

Abstract
Are there biological generalities that underlie hybrid sterility or inviability? Recently, around a dozen “speciation genes” have been identified mainly in Drosophila, and the biological functions of these genes are revealing molecular generalities. Major cases of hybrid sterility and inviability seem to result from chromatin evolution and molecular drive in speciation. Repetitive satellite DNAs within heterochromatin, especially at centromeres, evolve rapidly through molecular drive mechanisms (both meiotic and centromeric). Chromatin-binding proteins, therefore, must also evolve rapidly to maintain binding capability. As a result, chromatin binding proteins may not be able to interact with chromosomes from another species in a hybrid, causing hybrid sterility and inviability.
1. Introduction
Are there biological generalities that underlie hybrid sterility or inviability? In other words, do common mechanisms dictate that mules and leopons, for example, are sterile? The widely accepted Dobzhansky-Muller incompatibility (DMI) model of reproductive isolation [1, 2] does not provide an answer to this question. Instead, the DMI model only predicts that combinations of incompatible genes from different species lead to sterile or inviable hybrids. According to Mayr [3], reproductive isolation is an accidental byproduct of speciation. Recently, around a dozen “speciation genes” have been identified, and the biological functions of these genes are revealing molecular generalities that control hybrid sterility and inviability [4–8] (but see [9]). They are chromatin evolution and molecular drive in speciation.
Dover [10] argues, “In the case of many families of genes and noncoding sequences…, fixation of mutations within a population may proceed as a consequence of molecular mechanisms of turnover within the genome [i.e., molecular drive]. …There are circumstances in which the unusual concerted pattern of fixation permits the establishment of biological novelty and species discontinuities [i.e., reproductive isolation]…” Genes encoding heterochromatin proteins may have evolved rapidly to counteract mutations within repetitive DNA sequences in heterochromatin, which accumulate by molecular drive. The molecular drive theory once dominated the field of speciation, supported by the discovery that selfish transposable elements cause hybrid dysgenesis [11–14]. However, this hypothesis has been discounted, as there is no direct evidence that transposons are involved in reproductive isolation [15, 16] (but see [17, 18]). Even the most contemporary textbook concerning speciation [19] does not cite the Dover's [10].
2. Lhr and Hmr of Drosophila
When Drosophila melanogaster females mate with Drosophila simulans males, only weak, sterile, female hybrids eclose, as male hybrids die during larval stages [20]. Watanabe [21] discovered a D. simulans mutation, Lethal hybrid rescue (Lhr), that prevents hybrid larval lethality and restores female hybrid vigor [22]. It was thought that the wild-type allele of D. simulans Lhr was incompatible with X-linked genes from D. melanogaster. It has since been demonstrated that Lhr encodes a heterochromatin protein, HP3, which contains a boundary element-associated factor 32/Su(var)3-7/Stonewall (BESS) domain [23–25]. The X-linked Hybrid male rescue (Hmr) of D. melanogaster [26] has an effect similar to Lhr when mutated, and it also restores female hybrid fertility in this context [27]. Hmr encodes a DNA-binding protein with two myb/SANT-like in Adf-1 (MADF) domains [28].
LHR and HMR may physically interact through their BESS and MADF domains and may colocalize to specific chromatin regions. LHR also interacts with the heterochromatin proteins HP1 and HP6, as demonstrated by yeast two-hybrid (Y2H) experiments, RNA interference (RNAi) knockdown, and Bayesian network analysis [23, 25, 29–31]. The ratio of the number of nonsynonymous substitutions per nonsynonymous site to the number of synonymous substitutions per synonymous site (Ka/Ks) [32] and McDonald-Kreitman (MK) test [33] indicate that Hmr and a subset of genes encoding heterochromatin proteins (including Lhr and HP6) have evolved under positive selection [23, 28, 31, 34]. The involvement of Lhr and Hmr in reproductive isolation is reminiscent of speciation mediated by molecular drive. A comprehensive analysis of LHR, but not HMR, binding sites in the genome has been performed [35].
3. zhr of Drosophila
Involvement of heterochromatic repetitive sequences in hybrid inviability is evident when crosses between D. simulans females and D. melanogaster males (reciprocal to the cross discussed above) are analyzed. Progeny from this cross are sterile, male hybrids, as most female hybrids die during embryogenesis [20, 36]. We discovered zygotic hybrid rescue (zhr), a D. melanogaster gene that prevents female embryonic lethality in this context [37]. Genetic analyses using chromosome deficiencies and duplications [38–40] indicate that female hybrids are rescued if the number of 359-bp repetitive sequences (1.688 satellite) on the D. melanogaster X chromosome is decreased. In addition, hybrids of both sexes are inviable when repetitive sequences are added. In embryos from D. simulans mothers, chromatin regions rich in the 1.688 satellite are not properly condensed [41], resulting in mitotic defects such as chromosome bridges and irregularly spaced nuclei [41, 42].
The 1.688 satellite was one of the earliest sequences cloned in Drosophila [43, 44] and represents more than 4% of the D. melanogaster genome [45–47]. Related sequences are present in D. simulans, but the homology is low [48–51]. Heterochromatin regions rich in the 1.688 satellite may represent binding sites for the putative HMR/LHR complex. However, because zhr only affects hybrid viability when D. simulans females are crossed to D. melanogaster males (not the reciprocal cross), the larval and embryonic hybrid-inviability phenotypes associated with these crosses were thought to be independent (see [37, 52] for additional evidence). However, the possibility remains that female hybrids from D. melanogaster mothers are viable because proteins necessary to cope with D. melanogaster heterochromatin on the X chromosome are supplied maternally. This explanation is consistent with the model proposed by [53, 54]. Identification of proteins that bind to the 1.688 heterochromatin satellite will be informative [55–58]. maternal hybrid rescue (mhr) of D. simulans [52] and Simulans hybrid females rescue (Shfr) [59] represent loci encoding strong 1.688-binding candidates.
Although the 1.688 satellite does not seem to encode any proteins, it is transcribed in ovaries and silenced by the RNAi machinery. This silencing is mediated by repeat-associated small interfering RNA, also called Piwi-associated RNA [60]. In hybrids, failure to silence the 1.688 satellite may lead to heterochromatin decondensation and lethality [54]. Finally, the hybrid lethal on the X (hlx) locus of D. mauritiana affects viability of D. simulans hybrids and has been mapped to heterochromatin [61]. It will be interesting to determine whether this locus also consists of repetitive sequences, similar to zhr.
4. OdsH of Drosophila
In reciprocal crosses between D. mauritiana and D. simulans, female hybrids are fertile but male hybrids are sterile [62]. Many genes have been identified that affect this male hybrid sterility (for a review see [63]). These loci are scattered throughout the two genomes, but an X-linked gene, Odysseus (Ods), plays a particularly important role. When the D. mauritiana allele of Ods is cointrogressed with a closely linked gene onto the D. simulans genetic background, males become sterile [64, 65]. This hybrid male sterility gene has been isolated as Ods-site homeobox (OdsH) [66]. OdsH is paralogous to uncoordinated-4 (unc-4), which is expressed in postmitotic neurons and epidermal cells [67]. In Drosophila, OdsH is thought to have arisen through gene duplication and neofunctionalization, thereby assuming a novel role in spermatogenesis [66, 68, 69]. Ample evidence suggests that OdsH, especially its DNA-binding homeodomain, has evolved under positive selection [66, 69]. Four genes downregulated in sterile male hybrids are thought to lie downstream of OdsH [70]. And misexpressed genes are disproportionately more common on autosomes than on the X in the males with OdsH introgression [71]. Regulatory regions of these genes may contain binding sites for the OdsH transcription factor.
Alternatively, but not mutually exclusively, Bayes and Malik [72] suggested that the ODSH protein localizes to evolutionarily dynamic loci in heterochromatin and that ODSH abundance and localization during premeiotic phases of spermatogenesis are different between D. simulans and D. mauritiana. ODSH from D. mauritiana associates with the heterochromatic Y chromosome of D. simulans, leading to decondensation and male hybrid sterility [72]. These data reveal that rapid heterochromatin evolution affects the onset of male hybrid sterility [72], in addition to hybrid inviability [37, 41]. However, it remains unclear which DNA sequences ODSH binds with the highest affinity.
5. Nup160 and Nup96 of Drosophila
The discovery of strains that restore the fertility of D. simulans/D. melanogaster female hybrids [73] provided the tools to introgress D. simulans chromosomal segments onto the D. melanogaster genetic background [74]. Both male and female introgression homozygotes successfully made were sterile, and the genes responsible for the male and female sterility have been mapped [75–77]. Among them, Nucleoporin 160 (Nup160) of D. simulans was identified as the gene underlying female sterility on the D. melanogaster genetic background [78]. Both D. simulans Nup160 and Nucleoporin 96 (Nup96), which also encodes a component protein of the nuclear pore complex (NPC), cause inviability in D. melanogaster/D. simulans male hybrids [78–80]. This is independent of the F1 hybrid inviability that can be rescued by Lhr mutation and is only revealed in introgression bearers or hemizygotes made from D. melanogaster deficiencies [81, 82].
Population genetics studies have indicated that positive selection is operating in seven nucleoporin genes, including Nup160 and Nup96 [79, 80, 83] and have revealed significant correlated evolution between them [84]. Several hypotheses have been proposed for why nucleoporins are evolving so rapidly in Drosophila [78–80, 83], but here I will focus on the hypothesis most highly related to the molecular drive theory. The NPC forms channels that allow transport of macromolecules between the nucleus and cytoplasm (for a recent review see [85]). In addition, NPC components also function in kinetochore/spindle formation and transcriptional regulation (i.e., dosage compensation) [86–91]. The evolution of scaffold nucleoporins (the NUP107-160 complex) may have accelerated to recognize repetitive sequences in centromeric heterochromatin. In this way, incompatible NPCs may result in hybrid sterility and inviability through improper kinetochore formation. Alternatively, small RNAs derived from repetitive DNA sequences may not be properly trafficked in cells with incompatible NPCs. This leads to chromatin decondensation and, ultimately, sterility or inviability. Such a model has been proposed in the meiotic drive system of D. melanogaster (see below). In this case, mislocalized and truncated Ran GTPase Activating Protein (RanGAP), which is encoded by Segregation distortion (Sd) [92], disrupts proper nuclear transport of small RNAs derived from Responder (Rsp) and ribonucleoprotein complexes that are required to suppress the Rsp satellites [54, 93].
6. Prdm9 of Mice
Evidence for chromatin mechanisms in speciation is not restricted to Drosophila. In the cross between Mus musculus musculus and M. m. domesticus, female hybrids are fertile, but male hybrids are sterile (for a review see [101]; see also [102, 103]). Backcross analyses have indicated that three or more independently segregating loci are involved in this male hybrid sterility. One gene, Hybrid sterility 1 (Hst1) of M. m. domesticus, is polymorphic: the Hst1s allele causes sterility, but Hst1f does not [104]. This situation is similar to the hybrid rescue mutations in Drosophila. The Hst1 locus was mapped to the PR domain zinc finger protein 9 (Prdm9) gene, where PR stands for PRDIBF1 and RIZ homology. Prdm9 encodes a histone H3 lysine 4 (H3K4) trimethyltransferase [94], which is also known as the Meisetz, meiosis-induced factor containing a PR/SET domain and a zinc-finger motif [105]. Hybrid males sterilized by the Prdm9 introgression exhibit frequent dissociation of the X and Y chromosomes during meiosis [94], similar to the sterile male hybrid from a cross between M. m. musculus and M. spretus [106–108]. A gene involved in M. musculus/M. spretus male hybrid sterility and a gene responsible for X-Y dissociation in M. m. musculus/M. m. molossinus hybrid males (the latter termed Sex-chromosome association (Sxa)) have been mapped to the pseudoautosomal region of the X chromosome [95, 96]. The heterochromatin content of this region is quantitatively different among species or subspecies [109, 110].
The DNA-binding domain of PRDM9 consists of multiple, tandem C2H2 zinc finger domains and is evolving rapidly under positive selection in diverse metazoans, including rodents and primates. Rapid evolution of this binding domain likely results from recurrent selection for binding specificity to satellite DNAs [111–113]. The interaction between PRDM9 and repetitive sequences also affects meiotic recombination [114–116]. Histone H3 modifications are typical epigenetic events that determine chromatin status (for reviews see [117, 118]). Genomic regions characterized by heterochromatin-mediated gene silencing are rich in histone H3K9 methylation and have few histone acetylations. In contrast, histones in transcriptionally active euchromatic regions are highly acetylated and methylated at H3K4. Interestingly, chromatin structures regulated by H3K9 methylation, Su(var)3-9, HP1, or the RNAi pathway are required to maintain the structural integrity of tandemly repeated, heterochromatic sequences, like the 1.688 satellite, in D. melanogaster [119].
7. Three Drives in Speciation
The meiotic drive model of male hybrid sterility assumes an arms race between meiotic drive genes and suppressor genes in which male hybrids exhibit segregation distortion or sterility if they inherit drive genes, but not their corresponding suppressors [120, 121]. At first, this model was not accepted because cryptic segregation distortion was not detected in interspecies crosses of Drosophila [122, 123]. In the cross between D. mauritiana and D. simulans, one gene involved in male hybrid sterility is not separable from the meiotic drive gene, too much yin (tmy), by recombination [97]. In addition, the gene Overdrive (Ovd) causes both male hybrid sterility and meiotic drive in aged males when D. pseudoobscura pseudoobscura is crossed with D. p. bogotana [98, 124]. Interestingly, Ovd encodes a protein that contains a MADF DNA-binding domain [98], similar to HMR of D. melanogaster [28].
In the context of speciation, meiotic drive can be the manifestation of molecular drive. The most common example of this phenomenon is centromere drive. The centromere drive model assumes that both DNA and protein components of centromeric chromatin are evolving rapidly and that incompatibilities between rapidly evolving centromeric components may be responsible for hybrid sterility [125]. In particular, the expansion of centromeric repetitive sequences provides more microtubule attachment sites, thereby creating a stronger centromere that tends to be included in the oocyte nucleus [125]. This represents an alternative force from molecular drive that is distinct from a variety of mutational processes that include replication slippage, unequal exchange, transposition, and excision [10, 126–128]. To suppress potential nondisjunction of chromosomes that carry expanded satellite DNAs, the gene centromere identifier (cid) has evolved rapidly in diverse organisms including Drosophila [129, 130]. cid encodes centromeric histone H3-like, a homologue of human Centromere protein A (CENP-A). Examples of centromeric repeats affecting meiotic drive include the Rsp locus of D. melanogaster, which is the target of Sd [131], and the Cent728 repeat, which is responsible for female meiotic drive in the Monkeyflower hybrid between Mimulus guttatus and Mimulus nasutus [99].
8. Applicability and Related Issues
Above I proposed a theory that hybrid sterility and inviability are generally the manifestation of chromatin evolution and molecular drive in the context of speciation, but I do not claim that this model explains every case. Among hybrid incompatibility genes discussed in recent review papers, only 10 of 18 (Table 1 of [5]), 8 of 14 (Table 1 of [6]), and 7 of 14 (Table S1 of [9]) are consistent with this theory. In addition, as most hybrid incompatibility data are from Drosophila, a different trend may appear if reproductive isolation genes are identified from diverse taxa. A famous exception to this theory involves the JYalpha gene in Drosophila. JYalpha encodes a protein with sodium/potassium-exchanging ATPase activity and is located on chromosome 4 in D. melanogaster but on chromosome 3 in D. simulans. Therefore, males carrying homozygous introgression of D. simulans chromosome 4 on the D. melanogaster genetic background are sterile, as they do not inherit JYalpha from either species [100, 132, 133]. This is an example of male hybrid sterility caused by gene transposition between species, which is consistent with the gene duplication and nonfunctionalization model of speciation [134].
Haldane's rule is generally observed when hybrid sterility and inviability are encountered. This rule states that “when in the F1 offspring of two different animal races one sex is absent, rare, or sterile, that sex is the heterozygous [heterogametic (XY or ZW)] sex” [135]. This rule is empirical and seems to be a composite phenomenon [136–138], although the dominance theory is applicable in most cases [139]. Here I propose an additional explanation for Haldane's rule, based on chromatin evolution and molecular drive in speciation. In hybrid animals, chromatin-binding proteins supplied from one species may not be able to recognize the other species' Y or W chromosome, as these chromosomes are generally heterochromatic and have high levels of repetitive satellite DNAs. This results in meiotic or mitotic chromosome decondensation or nondisjunction and leads to hybrid sterility or inviability in the heterogametic sex.
There are several chromatin state systems that have not been discussed yet, which may be related to the present issue. First, inactivation of the X chromosome in primary spermatocytes is necessary for the normal progression of spermatogenesis in heterogametic (XY) males [140] (but see [141, 142]), a process termed meiotic sex chromosome inactivation (MSCI). In some cases, male hybrid sterility may result from ineffective MSCI, as DNA-binding proteins may not be able to recognize and inactivate X chromosomes from different species (e.g., [63, 108]). Second, genomic imprinting affects a subset of genes, resulting in monoallelic and parent-of-origin-specific expression. This process usually depends on DNA methylation or histone modification (e.g., [143–146]). Species-specific variations in epigenetic marks may disrupt imprinting and lead to hybrid inviability. This can explain classic observations of unilateral incompatibility in rodent and flowering plant species (e.g., [147–150]).
9. Conclusion
As has been discussed in this paper, major cases of hybrid sterility and inviability seem to result from chromatin evolution and molecular drive in speciation (Table 1). Repetitive satellite DNAs within heterochromatin, especially at centromeres, evolve rapidly through molecular drive mechanisms (both meiotic and centromeric). Chromatin-binding proteins, therefore, must also evolve rapidly to maintain binding capability. As a result, chromatin-binding proteins may not be able to interact with chromosomes from another species in a hybrid, causing hybrid sterility and inviability (Figure 1).
Table 1.
Hybrid incompatibility genes mentioned in the current paper. Whether data concerning these genes are consistent or inconsistent with the current hypothesis is indicated.
| Gene | Species | Phenotypea | Comment | Consistent | Reference |
|---|---|---|---|---|---|
| Lhr (HP3) | Drosophila melanogaster/D. simulans | F1-L | Interaction with heterochromatin proteins | Yes | [23, 24] |
| Hmr | D. melanogaster/D. simulans | F1-L, FS | Chromatin-binding | Yes | [28] |
| zhr (1.688) | D. melanogaster/D. simulans | F1-L | Centromeric repetitive DNA | Yes | [37, 41] |
| hlx | D. melanogaster/D. simulans | BC-L | Centromeric repetitive DNA? | Yes | [61] |
| OdsH | D. melanogaster/D. simulans | F1, BC-MS | Heterochromatin-binding | Yes | [66, 72] |
| Nup160 | D. melanogaster/D. simulans | BC-L, FS | Centromeric heterochromatin-binding? | Yes | [78, 80] |
| Nup96 | D. melanogaster/D. simulans | BC-L | Centromeric heterochromatin-binding? | Yes | [79] |
| Prdm9 | Mus m. musculus/M. m. domesticus | F1, BC-MS | Histone methylation | Yes | [94] |
| Sxa | M. m. musculus/M. m. domesticus; M. musculus/M. spretus | F1, BC-XY, MS | Heterochromatic repetitive DNA? | Yes | [95, 96] |
| tmy | D. simulans/D. mauritiana | BC-MS | Not separable from the gene causing meiotic drive | Yes | [97] |
| Ovd | D. p. pseudoobscura/D. p. bogotana | F1, BC-MS | Chromatin-binding; also causing meiotic drive | Yes | [98] |
| Cent728 | Mimulus guttatus/M. nasutus | F1, BC-FMD | Centromeric repetitive DNA | Yes | [99] |
| JYalpha | D. melanogaster/D. simulans | BC-MS | Transposition | No | [100] |
aF1: hybrid; BC: (equivalent to) backcross; L: lethal; FS: female sterile; MS: male sterile; XY: XY dissociation; FMD: female meiotic drive.
Figure 1.
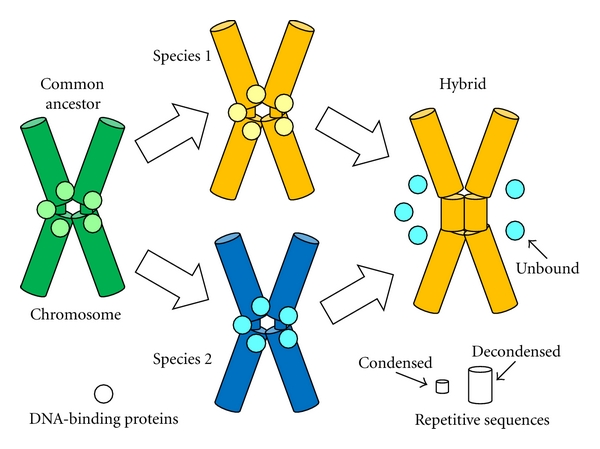
A hybrid sterility and inviability model based on chromatin evolution and molecular drive in speciation. Repetitive satellite DNAs evolve rapidly, thereby accelerating the evolution of chromatin-binding proteins (from the common ancestor to species 1 and species 2). Hybrids are sterile or inviable because the chromatin-binding proteins from species 2 cannot recognize the repetitive sequences of species 1.
Acknowledgment
The author's current study is supported by a Grant-in-Aid for Scientific Research (21570001) from the Japan Society for the Promotion of Science.
References
- 1.Dobzhansky T. Genetics and the Origin of Species. New York, NY, USA: Columbia University Press; 1937. [Google Scholar]
- 2.Muller HJ. Bearing of the Drosophila work on systematics. In: Huxley JS, editor. The New Systematics. Oxford, UK: Claredon Press; 1940. pp. 185–268. [Google Scholar]
- 3.Mayr E. Systematics and the Origin of Species. New York, NY, USA: Columbia University Press; 1942. [Google Scholar]
- 4.Michalak P. Epigenetic, transposon and small RNA determinants of hybrid dysfunctions. Heredity. 2009;102(1):45–50. doi: 10.1038/hdy.2008.48. [DOI] [PubMed] [Google Scholar]
- 5.Johnson NA. Hybrid incompatibility genes: remnants of a genomic battlefield? Trends in Genetics. 2010;26(7):317–325. doi: 10.1016/j.tig.2010.04.005. [DOI] [PubMed] [Google Scholar]
- 6.Presgraves DC. The molecular evolutionary basis of species formation. Nature Reviews Genetics. 2010;11(3):175–180. doi: 10.1038/nrg2718. [DOI] [PubMed] [Google Scholar]
- 7.McDermott SR, Noor MAF. The role of meiotic drive in hybrid male sterility. Philosophical Transactions of the Royal Society B. 2010;365(1544):1265–1272. doi: 10.1098/rstb.2009.0264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Meiklejohn CD, Tao Y. Genetic conflict and sex chromosome evolution. Trends in Ecology and Evolution. 2010;25(4):215–223. doi: 10.1016/j.tree.2009.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nosil P, Schluter D. The genes underlying the process of speciation. Trends in Ecology and Evolution. 2011;26(4):160–167. doi: 10.1016/j.tree.2011.01.001. [DOI] [PubMed] [Google Scholar]
- 10.Dover G. Molecular drive: a cohesive mode of species evolution. Nature. 1982;299(5879):111–117. doi: 10.1038/299111a0. [DOI] [PubMed] [Google Scholar]
- 11.Engels WR, Preston CR. Hybrid dysgenesis in Drosophila melanogaster: the biology of female and male sterility. Genetics. 1979;92(1):161–174. doi: 10.1093/genetics/92.1.161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Doolittle WF, Sapienza C. Selfish genes, the phenotype paradigm and genome evolution. Nature. 1980;284(5757):601–603. doi: 10.1038/284601a0. [DOI] [PubMed] [Google Scholar]
- 13.Kidwell MG. Intraspecific hybrid sterility. In: Ashburner M, Carson HL, Thompson JN Jr., editors. The Genetics and Biology of Drosophila. 3c. London, UK: Academic Press; 1983. pp. 125–154. [Google Scholar]
- 14.Rose MR, Doolittle WF. Molecular biological mechanisms of speciation. Science. 1983;220(4593):157–162. doi: 10.1126/science.220.4593.157. [DOI] [PubMed] [Google Scholar]
- 15.Coyne JA. Mutation rates in hybrids between sibling species of Drosophila . Heredity. 1989;63:p. 2. doi: 10.1038/hdy.1989.87. [DOI] [PubMed] [Google Scholar]
- 16.Hey J. Speciation via hybrid dysgenesis: negative evidence from the Drosophila affinis subgroup. Genetica. 1989;78(2):97–103. [Google Scholar]
- 17.Waugh O’Neill RJ, O’Neill MJ, Marshall Graves JA. Undermethylation associated with retroelement activation and chromosome remodelling in an interspecific mammalian hybrid. Nature. 1998;393(6680):68–72. doi: 10.1038/29985. [DOI] [PubMed] [Google Scholar]
- 18.Labrador M, Farré M, Utzet F, Fontdevila A. Interspecific hybridization increases transposition rates of Osvaldo . Molecular Biology and Evolution. 1999;16(7):931–937. doi: 10.1093/oxfordjournals.molbev.a026182. [DOI] [PubMed] [Google Scholar]
- 19.Coyne JA, Orr HA. Speciation. Sunderland, Mass, USA: Sinauer Associates; 2004. [Google Scholar]
- 20.Sturtevant AH. Genetic studies on Drosophila simulans. I. Introduction. Hybrids with Drosophila melanogaster . Genetics. 1920;5(5):488–500. doi: 10.1093/genetics/5.5.488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Watanabe TK. A gene that rescues the lethal hybrids between Drosophila melanogaster and D. simulans . Japanese Journal of Genetics. 1979;54(5):325–331. [Google Scholar]
- 22.Barbash DA, Roote J, Ashburner M. The Drosophila melanogaster Hybrid male rescue gene causes inviability in male and female species hybrids. Genetics. 2000;154(4):1747–1771. doi: 10.1093/genetics/154.4.1747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Brideau NJ, Flores HA, Wang J, Maheshwari S, Wang X, Barbash DA. Two Dobzhansky-Muller Genes interact to cause hybrid lethality in Drosophila . Science. 2006;314(5803):1292–1295. doi: 10.1126/science.1133953. [DOI] [PubMed] [Google Scholar]
- 24.Prigent SR, Matsubayashi H, Yamamoto MT. Transgenic Drosophila simulans strains prove the identity of the speciation gene Lethal hybrid rescue. Genes and Genetic Systems. 2009;84(5):353–360. doi: 10.1266/ggs.84.353. [DOI] [PubMed] [Google Scholar]
- 25.Greil F, De Wit E, Bussemaker HJ, Van Steensel B. HP1 controls genomic targeting of four novel heterochromatin proteins in Drosophila . EMBO Journal. 2007;26(3):741–751. doi: 10.1038/sj.emboj.7601527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hutter P, Ashburner M. Genetic rescue of inviable hybrids between Drosophila melanogaster and its sibling species. Nature. 1987;327(6120):331–333. doi: 10.1038/327331a0. [DOI] [PubMed] [Google Scholar]
- 27.Barbash DA, Ashburner M. A novel system of fertility rescue in Drosophila hybrids reveals a link between hybrid lethality and female sterility. Genetics. 2003;163(1):217–226. doi: 10.1093/genetics/163.1.217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Barbash DA, Siino DF, Tarone AM, Roote J. A rapidly evolving MYB-related protein causes species isolation in Drosophila . Proceedings of the National Academy of Sciences of the United States of America. 2003;100(9):5302–5307. doi: 10.1073/pnas.0836927100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Giot L, Bader JS, Brouwer C, et al. A Protein interaction map of Drosophila melanogaster . Science. 2003;302(5651):1727–1736. doi: 10.1126/science.1090289. [DOI] [PubMed] [Google Scholar]
- 30.Van Steensel B, Braunschweig U, Filion GJ, Chen M, Van Bemmel JG, Ideker T. Bayesian network analysis of targeting interactions in chromatin. Genome Research. 2010;20(2):190–200. doi: 10.1101/gr.098822.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Brideau NJ, Barbash DA. Functional conservation of the Drosophila hybrid incompatibility gene Lhr . BMC Evolutionary Biology. 2011;11(1, article 57) doi: 10.1186/1471-2148-11-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Nei M. Molecular Evolutionary Genetics. New York, NY, USA: Columbia University Press; 1987. [Google Scholar]
- 33.McDonald JH, Kreitman M. Adaptive protein evolution at the Adh locus in Drosophila . Nature. 1991;351(6328):652–654. doi: 10.1038/351652a0. [DOI] [PubMed] [Google Scholar]
- 34.Barbash DA, Awadalla P, Tarone AM. Functional divergence caused by ancient positive selection of a Drosophila hybrid incompatibility locus. PLoS Biology. 2004;2(6) doi: 10.1371/journal.pbio.0020142. Article ID e142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Filion GJ, van Bemmel JG, Braunschweig U, et al. Systematic protein location mapping reveals five principal chromatin types in Drosophila cells. Cell. 2010;143(2):212–224. doi: 10.1016/j.cell.2010.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hadorn E. Zur autonomie und phasenspezifität der latalität von bastarden zwischen Drosophila melanogaster und Drosophila simulans . Revue Suisse de Zoologie. 1961;68(2):197–207. [Google Scholar]
- 37.Sawamura K, Yamamoto MT, Watanabe TK. Hybrid lethal systems in the Drosophila melanogaster species complex. II. The Zygotic hybrid rescue (Zhr) gene of D. melanogaster . Genetics. 1993;133(2):307–313. doi: 10.1093/genetics/133.2.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sawamura K, Yamamoto MT. Cytogenetical localization of Zygotic hybrid rescue (Zhr), a Drosophila melanogaster gene that rescues interspecific hybrids from embryonic lethality. Molecular and General Genetics. 1993;239(3):441–449. doi: 10.1007/BF00276943. [DOI] [PubMed] [Google Scholar]
- 39.Sawamura K, Fujita A, Yokoyama R, et al. Molecular and genetic dissection of a reproductive isolation gene, zygotic hybrid rescue, of Drosophila melanogaster . Japanese Journal of Genetics. 1995;70(2):223–232. doi: 10.1266/jjg.70.223. [DOI] [PubMed] [Google Scholar]
- 40.Sawamura K, Yamamoto MT. Characterization of a reproductive isolation gene, zygotic hybrid rescue, of Drosophila melanogaster by using minichromosomes. Heredity. 1997;79(1):97–103. [Google Scholar]
- 41.Ferree PM, Barbash DA. Species-specific heterochromatin prevents mitotic chromosome segregation to cause hybrid lethality in Drosophila . PLoS Biology. 2009;7(10) doi: 10.1371/journal.pbio.1000234. Article ID e1000234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sawamura K, Wu CI, Karr TL. Early development and lethality in D. simulans/D. melanogaster hybrids. In: Proceedings of the Annual Drosophila Research Conference, vol. 38; 1997; p. 175. [Google Scholar]
- 43.Carlson M, Brutlag D. Cloning and characterization of a complex satellite DNA from Drosophila melanogaster . Cell. 1977;11(2):371–381. doi: 10.1016/0092-8674(77)90054-x. [DOI] [PubMed] [Google Scholar]
- 44.Hsieh T, Brutlag D. Sequence and sequence variation within the 1.688 g/cm3 satellite DNA of Drosophila melanogaster . Journal of Molecular Biology. 1979;135(2):465–481. doi: 10.1016/0022-2836(79)90447-9. [DOI] [PubMed] [Google Scholar]
- 45.Brutlag DL. Molecular arrangement and evolution of heterochromatic DNA. Annual Review of Genetics. 1980;14:121–144. doi: 10.1146/annurev.ge.14.120180.001005. [DOI] [PubMed] [Google Scholar]
- 46.Hilliker AJ, Appels R. Pleiotropic effects associated with the deletion of heterochromatin surrounding rDNA on the X chromosome of Drosophila . Chromosoma. 1982;86(4):469–490. doi: 10.1007/BF00330122. [DOI] [PubMed] [Google Scholar]
- 47.Lohe AR, Hilliker AJ, Roberts PA. Mapping simple repeated DNA sequences in heterochromatin of Drosophila melanogaster . Genetics. 1993;134(4):1149–1174. doi: 10.1093/genetics/134.4.1149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Barnes SR, Webb DA, Dover G. The distribution of satellite and main-band DNA components in the melanogaster species subgroup of Drosophila. I. Fractionation of DNA in actinomycin D and distamycin A density gradients. Chromosoma. 1978;67(4):341–363. doi: 10.1007/BF00285965. [DOI] [PubMed] [Google Scholar]
- 49.Strachan T, Coen E, Webb D, Dover G. Modes and rates of change of complex DNA families of Drosophila . Journal of Molecular Biology. 1982;158(1):37–54. doi: 10.1016/0022-2836(82)90449-1. [DOI] [PubMed] [Google Scholar]
- 50.Strachan T, Webb D, Dover GA. Transition stages of molecular drive in multiple-copy DNA families in Drosophila . EMBO Journal. 1985;4(7):1701–1708. doi: 10.1002/j.1460-2075.1985.tb03839.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lohe AR, Brutlag DL. Identical satellite DNA sequences in sibling species of Drosophila . Journal of Molecular Biology. 1987;194(2):161–170. doi: 10.1016/0022-2836(87)90365-2. [DOI] [PubMed] [Google Scholar]
- 52.Sawamura K, Taira T, Watanabe TK. Hybrid lethal systems in the Drosophila melanogaster species complex. I. The maternal hybrid rescue (mhr) gene of Drosophila simulans . Genetics. 1993;133(2):299–305. doi: 10.1093/genetics/133.2.299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Hutter P, Roote J, Ashburner M. A genetic basis for the inviability of hybrids between sibling species of Drosophila . Genetics. 1990;124(4):909–920. doi: 10.1093/genetics/124.4.909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ferree PM, Barbash DA. Distorted sex ratios: a window into RNAi-mediated silencing. PLoS Biology. 2007;5(11):2453–2457. doi: 10.1371/journal.pbio.0050303. Article ID e303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hsieh TS, Brutlag DL. A protein that preferentially binds Drosophila satellite DNA. Proceedings of the National Academy of Sciences of the United States of America. 1979;76(2):726–730. doi: 10.1073/pnas.76.2.726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Käs E, Laemmli UK. In vivo topoisomerase II cleavage of the Drosophila histone and satellite III repeats: DNA sequence and structural characteristics. EMBO Journal. 1992;11(2):705–716. doi: 10.1002/j.1460-2075.1992.tb05103.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Marshall WF, Straight A, Marko JF, et al. Interphase chromosomes undergo constrained diffusional motion in living cells. Current Biology. 1997;7(12):930–939. doi: 10.1016/s0960-9822(06)00412-x. [DOI] [PubMed] [Google Scholar]
- 58.Blattes R, Monod C, Susbielle G, et al. Displacement of D1, HP1 and topoisomerase II from satellite heterochromatin by a specific polyamide. EMBO Journal. 2006;25(11):2397–2408. doi: 10.1038/sj.emboj.7601125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Carracedo MC, Asenjo A, Casares P. Location of Shfr a new gene that rescues hybrid female viability in crosses between Drosophila simulans females and D. melanogaster males. Heredity. 2000;84(6):630–638. doi: 10.1046/j.1365-2540.2000.00658.x. [DOI] [PubMed] [Google Scholar]
- 60.Usakin L, Abad J, Vagin VV, De Pablos B, Villasante A, Gvozdev VA. Transcription of the 1.688 satellite DNA family is under the control of RNA interference machinery in Drosophila melanogaster ovaries. Genetics. 2007;176(2):1343–1349. doi: 10.1534/genetics.107.071720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Cattani MV, Presgraves DC. Genetics and lineage-specific evolution of a lethal hybrid incompatibility between Drosophila mauritiana and its sibling species. Genetics. 2009;181(4):1545–1555. doi: 10.1534/genetics.108.098392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.David J, Lemeunier F, Tsacas L, Bocquet C. Hybridization of a new species, Drosophila mauritiana with D. Melanogaster and D. simulans . Annales de Genetique. 1974;17(4):235–241. [PubMed] [Google Scholar]
- 63.Wu CI, Palopoli MF. Genetics of postmating reproductive isolation in animals. Annual Review of Genetics. 1994;28:283–308. doi: 10.1146/annurev.ge.28.120194.001435. [DOI] [PubMed] [Google Scholar]
- 64.Perez DE, Wu CI, Johnson NA, Wu ML. Genetics of reproductive isolation in the Drosophila simulans clade: DNA marker-assisted mapping and characterization of a hybrid-male sterility gene, Odysseus (Ods) . Genetics. 1993;134(1):261–275. doi: 10.1093/genetics/134.1.261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Perez DE, Wu CI. Further characterization of the Odysseus locus of hybrid sterility in Drosophila: one gene is not enough. Genetics. 1995;140(1):201–206. doi: 10.1093/genetics/140.1.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ting CT, Tsaur SC, Wu ML, Wu CI. A rapidly evolving homeobox at the site of a hybrid sterility gene. Science. 1998;282(5393):1501–1504. doi: 10.1126/science.282.5393.1501. [DOI] [PubMed] [Google Scholar]
- 67.Tabuchi K, Yoshikawa S, Yuasa Y, Sawamoto K, Okano H. A novel Drosophila paired-like homeobox gene related to Caenorhabditis elegans unc-4 is expressed in subsets of postmitotic neurons and epidermal cells. Neuroscience Letters. 1998;257(1):49–52. doi: 10.1016/s0304-3940(98)00799-x. [DOI] [PubMed] [Google Scholar]
- 68.Ting CT, Tsaur SC, Sun S, Browne WE, Patel NH, Wu CI. Gene duplication and speciation in Drosophila: evidence from the Odysseus locus . Proceedings of the National Academy of Sciences of the United States of America. 2004;101(33):12232–12235. doi: 10.1073/pnas.0401975101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sun S, Ting CT, Wu CI. The normal function of a speciation gene, Odysseus, and its hybrid sterility effect. Science. 2004;305(5680):81–83. doi: 10.1126/science.1093904. [DOI] [PubMed] [Google Scholar]
- 70.Michalak P, Noor MAF. Association of misexpression with sterility in hybrids of Drosophila simulans and D. mauritiana . Journal of Molecular Evolution. 2004;59(2):277–282. doi: 10.1007/s00239-004-2622-y. [DOI] [PubMed] [Google Scholar]
- 71.Lu X, Shapiro JA, Ting CT, et al. Genome-wide misexpression of X-linked versus autosomal genes associated with hybrid male sterility. Genome Research. 2010;20(8):1097–1102. doi: 10.1101/gr.076620.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bayes JJ, Malik HS. Altered heterochromatin binding by a hybrid sterility protein in Drosophila sibling species. Science. 2009;326(5959):1538–1541. doi: 10.1126/science.1181756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Davis AW, Roote J, Morley T, Sawamura K, Herrmann S, Ashburner M. Rescue of hybrid sterility in crosses between D. melanogaster and D. simulans . Nature. 1996;380(6570):157–159. doi: 10.1038/380157a0. [DOI] [PubMed] [Google Scholar]
- 74.Sawamura K, Davis AW, Wu CI. Genetic analysis of speciation by means of introgression into Drosophila melanogaster . Proceedings of the National Academy of Sciences of the United States of America. 2000;97(6):2652–2655. doi: 10.1073/pnas.050558597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Sawamura K, Yamamoto MT. The minimal interspecific introgression resulting in male sterility in Drosophila . Genetical Research. 2004;84(2):81–86. doi: 10.1017/s0016672304007001. [DOI] [PubMed] [Google Scholar]
- 76.Sawamura K, Karr TL, Yamamoto MT. Genetics of hybrid inviability and sterility in Drosophila: dissection of introgression of D. simulans genes in D. melanogaster genome. Genetica. 2004;120(1–3):253–260. doi: 10.1023/b:gene.0000017646.11191.b0. [DOI] [PubMed] [Google Scholar]
- 77.Sawamura K, Roote J, Wu CI, Yamamoto MT. Genetic complexity underlying hybrid male sterility in Drosophila . Genetics. 2004;166(2):789–796. doi: 10.1534/genetics.166.2.789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Sawamura K, Maehara K, Mashino S, et al. Introgression of Drosophila simulans nuclear pore protein 160 in Drosophila melanogaster alone does not cause inviability but does cause female sterility. Genetics. 2010;186(2):669–676. doi: 10.1534/genetics.110.119867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Presgraves DC, Balagopalan L, Abmayr SM, Orr HA. Adaptive evolution drives divergence of a hybrid inviability gene between two species of Drosophila . Nature. 2003;423(6941):715–719. doi: 10.1038/nature01679. [DOI] [PubMed] [Google Scholar]
- 80.Tang S, Presgraves DC. Evolution of the Drosophila nuclear pore complex results in multiple hybrid incompatibilities. Science. 2009;323(5915):779–782. doi: 10.1126/science.1169123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Sawamura K. Genetics of hybrid inviability and sterility in Drosophila: the Drosophila melanogaster-Drosophila simulans case. Plant Species Biology. 2000;15(3):237–247. [Google Scholar]
- 82.Presgraves DC. A fine-scale genetic analysis of hybrid incompatibilities in Drosophila . Genetics. 2003;163(3):955–972. doi: 10.1093/genetics/163.3.955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Presgraves DC, Stephan W. Pervasive adaptive evolution among interactors of the Drosophila hybrid inviability gene, Nup96 . Molecular Biology and Evolution. 2007;24(1):306–314. doi: 10.1093/molbev/msl157. [DOI] [PubMed] [Google Scholar]
- 84.Clark NL, Aquadro CF. A novel method to detect proteins evolving at correlated rates: identifying new functional relationships between coevolving proteins. Molecular Biology and Evolution. 2010;27(5):1152–1161. doi: 10.1093/molbev/msp324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Köhler A, Hurt E. Gene regulation by nucleoporins and links to cancer. Molecular Cell. 2010;38(1):6–15. doi: 10.1016/j.molcel.2010.01.040. [DOI] [PubMed] [Google Scholar]
- 86.Mendjan S, Taipale M, Kind J, et al. Nuclear pore components are involved in the transcriptional regulation of dosage compensation in Drosophila . Molecular Cell. 2006;21(6):811–823. doi: 10.1016/j.molcel.2006.02.007. [DOI] [PubMed] [Google Scholar]
- 87.Orjalo AV, Arnaoutov A, Shen Z, et al. The Nup107-160 nucleoporin complex is required for correct bipolar spindle assembly. Molecular Biology of the Cell. 2006;17(9):3806–3818. doi: 10.1091/mbc.E05-11-1061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Zuccolo M, Alves A, Galy V, et al. The human Nup107-160 nuclear pore subcomplex contributes to proper kinetochore functions. EMBO Journal. 2007;26(7):1853–1864. doi: 10.1038/sj.emboj.7601642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Capelson M, Liang Y, Schulte R, Mair W, Wagner U, Hetzer MW. Chromatin-bound nuclear pore components regulate gene expression in higher eukaryotes. Cell. 2010;140(3):372–383. doi: 10.1016/j.cell.2009.12.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Mishra RK, Chakraborty P, Arnaoutov A, Fontoura BMA, Dasso M. The Nup107-160 complex and γ-TuRC regulate microtubule polymerization at kinetochores. Nature Cell Biology. 2010;12(2):164–169. doi: 10.1038/ncb2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Vaquerizas JM, Suyama R, Kind J, Miura K, Luscombe NM, Akhtar A. Nuclear pore proteins Nup153 and megator define transcriptionally active regions in the Drosophila genome. PLoS Genetics. 2010;6(2) doi: 10.1371/journal.pgen.1000846. Article ID e1000846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Merrill C, Bayraktaroglu L, Kusano A, Ganetzky B. Truncated RanGAP encoded by the Segregation Distorter Locus of Drosophila . Science. 1999;283(5408):1742–1745. doi: 10.1126/science.283.5408.1742. [DOI] [PubMed] [Google Scholar]
- 93.Tao Y, Masly JP, Araripe L, Ke Y, Hartl DL. A sex-ratio meiotic drive system in Drosophila simulans. I: an autosomal suppressor. PLoS Biology. 2007;5(11):2560–2575. doi: 10.1371/journal.pbio.0050292. Article ID e292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Mihola O, Trachtulec Z, Vlcek C, Schimenti JC, Forejt J. A mouse speciation gene encodes a meiotic histone H3 methyltransferase. Science. 2009;323(5912):373–375. doi: 10.1126/science.1163601. [DOI] [PubMed] [Google Scholar]
- 95.Guenet JL, Nagamine C, Simon-Chazottes D, Montagutelli X, Bonhomme F. Hst-3: an X-linked hybrid sterility gene. Genetical Research. 1990;56(2-3):163–165. doi: 10.1017/s0016672300035254. [DOI] [PubMed] [Google Scholar]
- 96.Imai HT, Wada MY, Moriwaki K. The sex chromosome association (Sxa) gene is located on the X-chromosome in mice. Japanese Journal of Genetics. 1990;65(2):65–69. doi: 10.1266/jjg.65.65. [DOI] [PubMed] [Google Scholar]
- 97.Tao Y, Hartl DL, Laurie CC. Sex-ratio segregation distortion associated with reproductive isolation in Drosophila . Proceedings of the National Academy of Sciences of the United States of America. 2001;98(23):13183–13188. doi: 10.1073/pnas.231478798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Phadnis N, Orr HA. A single gene causes both male sterility and segregation distortion in Drosophila hybrids. Science. 2009;323(5912):376–379. doi: 10.1126/science.1163934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Fishman L, Saunders A. Centromere-associated female meiotic drive entails male fitness costs in monkeyflowers. Science. 2008;322(5907):1559–1562. doi: 10.1126/science.1161406. [DOI] [PubMed] [Google Scholar]
- 100.Masly JP, Jones CD, Noor MAF, Locke J, Orr HA. Gene transposition as a cause of hybrid sterility in Drosophila . Science. 2006;313(5792):1448–1450. doi: 10.1126/science.1128721. [DOI] [PubMed] [Google Scholar]
- 101.Forejt J. Hybrid sterility in the mouse. Trends in Genetics. 1996;12(10):412–417. doi: 10.1016/0168-9525(96)10040-8. [DOI] [PubMed] [Google Scholar]
- 102.Oka A, Mita A, Sakurai-Yamatani N, et al. Hybrid breakdown caused by substitution of the X chromosome between two mouse subspecies. Genetics. 2004;166(2):913–924. doi: 10.1534/genetics.166.2.913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Oka A, Aoto T, Totsuka Y, et al. Disruption of genetic interaction between two autosomal regions and the X chromosome causes reproductive isolation between mouse strains derived from different subspecies. Genetics. 2007;175(1):185–197. doi: 10.1534/genetics.106.062976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Forejt J, Ivanyi P. Genetic studies on male sterility of hybrids between laboratory and wild mice (Mus musculus L.) Genetical Research. 1974;24(2):189–206. doi: 10.1017/s0016672300015214. [DOI] [PubMed] [Google Scholar]
- 105.Hayashi K, Yoshida K, Matsui Y. A histone H3 methyltransferase controls epigenetic events required for meiotic prophase. Nature. 2005;438(7066):374–378. doi: 10.1038/nature04112. [DOI] [PubMed] [Google Scholar]
- 106.Matsuda Y, Hirobe T, Chapman VM. Genetic basis of X-Y chromosome dissociation and male sterility in interspecific hybrids. Proceedings of the National Academy of Sciences of the United States of America. 1991;88(11):4850–4854. doi: 10.1073/pnas.88.11.4850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Matsuda Y, Moens PB, Chapman VM. Deficiency of X and Y chromosomal pairing at meiotic prophase in spermatocytes of sterile interspecific hybrids between laboratory mice (Mus domesticus) and Mus spretus . Chromosoma. 1992;101(8):483–492. doi: 10.1007/BF00352471. [DOI] [PubMed] [Google Scholar]
- 108.Hale DW, Washburn LL, Eicher EM. Meiotic abnormalities in hybrid mice of the C57BL/6J x Mus spretus cross suggest a cytogenetic basis for Haldane’s rule of hybrid sterility. Cytogenetics and Cell Genetics. 1993;63(4):221–234. doi: 10.1159/000133539. [DOI] [PubMed] [Google Scholar]
- 109.Winking H, Nielsen K, Gropp A. Variable positions of NORs in Mus musculus . Cytogenetics and Cell Genetics. 1980;26(2–4):158–164. doi: 10.1159/000131437. [DOI] [PubMed] [Google Scholar]
- 110.Matsuda Y, Chapman VM. In situ analysis of centromeric satellite DNA segregating in Mus species crosses. Mammalian Genome. 1990;1(2):71–77. doi: 10.1007/BF02443781. [DOI] [PubMed] [Google Scholar]
- 111.Oliver PL, Goodstadt L, Bayes JJ, et al. Accelerated evolution of the Prdm9 speciation gene across diverse metazoan taxa. PLoS Genetics. 2009;5(12) doi: 10.1371/journal.pgen.1000753. Article ID e1000753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Thomas JH, Emerson RO, Shendure J. Extraordinary molecular evolution in the PRDM9 fertility gene. PloS One. 2009;4(12, article e8505) doi: 10.1371/journal.pone.0008505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Ponting CP. What are the genomic drivers of the rapid evolution of PRDM9? Trends in Genetics. 2011;27(5):165–171. doi: 10.1016/j.tig.2011.02.001. [DOI] [PubMed] [Google Scholar]
- 114.Baudat F, Buard J, Grey C, et al. PRDM9 is a major determinant of meiotic recombination hotspots in humans and mice. Science. 2010;327(5967):836–840. doi: 10.1126/science.1183439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Myers S, Bowden R, Tumian A, et al. Drive against hotspot motifs in primates implicates the PRDM9 gene in meiotic recombination. Science. 2010;327(5967):876–879. doi: 10.1126/science.1182363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Parvanov ED, Petkov PM, Paigen K. Prdm9 controls activation of mammalian recombination hotspots. Science. 2010;327(5967):p. 835. doi: 10.1126/science.1181495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Jenuwein T, Allis CD. Translating the histone code. Science. 2001;293(5532):1074–1080. doi: 10.1126/science.1063127. [DOI] [PubMed] [Google Scholar]
- 118.Peng JC, Karpen GH. Epigenetic regulation of heterochromatic DNA stability. Current Opinion in Genetics and Development. 2008;18(2):204–211. doi: 10.1016/j.gde.2008.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Peng JC, Karpen GH. H3K9 methylation and RNA interference regulate nucleolar organization and repeated DNA stability. Nature Cell Biology. 2007;9(1):25–35. doi: 10.1038/ncb1514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Frank SH. Divergence of meiotic drive-suppressors as an explanation for sex-biased hybrid sterility and inviability. Evolution. 1991;45(2):262–267. doi: 10.1111/j.1558-5646.1991.tb04401.x. [DOI] [PubMed] [Google Scholar]
- 121.Hurst LD, Pomiankowski A. Causes of sex ratio bias may account for unisexual sterility in hybrids: a new explanation of Haldane’s rule and related phenomena. Genetics. 1991;128(4):841–858. doi: 10.1093/genetics/128.4.841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Johnson NA, Wu CI. An empirical test of the meiotic drive models of hybrid sterility: sex- ratio data from hybrids between Drosophila simulans and Drosophila sechellia . Genetics. 1992;130(3):507–511. doi: 10.1093/genetics/130.3.507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Coyne JA, Orr HA. Further evidence against the involvement of meiotic drive in hybrid sterility. Evolution. 1993;47(2):685–687. doi: 10.1111/j.1558-5646.1993.tb02123.x. [DOI] [PubMed] [Google Scholar]
- 124.Orr HA, Irving S. Segregation distortion in hybrids between the Bogota and USA subspecies of Drosophila pseudoobscura . Genetics. 2005;169(2):671–682. doi: 10.1534/genetics.104.033274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Henikoff S, Ahmad K, Malik HS. The centromere paradox: stable inheritance with rapidly evolving DNA. Science. 2001;293(5532):1098–1102. doi: 10.1126/science.1062939. [DOI] [PubMed] [Google Scholar]
- 126.Smith GP. Evolution of repeated DNA sequences by unequal crossover. Science. 1976;191(4227):528–535. doi: 10.1126/science.1251186. [DOI] [PubMed] [Google Scholar]
- 127.Charlesworth B, Sniegowski P, Stephan W. The evolutionary dynamics of repetitive DNA in eukaryotes. Nature. 1994;371(6494):215–220. doi: 10.1038/371215a0. [DOI] [PubMed] [Google Scholar]
- 128.Csink AK, Henikoff S. Something from nothing: the evolution and utility of satellite repeats. Trends in Genetics. 1998;14(5):200–204. doi: 10.1016/s0168-9525(98)01444-9. [DOI] [PubMed] [Google Scholar]
- 129.Henikoff S, Ahmad K, Platero JS, Van Steensel B. Heterochromatic deposition of centromeric histone H3-like proteins. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(2):716–721. doi: 10.1073/pnas.97.2.716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Malik HS, Henikoff S. Adaptive evolution of Cid, a centromere-specific histone in Drosophila . Genetics. 2001;157(3):1293–1298. doi: 10.1093/genetics/157.3.1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Wu CI, Lyttle TW, Wu ML, Lin GF. Association between a satellite DNA sequence and the responder of segregation distorter in D. melanogaster . Cell. 1988;54(2):179–189. doi: 10.1016/0092-8674(88)90550-8. [DOI] [PubMed] [Google Scholar]
- 132.Muller HJ, Pontecorvo G. Recombinants between Drosophila species the F1 hybrids of which are sterile. Nature. 1940;146(3693):199–200. [Google Scholar]
- 133.Orr HA. Mapping and characterization of a “speciation gene” in Drosophila . Genetical Research. 1992;59(2):73–80. doi: 10.1017/s0016672300030275. [DOI] [PubMed] [Google Scholar]
- 134.Lynch M, Force AG. The origin of interspecific genomic incompatibility via gene duplication. American Naturalist. 2000;156(6):590–605. doi: 10.1086/316992. [DOI] [PubMed] [Google Scholar]
- 135.Haldane JBS. Sex ratio and unisexual sterility in hybrid animals. Journal of Genetics. 1922;12(2):101–109. [Google Scholar]
- 136.Wu CI, Johnson NA, Palopoli MF. Haldane’s rule and its legacy: why are there so many sterile males? Trends in Ecology and Evolution. 1996;11(7):281–284. doi: 10.1016/0169-5347(96)10033-1. [DOI] [PubMed] [Google Scholar]
- 137.Laurie CC. The weaker sex is heterogametic: 75 years of Haldane’s rule. Genetics. 1997;147(3):937–951. doi: 10.1093/genetics/147.3.937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Kulathinal RJ, Singh RS. The molecular basis of speciation: from patterns to processes, rules to mechanisms. Journal of Genetics. 2008;87(4):327–338. doi: 10.1007/s12041-008-0055-x. [DOI] [PubMed] [Google Scholar]
- 139.Turelli M, Orr HA. The dominance theory of Haldane’s rule. Genetics. 1995;140(1):389–402. doi: 10.1093/genetics/140.1.389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Lifschytz E, Lindsley DL. The role of X-chromosome inactivation during spermatogenesis. Proceedings of the National Academy of Sciences of the United States of America. 1972;69(1):182–186. doi: 10.1073/pnas.69.1.182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Lu X, Wu C-I. Sex, sex chromosomes and gene expression. BMC Biology. 2011;9, article 30 doi: 10.1186/1741-7007-9-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Mikhaylova LM, Nurminsky DI. Lack of global meiotic sex chromosome inactivation, and paucity of tissue-specific gene expression on the Drosophila X chromosome. BMC Biology. 2011;9, article 29 doi: 10.1186/1741-7007-9-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Haig D, Graham C. Genomic imprinting and the strange case of the insulin-like growth factor II receptor. Cell. 1991;64(6):1045–1046. doi: 10.1016/0092-8674(91)90256-x. [DOI] [PubMed] [Google Scholar]
- 144.Moore T, Haig D. Genomic imprinting in mammalian development: a parental tug-of-war. Trends in Genetics. 1991;7(2):45–49. doi: 10.1016/0168-9525(91)90230-N. [DOI] [PubMed] [Google Scholar]
- 145.Scott RJ, Spielman M. Deeper into the maize: new insights into genomic imprinting in plants. BioEssays. 2006;28(12):1167–1171. doi: 10.1002/bies.20508. [DOI] [PubMed] [Google Scholar]
- 146.Kinoshita T, Ikeda Y, Ishikawa R. Genomic imprinting: a balance between antagonistic roles of parental chromosomes. Seminars in Cell and Developmental Biology. 2008;19(6):574–579. doi: 10.1016/j.semcdb.2008.07.018. [DOI] [PubMed] [Google Scholar]
- 147.Zechner U, Shi W, Hemberger M, et al. Divergent genetic and epigenetic post-zygotic isolation mechanisms in Mus and Peromyscus . Journal of Evolutionary Biology. 2004;17(2):453–460. doi: 10.1046/j.1420-9101.2003.00656.x. [DOI] [PubMed] [Google Scholar]
- 148.Josefsson C, Dilkes B, Comai L. Parent-dependent loss of gene silencing during interspecies hybridization. Current Biology. 2006;16(13):1322–1328. doi: 10.1016/j.cub.2006.05.045. [DOI] [PubMed] [Google Scholar]
- 149.Kinoshita T. Reproductive barrier and genomic imprinting in the endosperm of flowering plants. Genes and Genetic Systems. 2007;82(3):177–186. doi: 10.1266/ggs.82.177. [DOI] [PubMed] [Google Scholar]
- 150.Wiley CD, Matundan HH, Duselis AR, Isaacs AT, Vrana PB. Patterns of hybrid loss of imprinting reveal tissue- and cluster-specific regulation. PLoS One. 2008;3(10) doi: 10.1371/journal.pone.0003572. Article ID e3572. [DOI] [PMC free article] [PubMed] [Google Scholar]