

Abstract
Many biodiversity hotspots are located in montane regions, especially in the tropics. A possible explanation for this pattern is that the narrow thermal tolerances of tropical species and greater climatic stratification of tropical mountains create more opportunities for climate-associated parapatric or allopatric speciation in the tropics relative to the temperate zone. However, it is unclear whether a general relationship exists among latitude, climatic zonation and the ecology of speciation. Recent taxon-specific studies obtained different results regarding the role of climate in speciation in tropical versus temperate areas. Here, we quantify overlap in the climatic distributions of 93 pairs of sister species of mammals, birds, amphibians and reptiles restricted to either the New World tropics or to the Northern temperate zone. We show that elevational ranges of tropical- and temperate-zone species do not differ from one another, yet the temperature range experienced by species in the temperate zone is greater than for those in the tropics. Moreover, tropical sister species tend to exhibit greater similarity in their climatic distributions than temperate sister species. This pattern suggests that evolutionary conservatism in the thermal niches of tropical taxa, coupled with the greater thermal zonation of tropical mountains, may result in increased opportunities for allopatric isolation, speciation and the accumulation of species in tropical montane regions. Our study exemplifies the power of combining phylogenetic and spatial datasets of global climatic variation to explore evolutionary (rather than purely ecological) explanations for the high biodiversity of tropical montane regions.
Keywords: allopatric speciation, diversification, ecological speciation, latitudinal diversity gradient, niche conservatism
1. Introduction
Montane regions are hotspots of species richness and endemism for many taxa, especially in the tropics [1,2]. Tropical mountains are also often outliers in macroecological studies that correlate species richness with present-day climate, suggesting that montane diversity and endemism might be attributed to historical or evolutionary factors in addition to contemporary ecological processes [3–5]. Yet we know surprisingly little about the processes that lead to increases or decreases in species richness (i.e. speciation, extinction and dispersal) in montane systems [6–8]. Thus, testing mechanistic hypotheses that link the high species richness of tropical montane systems to evolutionary and biogeographic processes is an important challenge for macroecologists and biogeographers.
In a highly influential paper, Janzen [9] noted that tropical mountains generally experience less seasonal variation in temperature relative to temperate mountains. This, he argued, would result in greater thermal stratification along elevational gradients in tropical montane systems than in temperate montane systems. Janzen went on to suggest that such extreme stratification selects for organisms with narrower thermal tolerances in tropical mountains, resulting in reduced dispersal over high- (and low-) elevation passes. Since the publication of his seminal paper, several studies have confirmed Janzen's predictions; in tropical montane regions species exhibit greater turnover along elevational climatic gradients, occupy more restricted elevational ranges and thermal regimes, and have narrower thermal tolerances [10–16].
Although Janzen [9] did not explicitly attempt to explain why the tropics harbour so many more species relative to other regions, there are obvious connections between his conjectures and evolutionary mechanisms promoting geographical variation in species richness [12,17]. If species from temperate-zone mountains generally inhabit broader thermal regimes (and have correspondingly broader thermal tolerances) relative to tropical species, then even the climatic differences between many mountaintops and valleys in the temperate zone may not be dramatic enough to act as dispersal barriers that promote isolation and speciation. In contrast, if tropical species adapt to narrower thermal regimes in response to the limited overlap in temperature at different elevations, then at least two mechanisms might promote greater opportunities for speciation along tropical mountains. If the thermal regimes and thermal tolerances of tropical species are more evolutionarily conserved than those of temperate species, then populations inhabiting adjacent mountaintops or valleys should experience greater isolation and increased opportunities for allopatric speciation [18]. Alternatively, the greater climatic stratification of tropical mountains might increase the likelihood of in situ divergence of populations and parapatric speciation along elevational climatic gradients [17,18]. The former scenario implies niche conservatism at speciation whereas the latter implies niche divergence.
These alternative pathways to tropical speciation predict contrasting patterns of evolution in the elevational distributions and thermal regimes of tropical versus temperate sister species. If thermal niches tend to be more conserved in the tropics than in the temperate zone (thereby promoting greater opportunities for allopatric speciation), then tropical sister species should inhabit similar elevational zones, and thus display a greater overlap in their thermal regimes, than temperate sister species [17]. In contrast, if the stronger elevational zonation of climate in the tropics drives speciation by promoting increased specialization of populations to non-overlapping thermal conditions, then sister species should exhibit a lesser overlap in their elevational distributions and thermal regimes in the tropics relative to the temperate zone [17].
Despite the potentially important linkages between climatic zonation and the build-up of species richness, few studies have examined the relationship between latitude, thermal-niche variation and speciation. Kozak & Wiens [17] compared the elevational and thermal overlaps of temperate and tropical sister species of plethodontid salamanders and found evidence for greater divergence in the thermal regimes of tropical species. In addition, rates of climatic niche divergence in these salamanders are positively correlated with rates of clade diversification and with latitudinal variation in species richness [19]. However, studies on other taxa have found no consistent relationship between latitude and the divergence or conservatism of sister species' thermal regimes [20], nor that rates of thermal regime divergence are actually slower in the tropics [21]. Thus, whether a general relationship between latitude, climatic zonation and speciation exists is unclear and requires further study across a wider range of taxa.
Here, we explore the relationship between spatial variation in climate and speciation in tropical and temperate mountains across 93 pairs of sister species identified from published molecular phylogenetic analyses. The selected species represent several major groups of terrestrial vertebrates and encompass taxa with a wide range of thermoregulation strategies and dispersal abilities. We used climatic information extracted from species' locality data (i) to test the prediction that tropical species occupy narrower elevational ranges and thermal regimes across their ranges than temperate species, and (ii) to examine whether tropical sister species overlap more or less in their elevational ranges and thermal regimes than do temperate sister species.
2. Material and methods
To characterize the elevational and thermal regimes of species, we constructed a database of 33 304 georeferenced localities representing known occurrence sites for 190 species from five vertebrate groups (bats, birds, frogs, lizards and snakes) occurring in the New World (electronic supplementary material, appendix A). We selected these species because they were included in published (mostly molecular) phylogenetic analyses that allowed us to identify pairs of sister species for analyses. Such pairs were identified only when 80 per cent or more of the members of the clade under study (e.g. a genus) had been included in a phylogenetic analysis. To be included in the dataset, each pair of sister species had to be restricted to either the tropics or temperate zone in continental areas of the New World. Further, at least one species in each pair was restricted to montane areas (above 1000 m), but in some pairs, especially of bats, one species also extended to the lowlands. Temperate-zone species were defined as those with distributions restricted to latitudes north of 30° N and tropical-zone species as those with distributions south of 30° N. We did not consider species from temperate South America, given the marked differences in climate between hemispheres [22,23]. The degree of overlap of the geographical distributions of species was not a criterion used to include them in our database, but in general sister species were allopatrically or parapatrically distributed (electronic supplementary material, appendix A). In total, we gathered data for 95 pairs of sister species. However, because sequence data for species in two of the pairs were lacking (identification of sister species in these cases was based on morphology-based phylogenies), only 93 species pairs were included in our analyses of sister species (51 tropical and 42 temperate). All 190 species, however, were included in our analyses of elevational range and thermal regime width (described below).
To associate known occurrence sites with environmental data, we obtained georeferenced localities for all study species from the Global Biodiversity Information Facility portal (http://www.gbif.org/). Using ArcGIS v. 9.2, we then superimposed them on temperature layers at a 1 km2 resolution obtained from the WorldClim database [24], and on a digital elevational model at 0.0625 km2 resolution obtained from the Shuttle Radar Topography Mission (http://www2.jpl.nasa.gov/srtm/). Prior to conducting analyses, we inspected the data for georeferencing errors and excluded obviously misplaced localities. We also discarded localities above and below the 97.5 and 2.5 percentiles of the elevation data for each species as probable outliers. We calculated the elevational range for each species as the difference between the maximum and minimum elevations across all localities.
To characterize the thermal regime of each species, we first extracted the maximum temperature of the warmest month (Bio5) and the minimum temperature of the coldest month (Bio6) for each of its collection sites. We then calculated the width of the thermal regime of each species as the difference between its maximum observed value of Bio5 and its minimum observed value of Bio6. We calculated the degree of overlap in elevational ranges of sister species by dividing the amount of overlap by the elevational range of the species with the smaller range [17]. Therefore, our metric of elevational overlap ranges from 0 (no overlap in elevation) to 1 (complete overlap).
To estimate the degree of overlap in the thermal regimes of sister species, we first estimated the monthly temperature range experienced by each species by calculating the mean minimum and mean maximum temperature across all of its collection sites. We then calculated the overall temperature overlap for each sister species pair using the formula
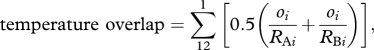 |
where RAi and RBi are the ambient temperature ranges experienced by species A and B, respectively, for month i, and oi is the overlap of RAi and RBi (in °C) for month i [17]. Because the possible range of temperatures that a species can experience differs between regions (i.e. tropics versus temperate zones), this formula standardizes the temperature overlap by the width of each species's thermal regime. Thus, this index is a measure of relative overlap ranging between 0 and 1 for each month, with the total annual temperature overlap ranging between 0 and 12.
Species can restrict the breadth of temperatures they experience by regulating activity times or selecting appropriate microhabitats. Biotic interactions might also restrict a species to a subset of the temperatures that it could otherwise tolerate. Therefore, we do not assume that thermal regime widths calculated from macroclimatic conditions are equivalent to species' actual thermal tolerances. However, our approach does assume that some relationship exists between the large-scale thermal variation across the geographical range of a species and its thermal tolerance. Previous studies have examined the relationship between thermal regime widths estimated from macroclimate data and thermal tolerance/performance breadth, and have generally found that these variables are positively correlated [12,23,25–28].
To test whether tropical species inhabit narrower elevational ranges and narrower thermal regimes than temperate sister species, we first calculated the standardized residuals of a linear regression between the number of localities sampled per species and species' elevational and temperature ranges. This approach was used to control for any effect of sample sizes on the estimated ranges. After testing for assumptions of parametric analyses, we ran one-way analyses of variance (ANOVAs) using these standardized residuals as dependent variables and region (temperate or tropical) as a factor [17]. These analyses were conducted for all taxa pooled, as well as separately for each major taxon.
Changes in the geographical ranges of species occurring after speciation can influence inferences made about the geographical and ecological contexts in which speciation occurred [29]. Because range shifts are more likely to occur over longer periods of time and because the ages of sister species may vary with respect to latitude [30], testing for regional differences in temperature and elevational overlap requires controlling for regional variation in divergence times between species. Therefore, we used analyses of covariance (ANCOVAs) to examine differences in elevational and temperature range overlap between temperate sister species and tropical sister species, while controlling for the effect of divergence times as indexed by genetic distances. If elevational and temperature overlaps are explained by regional variation in opportunities for post-speciational range shifts, rather than by differences in the degree of climatic zonation, then one would expect a significant interaction between timing of sister taxon divergence and region to explain these measures of overlap. To set up these analyses, we obtained mitochondrial DNA sequence data for all species in the analyses from GenBank (electronic supplementary material, appendix A) and calculated uncorrected p-distances (% sequence divergence) between sister species as a surrogate for divergence times. ANCOVAs included elevational and temperature overlaps as dependent variables, region (temperate or tropical) as main factor, p-distance as a covariate to control for divergence times and number of localities as a second covariate to control for differences in sample sizes. We acknowledge that rates of molecular evolution may not be constant across all species. However, using p-distance as a covariate simply assumes that there is a positive relationship between divergence time and genetic distance, not that rates of molecular divergence are constant across all sister species pairs.
We conducted sensitivity analyses to assess the robustness of our results to alternative analytical treatments of the data, including (i) restricting analyses to species with more than 20 locality records, and conducting comparisons of elevational and thermal ranges using raw data (not residuals of the regression between these ranges and number of localities); (ii) calculating elevational overlap using the same formula used to calculate temperature overlap; and (iii) calculating temperature overlap using the formula described by Janzen [9] and not that of Kozak & Wiens [17]. Because none of these procedures led to results different from those obtained using the methods described above, we do not comment on them further, but describe them in the electronic supplementary material, appendices B–G.
3. Results
Considering all five vertebrate groups together and all species (i.e. analyses not restricted to sister species comparisons), we found no strongly significant differences in the elevational ranges of temperate versus tropical species although the analysis was marginally non-significant (ANOVA: n = 190, F = 3.283, p = 0.072; figure 1). When we analysed each group separately, only bats showed the expected pattern of significantly wider elevational ranges for temperate species than for tropical species (n = 30, F = 3.589, p = 0.034); snakes showed a similar but non-significant trend (n = 28, F = 1.463, p = 0.237; figure 1). The other three groups showed trends counter to the prediction, but differences in the elevational ranges of temperate and tropical species were significant only for birds (birds: n = 60, F = 11.391, p = 0.001; frogs: n = 40, F = 0.914, p = 0.345; lizards: n = 32, F = 2.683, p = 0.112; figure 1). By contrast, analyses combining data from all groups and independent analyses of each of the groups consistently supported the prediction that temperate species occur over significantly larger temperature ranges than do tropical species (ANOVA: all groups: n = 190, F = 144.58, p < 0.0001; bats: n = 30, F = 109.96, p < 0.0001; birds: n = 60, F = 55.64, p < 0.0001; frogs: n = 40, F = 35.67, p < 0.0001; lizards: n = 32, F = 16.75, p < 0.0001; snakes: n = 28, F = 7.73, p = 0.01; figure 1).
Figure 1.

Elevation and temperature ranges occupied by temperate and tropical species in five vertebrate groups. Sample sizes are number of species. Boxplots show median, lower and upper quartiles, 5% and 95% percentiles, and outliers. Asterisks indicate statistically significant differences. Ranges are expressed as residuals resulting from the regressions between elevational/temperature range and number of localities sampled.
With respect to the overlap of sister species' elevational ranges, only temperate sister species of bats overlap less in elevational range than tropical sister species (ANCOVA: n = 15, F = 3.473, p = 0.042; figure 2). None of the other analyses showed significant differences across regions in the degree of elevational overlap (ANCOVA: all groups: n = 93, F = 0.575, p = 0.45; birds: n = 30, F = 0.726, p = 0.402; frogs: n = 20, F = 0.713, p = 0.411; lizards: n = 14, F = 0.480, p = 0.504; snakes: n = 14, F = 0.739, p = 0.410; figure 2; full results of ANCOVAs shown in the electronic supplementary material). All groups examined showed trends indicating a greater overlap of the thermal regimes of tropical sister species relative to temperate sister species. However, results were significant only for all the groups combined, birds and lizards (ANCOVA: all groups: n = 93, F = 10.42, p = 0.002; lizards: n = 14, F = 7.862, p = 0.02; bats: n = 15, F = 0.025, p = 0.88; birds: n = 30, F = 6.23, p = 0.02; frogs: n = 20, F = 0.187, p = 0.671; snakes: n = 14, F = 1.420, p = 0.26; figure 2; full results of ANCOVAs shown in the electronic supplementary material).
Figure 2.

Overlap in elevational range and temperature regime of temperate and tropical sister species in five vertebrate groups. Sample sizes are pairs of sister species identified from molecular phylogenetic analyses. Boxplots show median, lower and upper quartiles, 5% and 95% percentiles, and outliers. Asterisks indicate statistically significant differences.
4. Discussion
We explored the relationship between elevational climatic zonation and speciation by comparing latitudinal patterns of variation in the thermal regimes of 93 pairs of vertebrate sister species. Consistent with the predictions of Janzen's climatic zonation hypothesis [9], we found that the geographical distributions of tropical species encompass narrower thermal regimes than those of temperate species from the Northern Hemisphere in the New World. However, in contrast to recent studies on salamanders [17] and frogs [20], we found that sister species tend to exhibit a greater overlap in their thermal regimes in the tropics than in the temperate zone. Together, these results provide new insights on the evolutionary and biogeographic processes associated with the high species richness of tropical montane regions. Specifically, our study suggests that the thermal niches of tropical sister species tend to be narrower and more evolutionarily conserved than those of temperate sister species. As a result, populations may generally experience greater opportunities for isolation and allopatric speciation across elevational thermal gradients in tropical than in temperate montane regions.
Palaeontological and neontological studies have documented patterns of diversification consistent with the idea that rates of speciation are faster at lower latitudes (but see [30], reviewed by [31]). Some investigators have proposed that latitudinal variation in climatic zonation might contribute to this pattern by driving faster speciation in tropical montane organisms [12,17,19,20,32]. However, it has remained unclear whether the greater climatic zonation of tropical mountains might promote speciation by increasing opportunities for allopatric isolation of populations with evolutionarily conserved thermal regimes, or by driving adaptive divergence of populations distributed along elevational climatic gradients [12,17,18,20,32,33]. Based on our expanded sampling of vertebrate taxa, we find that thermal regimes of tropical sister species are generally more conserved than those of temperate ones, suggesting that the greater stability of temperature regimes along tropical mountain slopes could increase opportunities for isolation and allopatric speciation.
Of course, our results only indirectly support the idea that thermal tolerances of species are narrower and more evolutionarily conserved in tropical mountains relative to temperate ones. If species restrict the breadth of temperatures they experience by selecting appropriate microhabitats, regulating activity times or hibernating (especially at higher latitudes), then the use of macroclimatic data could overestimate the actual thermal tolerance breadths of species. However, the few studies that have systematically examined latitudinal variation in thermal tolerance have found a positive relationship between latitude, thermal regime breadth and thermal tolerance range [13,17,23,25–28,34], although interspecific variation in thermal tolerance breadth increases with latitude as a result of the presence of species engaging in periods of extended inactivity [23]. Thus, based on available evidence, our results do not appear to be driven by a greater mismatch between thermal regime breadths and thermal tolerance ranges in the temperate zone. Regardless, explicit tests of whether thermal regime, thermal tolerance, the thermal sensitivity of performance and the ability for thermal acclimation are correlated at geographical scales are sorely needed [35].
Similarly, if biotic interactions (e.g. competition) limit the ranges of species [36,37], then thermal regime widths inferred from geographical distribution data may encompass only a subset of the thermal conditions that species can tolerate. Although biotic interactions are generally thought to be stronger in the tropics [38], there is some evidence to the contrary. For example, Huey [10] found that the turnover of species along tropical versus temperate elevational gradients is unrelated to variation in the number of co-occurring species, a finding that contradicts the idea that competition plays a greater role in driving faunal turnover in the tropics. Likewise, in treefrogs, competition appears to play a role in community assembly in the temperate zone, but not in the tropics [39]. It also seems unlikely that the latitudinal trends we document here are explained entirely by mismatches between the realized and fundamental thermal niches of species. First, given the stronger thermal gradients in tropical mountains, one would expect sister species to have divergent thermal niches, which is the exact opposite of the trend that we recovered. Second, interactions between species should reinforce any impacts of elevational climatic zonation on the physiological tolerances of species. For example, competitive interactions would be more likely to prevent a tropical than a temperate species from encountering a wide range of climatic conditions. Over evolutionary time, one might reasonably expect such pre-emptive occupation of geographical space to cause tropical species to become more physiologically specialized than temperate species. Thus, latitudinal patterns in thermal niche evolution may ultimately arise as a result of interactions between abiotic and biotic factors [19].
In contrast to the predictions of Janzen's climatic zonation hypothesis and the results of a recent study that quantified vertebrate elevational range sizes across latitude [15], we did not find that tropical species had narrower elevational extents than temperate species. However, we point out that patterns of thermal variation, rather than elevation per se, formed the core of Janzen's idea. If species shift their elevational ranges locally to track preferred temperatures (e.g. moving to higher elevations at lower latitudes and vice versa), then range-wide measures of elevational ranges (as we report here) are likely to provide a misleading proxy for the range of thermal conditions over which a species occurs [40]. Previous studies focused on local transects or single mountains [9,10,15] have not encountered this additional complexity, which may emerge when the ranges of elevations occupied by multiple populations of any given species are evaluated. Future studies should address whether populations of elevationally wide-ranging species show a greater propensity for local thermal adaptation in the tropics relative to the temperate zone (e.g. low seasonality in the tropics may enable local adaptation of physiology at small scales more readily).
Our inferences assume that the geographical and ecological contexts of speciation have not been obscured by regional differences (i.e. tropics versus temperate zones) in opportunities for post-speciational changes in the distributions of species. By including genetic distance between species as a covariate in analyses (a surrogate for the time available for post-speciation range shifts), we have attempted to reduce this potential source of error. Genetic distances did not vary significantly between temperate and tropical species in our dataset, suggesting that regional differences in the time available for species ranges to change in position and size [30,41] do not explain the latitudinal trends that we found, although we note that the rate at which species distributions shift following speciation can vary with latitude [42,43]. Further, we cannot rule out the possibility that species in both regions have shifted their elevational and climatic distributions since their formation in response to climate change, mountain uplift or species interactions [36,44,45]. Nevertheless, it is difficult to envision how extensive post-speciation range shifts would result in a statistically significant, rather than random, pattern of the thermal overlap of tropical versus temperate sister species. Regardless, genetic studies of inter-population migration rates will be critical to further test the hypothesis that climatic gradients on mountains are stronger barriers to dispersal in the tropics compared with temperate systems.
Our results suggest that the thermal niches of tropical vertebrates are generally more conserved at the level of sister species than are those of temperate ones, a result consistent with a recent analysis focused on mammals using a different comparative approach [21]. However, exactly why tropical species should exhibit greater conservatism of their thermal niches is unclear and will require further study. One possible explanation is that greater climatic stability in the tropics over time [46] has promoted phylogenetic conservatism in the thermal niches of species. Alternatively, but non-exclusively, biotic interactions in the tropics could restrict the climatic distributions of lineages over time [47].
Although our results suggest that climatic niches of vertebrate sister species are generally more conserved in the tropics than in the temperate zone, this pattern is not universal. For example, the narrower thermal tolerances of tropical plethodontid salamanders lead to a greater tendency for climatic niche divergence and speciation along elevational climatic gradients [17]. In general, the greater climatic zonation of tropical mountains should increase opportunities for either allopatric or parapatric speciations. However, the extent to which such climatic zonation triggers speciation along elevational gradients may ultimately depend on the balance between dispersal and selection [48]. Thus, one might predict that ecological speciation through climatic niche divergence along mountain slopes would be more prevalent in tropical taxa with the most limited dispersal abilities. Among the vertebrate taxa examined to date, plethodontid salamanders exhibit the most extreme spatial genetic structuring of populations [49], suggesting that they are probably more dispersal-limited than birds, mammals, reptiles and anurans. Thus, differences in dispersal abilities might explain the different speciation patterns between plethodontids and other vertebrate taxa.
Taken together, and bearing in mind the potential shortcomings of exploring thermal regime variation from distributional data, our results suggest that tropical sister species exhibit greater evolutionary conservatism in their thermal niches than temperate sister species. Although numerous studies have documented dramatic differences in species richness and rates of diversification between the tropics and temperate zones, few have quantified, to the extent demonstrated here, how the interplay between climatic conditions, the evolution of species' niches and speciation might shape patterns of diversity. This work, we hope, will inspire more detailed examination of the physiological mechanisms that might underlie the patterns we document here, and whether and how they influence variation in speciation mechanisms among taxa and latitudes.
Acknowledgements
This study was conducted as part of the working group Montane Biodiversity in Space and Time: Linking Evolutionary Biology and Macroecology, supported by the National Evolutionary Synthesis Center (NESCent), NSF EF-0423461. We thank the editors, CDC's laboratory group and two anonymous reviewers for helpful comments on the manuscript.
References
- 1.Myers N., Mittermeier R. A., Mittermeier C. G., da Fonseca G. A. B., Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature 403, 853–858 10.1038/35002501 (doi:10.1038/35002501) [DOI] [PubMed] [Google Scholar]
- 2.Orme C. D. L., et al. 2005. Global hotspots of species richness are not congruent with endemism or threat. Nature 436, 1016–1019 10.1038/nature03850 (doi:10.1038/nature03850) [DOI] [PubMed] [Google Scholar]
- 3.Jetz W., Rahbek C. 2002. Geographic range size and determinants of avian species richness. Science 297, 1548–1551 10.1126/science.1072779 (doi:10.1126/science.1072779) [DOI] [PubMed] [Google Scholar]
- 4.Jetz W., Rahbek C., Colwell R. K. 2004. The coincidence of rarity and richness and the potential signature of history in centres of endemism. Ecol. Lett. 7, 1180–1191 10.1111/j.1461-0248.2004.00678.x (doi:10.1111/j.1461-0248.2004.00678.x) [DOI] [Google Scholar]
- 5.Rahbek C., Gotelli N. J., Colwell R. K., Entsminger G. L., Rangel T. F. L. V. B., Graves G. R. 2007. Predicting continental-scale patterns of bird species richness with spatially explicit models. Proc. R. Soc. B 274, 165–174 10.1098/rspb.2006.3700 (doi:10.1098/rspb.2006.3700) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kozak K. H., Wiens J. J. 2010. Niche conservatism drives elevational diversity patterns in Appalachian salamanders. Am. Nat. 176, 40–54 10.1086/653031 (doi:10.1086/653031) [DOI] [PubMed] [Google Scholar]
- 7.Lomolino M. V. 2001. Elevation gradients of species-density: historical and prospective views. Glob. Ecol. Biogeogr. 10, 3–13 10.1046/j.1466-822x.2001.00229.x (doi:10.1046/j.1466-822x.2001.00229.x) [DOI] [Google Scholar]
- 8.McCain C. M. 2009. Global analysis of bird elevational diversity. Glob. Ecol. Biogeogr. 18, 346–360 10.1111/j.1466-8238.2008.00443.x (doi:10.1111/j.1466-8238.2008.00443.x) [DOI] [Google Scholar]
- 9.Janzen D. H. 1967. Why mountain passes are higher in the tropics. Am. Nat. 101, 233–249 10.1086/282487 (doi:10.1086/282487) [DOI] [Google Scholar]
- 10.Huey R. B. 1978. Latitudinal pattern of between-altitude faunal similarity: mountains might be higher in the tropics. Am. Nat. 112, 225–229 10.1086/283262 (doi:10.1086/283262) [DOI] [Google Scholar]
- 11.Wake D. B., Lynch J. F. 1976. The distribution, ecology, and evolutionary history of plethodontid salamanders in tropical America. Sci. Bull. Nat. Hist. Mus. Los Angeles Co. 25, 1–65 [Google Scholar]
- 12.Ghalambor C., Huey R. B., Martin P. R., Tewksbury J. J., Wang G. 2006. Are mountain passes higher in the tropics? Janzen's hypothesis revisited. Int. Comp. Biol. 46, 5–17 10.1093/icb/icj003 (doi:10.1093/icb/icj003) [DOI] [PubMed] [Google Scholar]
- 13.Deutsch C. A., Tewksbury J. J., Huey R. B., Sheldon K. S., Ghalambor C. K., Haak D. C., Martin P. R. 2008. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl Acad. Sci. USA 105, 6668–6672 10.1073/pnas.0709472105 (doi:10.1073/pnas.0709472105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Huey R. B., Deutsch C. A., Tewksbury J. J., Vitt L. J., Hertz P. E., Álvarez Pérez H. J., Garland T. 2009. Why tropical forest lizards are vulnerable to climate warming. Proc. R. Soc. B 276, 1939–1948 10.1098/rspb.2008.1957 (doi:10.1098/rspb.2008.1957) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.McCain C. M. 2009. Vertebrate range sizes indicate that mountains may be ‘higher’ in the tropics. Ecol. Lett. 12, 550–560 10.1111/j.1461-0248.2009.01308.x (doi:10.1111/j.1461-0248.2009.01308.x) [DOI] [PubMed] [Google Scholar]
- 16.Buckley L. B., Jetz W. 2008. Linking global turnover of species and environments. Proc. Natl Acad. Sci. USA 105, 17 836–17 841 10.1073/pnas.0803524105 (doi:10.1073/pnas.0803524105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kozak K. H., Wiens J. J. 2007. Climatic zonation drives latitudinal variation in speciation mechanisms. Proc. R. Soc. B 274, 2995–3003 10.1098/rspb.2007.1106 (doi:10.1098/rspb.2007.1106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Moritz C., Patton J. L., Schneider C. J., Smith T. B. 2000. Diversification of rainforest faunas: an integrated molecular approach. Ann. Rev. Ecol. Syst. 31, 533–563 10.1146/annurev.ecolsys.31.1.533 (doi:10.1146/annurev.ecolsys.31.1.533) [DOI] [Google Scholar]
- 19.Kozak K. H., Wiens J. J. 2010. Accelerated rates of climatic-niche evolution underlie rapid species diversification. Ecol. Lett. 13, 1378–1389 10.1111/j.1461-0248.2010.01530.x (doi:10.1111/j.1461-0248.2010.01530.x) [DOI] [PubMed] [Google Scholar]
- 20.Hua X., Wiens J. J. 2010. Latitudinal variation in speciation mechanisms in frogs. Evolution 64, 429–443 10.1111/j.1558-5646.2009.00836.x (doi:10.1111/j.1558-5646.2009.00836.x) [DOI] [PubMed] [Google Scholar]
- 21.Cooper N., Freckleton R. P., Jetz W. 2011. Phylogenetic conservatism of environmental niches in mammals. Proc. R. Soc. B 278, 2384–2391 10.1098/rspb.2010.2207 (doi:10.1098/rspb.2010.2207) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chown S. L., Sinclair B. J., Leinaas H. P., Gaston K. J. 2004. Hemispheric asymmetries in biodiversity—a serious matter for ecology. PLoS Biol. 2, e406. 10.1371/journal.pbio.0020406 (doi:10.1371/journal.pbio.0020406) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Addo-Bediako A., Chown S. L., Gaston K. J. 2000. Thermal tolerance, climatic variability and latitude. Proc. R. Soc. Lond. B 267, 739–745 10.1098/rspb.2000.1065 (doi:10.1098/rspb.2000.1065) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hijmans R. J., Cameron S. E., Parra J. L., Jones P. G., Jarvis A. 2005. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25, 1965–1978 10.1002/joc.1276 (doi:10.1002/joc.1276) [DOI] [Google Scholar]
- 25.Snyder G. K., Weathers W. W. 1975. Temperature adaptations in amphibians. Am. Nat. 109, 93–101 10.1086/282976 (doi:10.1086/282976) [DOI] [Google Scholar]
- 26.Feder M. E. 1976. Environmental variability and thermal acclimation of metabolism in neotropical and temperate zone salamanders. Phys. Zool. 51, 7–16 [Google Scholar]
- 27.van Berkum F. H. 1988. Latitudinal patterns of thermal sensitivity of sprint speed in lizards. Am. Nat. 132, 327–343 10.1086/284856 (doi:10.1086/284856) [DOI] [Google Scholar]
- 28.Calosi P., Bilton D. T., Spicer J. I., Votier S. C., Atfield A. 2010. What determines a species' geographical range? Thermal biology and latitudinal range size relationships in European diving beetles (Coleoptera: Dytiscidae). J. Anim. Ecol. 79, 194–204 10.1111/j.1365-2656.2009.01611.x (doi:10.1111/j.1365-2656.2009.01611.x) [DOI] [PubMed] [Google Scholar]
- 29.Losos J. B., Glor R. E. 2003. Phylogenetic comparative methods and the geography of speciation. Trends Ecol. Evol. 18, 220–227 10.1016/S0169-5347(03)00037-5 (doi:10.1016/S0169-5347(03)00037-5) [DOI] [Google Scholar]
- 30.Weir J. T., Schluter D. 2007. The latitudinal gradient in recent speciation and extinction rates of birds and mammals. Science 315, 1574–1576 10.1126/science.1135590 (doi:10.1126/science.1135590) [DOI] [PubMed] [Google Scholar]
- 31.Mittlebach G. G., et al. 2007. Evolution and the latitudinal diversity gradient: speciation, extinction and biogeography. Ecol. Lett. 10, 315–331 10.1111/j.1461-0248.2007.01020.x (doi:10.1111/j.1461-0248.2007.01020.x) [DOI] [PubMed] [Google Scholar]
- 32.Wiens J. J. 2004. Speciation and ecology revisited: phylogenetic niche conservatism and the origin of species. Evolution 58, 193–197 [DOI] [PubMed] [Google Scholar]
- 33.Smith T. B., Wayne R. K., Girman D., Bruford M. W. 2005. Evaluating the divergence-with-gene-flow model in natural populations: the importance of ecotones in rainforest speciation. In Tropical rainforests: past, present, and future (eds Bermingham E., Dick C. W., Moritz C.), pp. 148–165 Chicago, IL: The University of Chicago Press [Google Scholar]
- 34.Sunday J. M., Bates A. E., Dulvy N. K. 2011. Global analysis of thermal tolerance and latitude in ectotherms. Proc. R. Soc. B 278, 1823–1830 10.1098/rspb.2010.1295 (doi:10.1098/rspb.2010.1295) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chown S. L., Gaston K. J. 2008. Macrophysiology for a changing world. Proc. R. Soc. B 275, 1469–1478 10.1098/rspb.2008.0137 (doi:10.1098/rspb.2008.0137) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cadena C. D. 2007. Testing the role of interspecific competition in the evolutionary origin of elevational zonation: an example with Buarremon brush-finches (Aves, Emberizidae) in the neotropical mountains. Evolution 61, 1120–1136 10.1111/j.1558-5646.2007.00095.x (doi:10.1111/j.1558-5646.2007.00095.x) [DOI] [PubMed] [Google Scholar]
- 37.Jankowski J. E., Robinson S. K., Levey D. J. 2010. Squeezed at the top: interspecific aggression may constrain elevational ranges in tropical birds. Ecology 91, 1877–1884 10.1890/09-2063.1 (doi:10.1890/09-2063.1) [DOI] [PubMed] [Google Scholar]
- 38.Schemske D. W., Mittlebach G. G., Cornell H. V., Sobel J. M., Roy K. 2009. Is there a latitudinal gradient in the importance of biotic interactions? Ann. Rev. Ecol. Evol. Syst. 40, 245–269 10.1146/annurev.ecolsys.39.110707.173430 (doi:10.1146/annurev.ecolsys.39.110707.173430) [DOI] [Google Scholar]
- 39.Algar A., Kerr J., Currie D. 2011. Quantifying the importance of regional and local filters for community trait structure in tropical and temperate zones. Ecology 92, 903–914 10.1890/10-0606.1 (doi:10.1890/10-0606.1) [DOI] [PubMed] [Google Scholar]
- 40.Cadena C. D., Loiselle B. A. 2007. Limits to elevational distributions in two species of emberizine finches: disentangling the role of interspecific competition, autoecology, and geographic variation in the environment. Ecography 30, 491–504 [Google Scholar]
- 41.Webb T. J., Gaston K. J. 2000. Geographic range size and evolutionary age in birds. Proc. R. Soc. Lond. B 267, 1843–1850 10.1098/rspb.2000.1219 (doi:10.1098/rspb.2000.1219) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Martin P. R., Montgomerie R., Lougheed S. C. 2010. Rapid sympatry explains greater color pattern divergence in high latitude birds. Evolution 64, 336–347 10.1111/j.1558-5646.2009.00831.x (doi:10.1111/j.1558-5646.2009.00831.x) [DOI] [PubMed] [Google Scholar]
- 43.Weir J. T., Price T. D. 2011. Limits to speciation inferred from times to secondary sympatry and ages of hybridizing species along a latitudinal gradient. Am. Nat. 177, 462–469 10.1086/658910 (doi:10.1086/658910) [DOI] [PubMed] [Google Scholar]
- 44.Diamond J. M. 1973. Distributional ecology of New Guinea birds. Science 179, 759–769 10.1126/science.179.4075.759 (doi:10.1126/science.179.4075.759) [DOI] [PubMed] [Google Scholar]
- 45.Terborgh J. 1971. Distribution on environmental gradients: theory and a preliminary interpretation of distributional patterns in the avifauna of the Cordillera Vilcabamba, Peru. Ecology 52, 23–40 10.2307/1934735 (doi:10.2307/1934735) [DOI] [Google Scholar]
- 46.Jansson R., Dynesius M. 2002. The fate of clades in a world of recurrent climatic change: Milankovitch oscillations and evolution. Ann. Rev. Ecol. Syst. 33, 741–747 10.1146/annurev.ecolsys.33.010802.150520 (doi:10.1146/annurev.ecolsys.33.010802.150520) [DOI] [Google Scholar]
- 47.Wiens J. J., et al. 2010. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 13, 1310–1324 10.1111/j.1461-0248.2010.01515.x (doi:10.1111/j.1461-0248.2010.01515.x) [DOI] [PubMed] [Google Scholar]
- 48.Gavrilets S. 2004. Fitness landscapes and the origin of species. Monographs in Population Biology. Princeton, NJ: Princeton University Press [Google Scholar]
- 49.Wake D. B. 2009. What salamanders have taught us about evolution. Ann. Rev. Ecol. Evol. Syst. 40, 333–352 10.1146/annurev.ecolsys.39.110707.173552 (doi:10.1146/annurev.ecolsys.39.110707.173552) [DOI] [Google Scholar]