 1996), the APS plants are interesting material to test for their tolerance to metals and other stresses. That ATP sulfurylase may be important for heavy metal tolerance is also suggested by the observation that APS1 gene expression in Arabidopsis was enhanced by Cd treatment (Chen and Leustek, 1995). The higher GSH levels in the APS plants may also have contributed to their increased Se tolerance; it is not known if GSH plays any role in Se tolerance.
In conclusion, these studies have provided better insight into the Se assimilation pathway and the rate-limiting steps involved. Furthermore, we have succeeded in creating transgenic plants that are more tolerant to Se and that accumulate 2- to 3-fold higher Se concentrations in their tissues. These APS plants offer great promise for increasing the efficiency of Se phytoremediation.
ACKNOWLEDGMENTS
We thank Farrel W. Lytle for help with the XAS analyses and Adel Zayed and Mark de Souza for helpful discussions and critical reading of the manuscript.
Abbreviations:
- APS
ATP-sulfurylase-overexpressing transgenic plants
- SeMet
selenomethionine
- XAS
x-ray absorption spectroscopy
Footnotes
This work was supported by Electric Power Research Institute grant no. W04163 to N.T., a TALENT stipend by the Dutch Organization for Scientific Research to E.A.H.P.-S., and Stanford Synchrotron Radiation Laboratory grant no. 2413 to C.M.L. and N.T.
LITERATURE CITED
- Anderson JW. Selenium interactions in sulfur metabolism. In: De Kok LJ, editor. Sulfur Nutrition and Assimilation in Higher Plants: Regulatory, Agricultural and Environmental Aspects. The Hague, The Netherlands: SPB Academic Publishing; 1993. pp. 49–60. [Google Scholar]
- Arvy MP. Selenate and selenite uptake and translocation in bean plants (Phaseolus vulgaris) J Exp Bot. 1993;44:1083–1087. [Google Scholar]
- Asher CJ, Evans CS, Johnson CM. Collection and partial characterization of volatile selenium compounds from Medicago sativa L. Aust J Biol Sci. 1967;20:737–748. [Google Scholar]
- Banuelos GS, Mead R, Wu L, Beuselinck P, Akohoue S. Differential selenium accumulation among forage plant species from soils amended with selenium-enriched plant tissue. J Soil Water Conserv. 1992;47:338–342. [Google Scholar]
- Banuelos GS, Meek DW. Accumulation of selenium in plants grown on selenium-treated soil. J Environ Qual. 1990;19:772–777. [Google Scholar]
- Banuelos GS, Schrale G. Crop selection for removing selenium from the soil. Calif Agric. 1989;43:19–20. [Google Scholar]
- Banuelos GS, Terry N, Zayed A, Wu L (1995) Managing high soil Se with phytoremediation In GE Schuman, GF Vance, eds, Selenium: Mining, Reclamation, and Environmental Impact. Proceedings of the 12th Annual National Meeting of the American Society of Surface Mining and Reclamation, Gillette, WY, pp 394–405
- Banuelos GS, Zayed A, Terry N, Mackey B, Wu L, Akohoue S, Zambrzuski S. Accumulation of selenium by different plant species grown under increasing salt regimes. Plant Soil. 1997;183:49–59. [Google Scholar]
- Burnell JN. Selenium metabolism in Neptunia amplexicaulis. Plant Physiol. 1981;67:316–324. doi: 10.1104/pp.67.2.316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y, Leustek T. Sulfate-regulated expression of ATP sulfurylase and adenosine-5′-phosphosulfate kinase in Brassica juncea (abstract No. 319) Plant Physiol. 1995;108:S-72. [Google Scholar]
- Chen Y, Leustek T, Lee S. Analysis of ATP sulfurylase overexpression in Arabidopsis thaliana (abstract no. 702) Plant Physiol. 1997;114:S-148. [Google Scholar]
- de Souza MP, Pilon-Smits EAH, Lytle CM, Hwang SB, Tai JC, Honma TSU, Yeh L, Terry N. Rate limiting steps in Se assimilation and volatilization by Brassica juncea. Plant Physiol. 1998;117:1487–1494. doi: 10.1104/pp.117.4.1487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dilworth GL, Bandurski RS. Activation of selenate by adenosine 5′-triphosphate sulphurylase from Saccharomyces cerevisiae. Biochem J. 1977;163:521–529. doi: 10.1042/bj1630521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duckart EC, Waldron LJ, Doner HE. Selenium uptake and volatilization from plants growing in soil. Soil Sci. 1992;153:94–99. [Google Scholar]
- Ellman GL. Tissue sulfhydryl groups. Arch Biochem Biophys. 1959;82:70–77. doi: 10.1016/0003-9861(59)90090-6. [DOI] [PubMed] [Google Scholar]
- Fassel VA. Quantitative elemental analyses by plasma emission spectroscopy. Science. 1978;202:183–191. doi: 10.1126/science.202.4364.183. [DOI] [PubMed] [Google Scholar]
- Feinberg AP, Vogelstein B. A technique for radiolabelling DNA restriction endonuclease fragments to high specific activity. Anal Biochem. 1983;132:6–13. doi: 10.1016/0003-2697(83)90418-9. [DOI] [PubMed] [Google Scholar]
- Galli U, Schuepp H, Brunold C. Thiols in cadmium- and copper-treated maize (Zea mays L.) Planta. 1996;198:139–143. [Google Scholar]
- Ganther HE, Levander OA, Saumann CA. Dietary control of selenium volatilization in the rat. J Nutr. 1966;88:55–60. doi: 10.1093/jn/88.1.55. [DOI] [PubMed] [Google Scholar]
- Hansen D, Duda P, Zayed AM, Terry N. Selenium removal by constructed wetlands: role of biological volatilization. Environ Sci Technol. 1998;32:591–597. doi: 10.1021/es0260216. [DOI] [PubMed] [Google Scholar]
- Hatzfeld Y, Cathala N, Grignon C, Davidian J-C. Effect of ATP sulfurylase overexpression in bright yellow 2 tobacco cells. Plant Physiol. 1998;116:1307–1313. doi: 10.1104/pp.116.4.1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawkesford MJ, Davidian J-C, Grignon C. Sulfate/proton cotransport on plasma-membrane vesicles isolated from roots of Brassica napus L.: increased transport in membranes isolated from sulfur-starved plants. Planta. 1993;190:297–304. [Google Scholar]
- Hermsen WLJM, McMahon PJ, Anderson JW. Determination of glutathione in plant extracts as the 1-chloro-2,4-dinitrobenzene conjugate in the presence of glutathione S-transferase. Plant Physiol Biochem. 1997;35:491–496. [Google Scholar]
- Herrin DL, Schmidt GW. Rapid, reversible staining of northern blots prior to hybridization. BioTechniques. 1988;6:196–200. [PubMed] [Google Scholar]
- Hoagland D, Arnon DI (1938) The water culture method for growing plants without soil. California Agricultural Experiment Station Circulation 347, pp 1–39
- Hwang S, Herrin DL. Control of lhc gene transcription by the circadian clock in Chlamydomonas reinhardtii. Plant Mol Biol. 1994;26:557–569. doi: 10.1007/BF00013743. [DOI] [PubMed] [Google Scholar]
- Lauchli A. Selenium in plants: uptake, functions, and environmental toxicity. Bot Acta. 1993;106:455–468. [Google Scholar]
- Leggett JE, Epstein E. Kinetics of sulfate adsorption by barley roots. Plant Physiol. 1956;31:222–226. doi: 10.1104/pp.31.3.222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leustek T. Molecular genetics of sulfate assimilation in plants. Physiol Plant. 1996;97:411–419. [Google Scholar]
- Leustek T, Murillo M, Cervantes M. Cloning of a cDNA encoding ATP sulfurylase from Arabidopsis thaliana by functional expression in Saccharomyces cerevisiae. Plant Physiol. 1994;105:897–902. doi: 10.1104/pp.105.3.897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis BG, Johnson CM, Delwiche CC. Release of volatile selenium compounds by plants: collection procedures and preliminary observations. J Agric Food Chem. 1966;14:638–640. [Google Scholar]
- Logan HM, Cathala N, Grignon C, Davidian JC. Cloning of a cDNA encoded by a member of the Arabidopsis thaliana ATP sulfurylase multigene family: expression studies in yeast and in relation to plant sulfur nutrition. J Biol Chem. 1996;271:12227–12233. doi: 10.1074/jbc.271.21.12227. [DOI] [PubMed] [Google Scholar]
- Martin TD. Determining selenium in wastewater sediment and sludge by flameless atomic absorption. Atomic Absorption Newsletter. 1975;14:109–116. [Google Scholar]
- McConnell KP, Portman OW. Toxicity of dimethyl selenide in the rat and mouse. Proc Soc Exp Biol Med. 1952;79:230–231. doi: 10.3181/00379727-79-19333. [DOI] [PubMed] [Google Scholar]
- McNeal JM, Balisteri LS (1989) Geochemistry and Occurrence of Selenium: An Overview. Selenium in Agriculture and the Environment. Special publication No. 23. Soil Science Society of America, Madison, WI
- Mikkelsen R (1987) Materials and methods for determination of selenium in plants and soils. In Workshop on Analytical Methods for Selenium, Other Trace Elements, and on Quality Control and Quality Assurance, Sacramento, CA, March 24–25, 1987, pp 63–66
- Murphy A, Taiz L. A new vertical mesh transfer technique for metal-tolerance studies in Arabidopsis. Ecotypic variation and copper-sensitive mutants. Plant Physiol. 1995;108:29–38. doi: 10.1104/pp.108.1.29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng BH, Anderson JW. Light-dependent incorporation of selenite and sulphite into selenocysteine and cysteine by isolated pea chloroplasts. Phytochemistry. 1979;18:573–580. [Google Scholar]
- Noctor G, Strohm M, Jouanin L, Kunert KJ, Foyer CH, Rennenberg H. Synthesis of glutathione in leaves of transgenic poplar overexpressing γ-glutamylcysteine synthetase. Plant Physiol. 1996;112:1071–1078. doi: 10.1104/pp.112.3.1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohlendorf HM, Hoffman DJ, Salki MK, Aldrich TW. Embryonic mortality and abnormalities of aquatic birds: apparent impacts of selenium from irrigation drain water. Sci Total Environ. 1986;52:49–63. [Google Scholar]
- Rauser WE. Phytochelatins and related peptides. Plant Physiol. 1995;109:1141–1149. doi: 10.1104/pp.109.4.1141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renosto F, Martin RL, Borrel JL, Nelson DC, Segel IH. ATP sulfurylase from trophosome tissue of Riftia pachyptila (hydrothermal vent tube worm) Arch Biochem Biophys. 1991;290:66–78. doi: 10.1016/0003-9861(91)90592-7. [DOI] [PubMed] [Google Scholar]
- Schwenn JD. Photosynthetic sulphate reduction. Z Naturforsch. 1994;49c:531–539. [Google Scholar]
- Setya A, Murillo M, Leustek T. Sulfate reduction in higher plants: molecular evidence for a novel 5′-adenylylsulfate reductase. Proc Natl Acad Sci USA. 1996;93:13383–13388. doi: 10.1073/pnas.93.23.13383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw WH, Anderson JW. Purification, properties and substrate specificity of adenosine triphosphate sulphurylase from spinach leaf tissue. Biochem J. 1972;127:237–247. doi: 10.1042/bj1270237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith FW, Ealing PM, Hawkesford MJ, Clarkson DT. Plant members of a family of sulfate transporters reveal functional subtypes. Proc Natl Acad Sci USA. 1995;92:9373–9377. doi: 10.1073/pnas.92.20.9373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith GS, Watkinson JH. Selenium toxicity in perennial ryegrass and white clover. New Phytol. 1984;97:557–564. [Google Scholar]
- Strohm M, Jouanin L, Kunert KJ, Pruvost C, Polle A, Foyer CH, Rennenberg H. Regulation of glutathione synthesis in leaves of transgenic poplar (Populus tremula × P. alba) overexpressing glutathione synthetase. Plant J. 1995;7:141–145. [Google Scholar]
- Terry N, Carlson C, Raab TK, Zayed AM. Rates of selenium volatilization among crop species. J Environ Qual. 1992;21:341–344. [Google Scholar]
- Terry N, Zayed AM (1994) Selenium volatilization by plants. In WT Frankenberger Jr, S Benson, eds, Selenium in the Environment. Marcel Dekker, New York, pp 343–369
- Terry N, Zayed AM (1998) Phytoremediation of selenium. In WT Frankenberger Jr, RA Engberg, eds, Environmental Chemistry of Selenium. Marcel Dekker, New York, pp 633–657
- Timmermans MCP, Maliga P, Viera J, Messing J. The pFF plasmids: cassettes utilizing CaMV sequences for expression of foreign genes in plants. J Biotechnol. 1990;14:333–344. doi: 10.1016/0168-1656(90)90117-t. [DOI] [PubMed] [Google Scholar]
- Velinsky D, Cutter GA. Geochemistry of selenium in a coastal salt marsh. Geochim Cosmochim Acta. 1991;55:179–191. [Google Scholar]
- Wilber CG. Toxicology of selenium: a review. Clin Toxicol. 1980;17:171–230. doi: 10.3109/15563658008985076. [DOI] [PubMed] [Google Scholar]
- Wu L, Huang ZH, Burau RG. Selenium accumulation and selenium-salt cotolerance in five grass species. Crop Sci. 1988;28:517–522. [Google Scholar]
- Wu L, Van Mantgem PJ, Guo X. Effects of forage plant and field legume species on soil selenium redistribution, leaching, and bioextraction in soils contaminated by agricultural drainage water. Arch Environ Contam Toxicol. 1996;31:329–338. [Google Scholar]
- Zayed AM, Lytle CM, Terry N. Accumulation and volatilization of different chemical species of selenium by plants. Planta. 1998;206:284–292. [Google Scholar]
- Zayed AM, Terry N. Selenium volatilization in broccoli as influenced by sulfate supply. J Plant Physiol. 1992;140:646–652. [Google Scholar]
- Zayed AM, Terry N. Selenium volatilization in roots and shoots: effects of shoot removal and sulfate level. J Plant Physiol. 1994;143:8–14. [Google Scholar]
- Zenk MH. Heavy metal detoxification in higher plants: a review. Gene. 1996;179:21–30. doi: 10.1016/s0378-1119(96)00422-2. [DOI] [PubMed] [Google Scholar]
- Zieve R, Peterson PJ. Volatilization of selenium from plants and soil. Sci Total Environ. 1984;32:197–202. [Google Scholar]