

Abstract
Two cercosporoid species are respectively described from Mexican whorled milkweed (Asclepias fascicularis), and spineless safflower (Carthamus tinctorius) from California. Passalora californica represents a new pathogen on Asclepias fascicularis, while Ramularia cynarae is confirmed on Carthamus tinctorius and Cynara cardunculus (Asteraceae), and an epitype designated. Pathogenicity is also established for both pathogens based on Koch’s postulate.
Keywords: ITS, Passalora, Ramularia, systematics
INTRODUCTION
During the course of routine collecting of cercosporoid fungi on different host plants in California, two species were collected on respectively Mexican whorled milkweed (Asclepias fascicularis) and spineless safflower (Carthamus tinctorius).
Asclepias fascicularis is an erect, native perennial plant that grows upright and can reach 1 m tall. The plant grows in grasslands, pastures, roadside ditches, stream banks, and on the borders of cultivated fields. Mexican whorled milkweed contains potent neurotoxins and is often responsible for the poisoning of livestock that eat the plant. In 2010 in coastal Santa Clara County, California, milkweed plants were affected by an undescribed leaf spot disease. Initial symptoms consisted of grey-green, irregularly shaped patches on leaves. Patches expanded and turned brown as disease progressed. For severely affected leaves, most of the laminar portion of the leaf could be diseased; such leaves turned brown, twisted and curled, and dried up. Dark green to black fungal growth was consistently observed on both the adaxial and abaxial sides of affected leaves. In advanced stages of the disease, leaf spots expanded up to 2 cm diam.
Spineless selections of Carthamus tinctorius are grown as commercial cut flower crops in coastal California. In 2010, field plantings of spineless safflower in Santa Clara County showed symptoms of a leaf spot disease. Symptoms consisted of oval to mostly round, tan to light brown leaf spots that primarily affected the foliage on the lower third of the single-stem plants. Spots ranged from 3 to 15 mm diam. Spots often had a concentric ring appearance due to alternating tan and light brown rings in the centre of the spot surrounded by a grey outer ring. White sporulation was visible on both the adaxial and abaxial sides of the spots. Hyaline conidiophores emerged as fascicles from leaf stomata, bearing chains of hyaline conidia.
The aims of the present study were to firstly identify the pathogens associated with the leaf spot diseases on the two respective hosts, and secondly to establish pathogenicity, thereby confirming Koch’s postulates.
MATERIALS AND METHODS
Isolates
Single conidial colonies were established from sporulating conidiomata on Petri dishes containing 2 % malt extract agar (MEA; Crous et al. 2009d) as described earlier (Crous et al. 1991). Colonies were sub-cultured onto potato-dextrose agar (PDA), oatmeal agar (OA), synthetic nutrient-poor agar (SNA), and MEA (Crous et al. 2009d), and incubated at 25°C under continuous near-ultraviolet light to promote sporulation. Reference strains are maintained in the CBS-KNAW Fungal Biodiversity Centre (CBS) Utrecht, The Netherlands.
DNA isolation, amplification and phylogenetic analysis
Genomic DNA was isolated from fungal mycelium grown on MEA, using the UltraCleanTM Microbial DNA Isolation Kit (MoBio Laboratories, Inc., Solana Beach, CA, USA) according to the manufacturer’s protocols. The primers V9G (de Hoog & Gerrits van den Ende 1998) and LR5 (Vilgalys & Hester 1990) were used to amplify part of the nuclear rDNA operon spanning the 3′ end of the 18S rRNA gene (SSU), the internal transcribed spacer 1, the 5.8S rRNA gene, the internal transcribed spacer 2 (ITS) and the first 900 bases at the 5′ end of the 28S rRNA gene (LSU). The primers ITS4 (White et al. 1990) and LSU1Fd (Crous et al. 2009b) were used as internal sequence primers to ensure good quality sequences over the entire length of the amplicon. The PCR conditions, sequence alignment and subsequent phylogenetic analysis followed the methods of Crous et al. (2006, 2009a). Sequences were compared with the sequences available in NCBI’s GenBank nucleotide (nr) database using a megablast search and results are discussed in the relevant species notes where applicable. Alignment gaps were treated as new character states. Sequences derived in this study were lodged at GenBank, the alignment in TreeBASE (<treebase.org/treebase/index.html>), and taxonomic novelties in MycoBank (<MycoBank.org>; Crous et al. 2004).
Pathogenicity
Two different approaches were followed to confirm Koch’s postulates on the respective host plants. To demonstrate pathogenicity on Asclepias fascicularis, diseased leaves having ample sporulation were gently rubbed against leaves of healthy potted milkweed plants. Inoculated plants, placed on top of pans containing water, were enclosed in clear plastic bags for 48 h and then maintained in a greenhouse (24 to 26 °C). Leaves of control plants were gently rubbed with sterile cotton and then handled in the same way.
For pathogenicity tests on Carthamus tinctorius, a suspension of mycelial fragments was prepared. The spore-less mycelial growth from agar cultures was removed, placed in water, and then macerated with a polytron homogeniser (Brinkmann, New York). The resulting suspension was filtered through cheesecloth and then sprayed onto potted spineless safflower plants. Inoculated plants, placed on top of pans containing water, were enclosed in clear plastic bags for 48 h and then maintained in a greenhouse (24 to 26 °C). Control plants were sprayed with sterile distilled water and then handled in the same way.
Morphology
Morphological descriptions are based on preparations made from host material in clear lactic acid, with 30 measurements determined per structure, using a Zeiss Axioscope 2 microscope with differential interference contrast (DIC) illumination. Colony characters and pigment production were noted after 2 wk of growth at 25 °C on MEA, PDA, and OA (Crous et al. 2009d). Colony colours (surface and reverse) were rated according to the colour charts of Rayner (1970).
RESULTS AND DISCUSSION
Phylogeny
Approximately 1700 bases, spanning the ITS and LSU regions, were obtained from the sequenced cultures. Only the ITS sequences were used in the phylogenetic analyses. The manually adjusted ITS alignment contained 34 taxa (including the Cladosporium bruhnei outgroup sequence) and, of the 509 characters (including alignment gaps) used in the phylogenetic analysis, 96 were parsimony-informative, 99 were variable and parsimony-uninformative, and 314 were constant. Only the first 1 000 equally most parsimonious trees were retained from the heuristic search, the first of which is shown in Fig. 1 (TL = 353, CI = 0.756, RI = 0.879, RC = 0.665). The phylogenetic tree of the ITS region (Fig. 1) shows that the obtained sequences cluster in the Mycosphaerellaceae, specifically in the clades called “Clade 3, Ramularia” (Crous et al. 2009c) and close relatives to “Clade 7, Dothistroma” (Crous et al. 2009c).
Fig. 1.
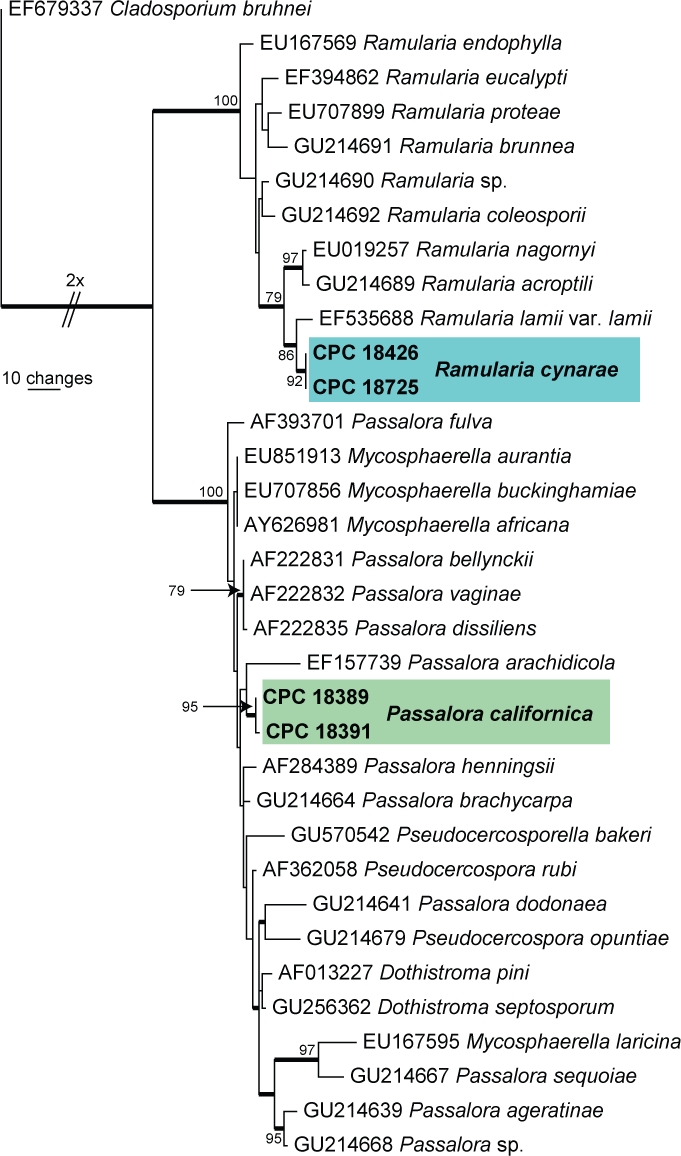
The first of 1 000 equally most parsimonious trees obtained from a heuristic search with 100 random taxon additions of the LSU sequence alignment. The scale bar shows 10 changes and bootstrap support values > 75 % from 1 000 replicates are shown at the nodes. Novel sequences generated in this study are shown in bold and the two treated species in coloured boxes. The tree was rooted to Cladosporium bruhnei (GenBank EF679337).
Pathogenicity
Fourteen days after inoculation of Asclepias fascicularis, symptoms similar to those seen in the field appeared on inoculated milkweed leaves and sporulation was observed 6 to 8 d later. The fungal sporulation was examined and found to be morphologically the same as the originally described fungus. Control plants did not develop the leaf spot disease. The experiment was repeated and the results were the same. Ten days after inoculation of Carthamus tinctorius, symptoms similar to those seen in the field appeared on inoculated plants and sporulation was observed 5 to 7 d later. The fungal sporulation was examined and found to be morphologically the same as the originally described fungus. Water-treated control plants did not develop the leaf spot disease. The experiment was repeated and the results were the same.
Taxonomy
Passalora californica S.T. Koike & Crous, sp. nov.
MycoBank MB517865
(Fig. 2)
Fig. 2.

Passalora californica (CBS H-20512). A, B. Leaves of Asclepias fascicularis infected with P. californica. C. Fructifications on leaf surface. D. Diffuse red pigment surrounding colonies on OA. E. Red crystals forming in PDA. F, G. Conidiophores with conidiogenous cells that can become cylindrical and elongated (arrows denote scars). I. Conidia with characteristic basal attenuation. Bars = 10 μm.
Etymology: Named after the state in which it was collected, California, USA.
Passalorae clavatae var. hansenii similis, sed conidiis angustioribus, (32–)55–95(–180) × (4–)5–6 μm, diametro maximo in medio cellulae basalis.
Typus: USA: California: Santa Clara County, on leaves of Asclepias fascicularis, 19 July 2010, S.T. Koike (CBS H-20512 – holotypus; cultures ex-holotype CPC 18389 = CBS 128857, CPC 18391 (GenBank accession numbers: ITS HQ728115 and HQ728116, for CPC 18389 and 18391, respectively).
Leaf spots irregular, frequently covering the breadth and length of the leaf, black due to profuse sporulation. Conidiomata amphigenous, sporodochial, arising from stromata; stroma globose, brown, 30–100 μm wide, 10–30 μm high, giving rise to conidiophores. Conidiophores in dense sporodochia, brown, verruculose, frequently reduced to conidiogenous cells or with one supporting cell, 15–25 × 3–8 μm, subcylindrical, mostly straight, at times once geniculate-sinuous; under moist conditions on underside of leaf developing further, sporodochia consisting of a mixture of short, stubby, and longer flexuous conidiophores, straight, subcylindrical, sometimes geniculate-sinuous at apex, up to 100 μm long, and 4–5 μm wide. Conidiogenous cells on reduced conidiophores terminal, integrated, brown, verruculose, 10–15 × 4–6 μm; scars apical and lateral, thickened, darkened, refractive, 1–1.5 μm diam; conidiogenous cells on elongated conidiophores terminal, subcylindrical, straight or geniculate-sinuous, medium brown, 15–35 × 4–5 μm. Conidia solitary, obclavate to subcylindrical, brown, verruculose, guttulate, apex obtusely rounded, base obconically truncate with prominent taper in basal cell; hilum 2 μm diam, darkened, thickened, refractive, (32–)55–95(–180) × (4–)5–6 μm, (1–)3–5(–9)-septate; conidia formed on PDA and MEA became up to 300 μm long, but retained the same conidial width and characteristic obconically truncate basal cell with prominent taper.
Culture characteristics: Colonies erumpent, irregular, with sparse aerial mycelium and feathery margins, reaching up to 4 mm on all media tested after 2 wk at 25 °C. On MEA surface umber, reverse chestnut; on PDA surface grey-olivaceous, reverse fuscous-black; on SNA surface rust, with red crystals forming in agar, reverse umber; on OA surface grey-olivaceous, with red pigment diffusing into agar.
Notes: Several species of Passalora have been described from Asclepias that need to be compared to P. californica. Passalora clavata var. clavata has olivaceous to olivaceous-brown, 1–6-septate conidia, 20–80 × (3–)4–6(–7) μm, thus smaller than those of P. californica. Passalora clavata var. hansenii (on Asclepias syrica, California), is similar to P. californica in having 1–14-septate conidia, 15–100(–180) × (4–)5–8(–10) μm, but differs in having conidia that are wider (4–)5–8(–10) μm, being widest in the second or third basal cell, not in the middle of the basal cell as in P. californica, which has a prominently abrupt basal taper (basal cell abruptly obconic, versus long obconically truncate basal region in P. clavata var. clavata and P. clavata var. hansenii; Braun & Mel’nik 1997). Other species on this host include P. elaeochroma, which has strongly geniculate-sinuous conidiogenous cells and loosely fasciculate, frequently branched conidiophores (conidia 20–90 × 4–6.5 μm, 1–7-septate), and P. venturioides, which has secondary hyphae with solitary conidiophores, diffuse leaf spots, and shorter conidia, 20–80(–100) × 4–7 μm, 1–10-septate (Braun & Mel’nik 1997). The two cultures of P. californica sequenced in this study differed with one nucleotide in their second internal transcribed spacer region and were distantly related to, amongst others, Mycosphaerella arachidis (GenBank EF157739) and Phaeoramularia dissiliens (GenBank AF222835) (Fig. 1).
Ramularia cynarae Sacc., Michelia 1: 536 (1879).
Synonyms: See Braun (1998).
(Fig. 3)
Fig. 3.

Ramularia cynarae (CBS H-20513). A. Leaf spots on artichoke (Cynara cardunculus). B. Conidiophore fascicles on leaf surface. C–F. Branched conidial chains in vivo. G–K. Conidiophores and branched conidial chains on SNA. Bars = 10 μm.
Leaf spots amphigenous, medium brown, with pale brown centre, subcircular to somewhat ellipsoid, up to 14 mm diam. Mycelium internal and external, consisting of hyaline, smooth, branched, septate hyphae, 1.5–2 μm wide. Conidiophores fasciculate, amphigenous, subcylindrical in loose fascicles, arising from brown stomata, erect, geniculate-sinuous, 0–4-septate, hyaline, smooth, 60–120 × 2–3 μm. Conidiogenous cells terminal, unbranched, smooth, hyaline, subcylindrical, 20–50 × 2–3 μm; scars terminal and lateral, thickened, darkened and somewhat refractive, 1 μm diam. Conidia hyaline, smooth, with obconically truncate ends. Ramoconidia subcylindrical to obclavate, (15–)20–25(–45) × (3–)3.5–4 μm, 1(–3)-septate, with 1–3 subdenticulate hila that are thickened, darkened and somewhat refractive, 1–1.5 μm diam. Intercalary conidia aseptate, ellipsoid-ovoid, (7–)9–11(–15) × (3.5–)4 μm, 0(–1)-septate; terminal conidia aseptate, ellipsoid-ovoid, apex rounded to obconically truncate, base obconically truncate, (5–)7–9(–10) × 3–4 μm; hila thickened, darkened, somewhat refractive, 1 μm diam. When sporulating on SNA, ramoconidia were (12–)17–22(–32) × (2.5–)3–4 μm, intercalary conidia (7–)8–10(–13) × 4–5(–6) μm, and terminal conidia 7–9 × 4–5 μm.
Culture characteristics: Colonies erumpent, spreading, surface folded, with sparse to moderate aerial mycelium, and smooth, lobate margins, reaching 15–20 mm after 2 wk at 25 °C. On MEA surface smoke-grey, reverse iron-grey; on PDA surface pale olivaceous-grey in middle, olivaceous-grey in outer region, iron-grey underneath; on OA white in middle (due to profuse sporulation), olivaceous-grey in outer region.
Specimens examined: France: Saintes: Brunaud, on Cynara scolymus, herb. Saccardo in PAD, holotype. USA: California: Santa Clara County, Morgan Hill, on leaves of Carthamus tinctorius, 19 Oct. 2010, S.T. Koike (CBS H-20513, cultures CPC 18725, 18726 = CBS 128779 (GenBank accession number: HQ728118); Monterey County, Castroville, on leaves of Cynara cardunculus, 10 Aug. 2010, S.T. Koike (CBS H-20514 – epitypus hic designatus; cultures ex-epitype CPC 18427, 18426 = CBS 128912; GenBank accession number: HQ728117).
Notes: Based on a recent revision of the genus Ramularia (Braun 1998), two species were accepted on Carthamus, namely R. cynarae and R. cercosporelloides (a nom. nov. for Cercosporella carthami, see Braun 1998). Ramularia cercosporelloides is distinct in having obovoid (–subclavate), cylindrical conidia, that are longer and wider, (15–)20–45(–55) × 3.5–10 μm, than the range observed in the present specimens. The description provided by Braun (1998) for R. cynarae as a wide range pathogen on Asteraceae, however, strongly overlaps with the present collections, conidia being (5–)10–35 × (1.5–)2–5(–7) μm) (Braun 1998). Isolates obtained from Cynara in the present study were found to be identical in morphology and on their ITS DNA sequences to isolates from Carthamus, supporting the hypothesis that R. cynarae is a wide-range pathogen on Asteraceae, rather than a species complex (Braun 1998). A species of Ramularia identified as R. carthami has previously been reported in California on agronomic safflower grown for oil production in northern, inland counties in 2005 (Hostert et al. 2006). Given our present knowledge of the Ramularia complex occurring on Asteraceae, this record will have to be re-confirmed, as it may represent R. cynarae.
Acknowledgments
We thank the technical staff, Arien van Iperen (cultures), Marjan Vermaas (photo plates), and Mieke Starink-Willemse (DNA isolation, amplification and sequencing) for their invaluable assistance.
REFERENCES
- Braun U. (1998) A monograph of Cercosporella, Ramularia and allied genera (phytopathogenic Hyphomycetes). Vol. 2 Eching: IHW-Verlag; [Google Scholar]
- Braun U, Mel’nik VA. (1997) Cercosporoid fungi fom Russia and adjacent countries. Proceedings of the Komarov Botanical Institute 20: 1–130 [Google Scholar]
- Crous PW, Gams W, Stalpers JA, Robert V, Stegehuis G. (2004) MycoBank: an online initiative to launch mycology into the 21st century. Studies in Mycology 50: 19–22 [Google Scholar]
- Crous PW, Groenewald JZ, Risède J-M, Simoneau P, Hyde KD. (2006) Calonectria species and their Cylindrocladium anamorphs: species with clavate vesicles. Studies in Mycology 55: 213–226 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Groenewald JZ, Summerell BA, Wingfield BD, Wingfield MJ. (2009a) Co-occurring species of Teratosphaeria on Eucalyptus. Persoonia 22: 38–48 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Schoch CL, Hyde KD, Wood AR, Gueidan C, Hoog GS de, Groenewald JZ. (2009b) Phylogenetic lineages in the Capnodiales. Studies in Mycology 64: 17–47 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Summerell BA, Carnegie AJ, Wingfield MJ, Hunter GC, Burgess TI, Andjic V, Barber PA, Groenewald JZ. (2009c) Unraveling Mycosphaerella: do you believe in genera? Persoonia 23: 99–118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Verkley GJM, Groenewald JZ, Samson RA. (eds) (2009d) Fungal Biodiversity. CBS Laboratory Manual Series 1: 1–269 Centraalbureau voor Schimmelcultures, Utrecht, Netherlands: [Google Scholar]
- Crous PW, Wingfield MJ, Park RF. (1991) Mycosphaerella nubilosa a synonym of M. molleriana. Mycological Research 95: 628–632 [Google Scholar]
- Hoog GS de, Gerrits van den Ende AHG. (1998) Molecular diagnostics of clinical strains of filamentous basidiomycetes. Mycoses 41: 183–189 [DOI] [PubMed] [Google Scholar]
- Hostert ND, Thomas SL, Fogel DL, Blomquist CL, Davis RM. (2006) First report of Ramularia carthami, causal agent of Ramularia leaf spot of Safflower in California. Plant Disease 90: 1260 [DOI] [PubMed] [Google Scholar]
- Rayner RW. (1970) A mycological colour chart. CMI and British Mycological Society; Kew, Surrey, England: [Google Scholar]
- Vilgalys R, Hester M. (1990) Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. Journal of Bacteriology 172: 4238–4246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- White TJ, Bruns T, Lee J, Taylor J. (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ. (eds), PCR protocols: a guide to methods and applications: 315–322 Academic Press, San Diego, California, USA: [Google Scholar]