

Abstract
The mechanisms that facilitate success of an invasive species include both ecological and evolutionary processes. Investigating the evolutionary dynamics of founder populations can enhance our understanding of patterns of invasiveness and provide insight into management strategies for controlling further establishment of introduced populations. Our aim is to analyze the evolutionary consequences of ecological processes (i.e., propagule pressure and threshold density effects) that impact successful colonization. We address our questions using a spatially-explicit modeling approach that incorporates dispersal, density dependent population growth, and selection. Our results show that adaptive evolution may occur in small or sparse populations, providing a means of mitigating or avoiding inverse density dependent effects (i.e., Allee effects). The rate at which this adaptation occurs is proportional to the amount of genetic variance and is a crucial component in assessing whether natural selection can rescue a population from extinction. We provide theoretical evidence for the importance of recognizing evolution in predicting and explaining successful biological invasions.
Keywords: adaptive evolution, Allee effects, biological invasion, reaction-diffusion equation
Introduction
While advances in free trade and globalization increase the movement and accelerate the accumulation of invasive species (Lockwood et al. 2005), it is still unclear how introduced populations can successfully establish. As Elton (1958) pointed out, for every successful invasion that occurs, ‘there are enormously more invasions that never happen, or fail quite soon or even after a good many years’ (page 109). What then determines success and failure? This modern biological paradox cannot readily be reconciled, especially in the characteristic case where the founder population is small, as ‘such populations are definitely in a precarious position’ (Mayr 1965; page 42). Introductions of populations at low density and/or small size are often faced with inverse density dependent effects, attributed to demographic stochasticity or reduced cooperative interactions (Courchamp et al. 1999). Allee (1931) first proposed that under these conditions, populations may suffer a decrease in the per-capita rate of increase, from here on referred to as the Allee effect.
Upon arrival in a novel environment, individuals need to overcome a series of challenges in order to reduce the population's risk of extinction. The time period in which this occurs is generally considered the initial establishment phase, and is thought to be a common feature and general pattern of invasion and the process of growth and expansion (Shigesada and Kawasaki 1997; Sakai et al. 2001). The occurrence of a lag phase that precedes a noticeable increase in population growth and density can result from ecological and/or evolutionary phenomena (Sakai et al. 2001). Small populations that undergo logistic growth slowly increase through the initial phase of the exponential curve, leading to the perception of a time lag. Where this time lag is more pronounced, populations may be recovering from inverse density dependent effects (i.e., Allee effects).
Individuals may suffer a reduction in fitness at low densities for many reasons (reviewed by Courchamp et al. 2008). Even when the initial population is large, rapid dispersal required for expansion could be suicidal as the population density decreases, thereby enhancing inverse density dependent effects (Lewis and Kareiva 1993; Drake et al. 2005; Drury et al. 2007). Commonly, a distinction is made between a ‘demographic’ Allee effect and a ‘component’ Allee effect where the population size and density affects the mean overall fitness or some component of individual fitness, respectively (Stephens et al. 1999; Gascoigne et al. 2009). It is often difficult to decipher the exact mechanism that manifests Allee effects (and it is not always the case that component Allee effects lead to demographic Allee effects). Nevertheless, we focus our attention on population level demographic Allee effects with the underlying assumption that a component Allee effect led to the demographic Allee effect. Essentially, it is the case that introduced individuals may be maladapted to small population sizes where their survival and reproductive ability are significantly impacted, and these impacts on individual fitness combine to produce an overall decrease in abundance (i.e., demographic Allee effects). As Allee effects impact individual fitness, the underlying traits that influence these effects (i.e., component Allee effects) may be subject to natural selection (Courchamp et al. 2008).
Whereas propagule pressure is an emerging explanation for the establishment of invasive species (Lockwood et al. 2005), it relies on an obvious relationship between the number and size of introduction events and the probability of success, as safety in numbers helps combat Allee effects and stochastic extinction. In the event that propagule size is not large enough to overcome inverse density dependent effects, a population may still become established if individuals can adapt to mitigate or avoid Allee effects. Traits that may be responsible for reproductive success and survival at small population densities and sizes include mate finding cues (pheromones and vocal/visual signals), dispersal/aggregation behavior, habitat preferences, mating synchronicity, and gamete morphology and performance (see Courchamp et al. 2008 for detailed studies). Direct evidence for the evolution of these traits as functional adaptations to Allee effects is limited, but we can infer an adaptive evolutionary origin of these traits from studies of sexual selection (Courchamp et al. 2008; Gascoigne et al. 2009).
An evolutionary response to intensive selection pressure imposed by density dependent survival and reproduction relies on genetic variants for adaptive evolution. According to neutral quantitative genetic theory, a loss of genetic variation is expected from population bottlenecks and founder effects (Nei et al. 1975). However it is not neutral variation that matters, but rather evolvability depends on the variation relevant to selection. Maintenance or even increases of this (additive) genetic variation has been theoretically and empirically observed following a bottleneck or in small founder populations (Bryant et al. 1986; Goodnight 1988; Willis and Orr 1993; Cheverud and Routman 1996; Briggs and Goldman 2006; Turelli and Barton 2006). Additionally, many recent studies suggest that there is actually no significant reduction in genetic diversity in most successful invaders (Lee 2002; Allendorf and Lundquist 2003; Roman and Darling 2007 and references therein), and that evolution can occur on contemporary timescales (Reznick and Ghalambor 2001; Carroll et al. 2007; Kinnison and Hairston 2007). Our purpose here is to explore the feasibility of small populations that may adaptively respond to overcome Allee effects in order to establish, given any amount of genetic variation.
In this paper, we present evidence for the enhanced potential for growth and spread of a small introduced population of organisms faced with Allee effects when adaptation occurs. We model the growth and spread of this population according to a reaction-diffusion equation, and allow evolution to influence inverse density dependent effects through a genetic subsystem that provides the opportunity for successful invasion when otherwise (under the current, ecological paradigm) the population would go extinct.
Model description
The deterministic model that we explore in this paper broadly describes population dynamics with density-mediated growth (i.e., an Allee effect) and diffusive dispersal. This type of demographic model has been used as a compact and tractable representation of invasion (e.g., Skellam 1951; Lewis and Kareiva 1993). Specifically, it has been applied to systems such as introductions of nonindigenous freshwater and marine species through ballast water discharge (Drake et al. 2005; Drury et al. 2007). Using this approach, Drake et al. (2005) report acceptable volumes of discharge for various organisms (with differing reproductive rates) for a range of invasion risk tolerances. Here, we consider populations that are introduced below the invasion risk threshold, but nonetheless succeed if evolutionary dynamics are considered in conjunction with the ecological system.
The ecological system
The growth and spread of an introduced population of organisms is represented by a reaction-diffusion equation described by (Lewis and Kareiva 1993):
| (1) |
which determines the rate of change in the local population density relative to the carrying capacity [u which denotes u(x,t)] over time, at a point in space. This equation models the growth of the population (the first term on the right hand side of Eqn. 1) at a spatial location that is subject to an Allee effect in addition to migration (which depends on the second term on the rhs of Eqn. 1). The diffusion coefficient (D) scales the rate of population spread, in this case across a one-dimensional habitat x. The reproductive rate is regulated by r, and a2 (which is a function of space and time, derived below) is the local critical density or Allee threshold that determines if population growth is positive or negative (Fig. 1A).
Figure 1.
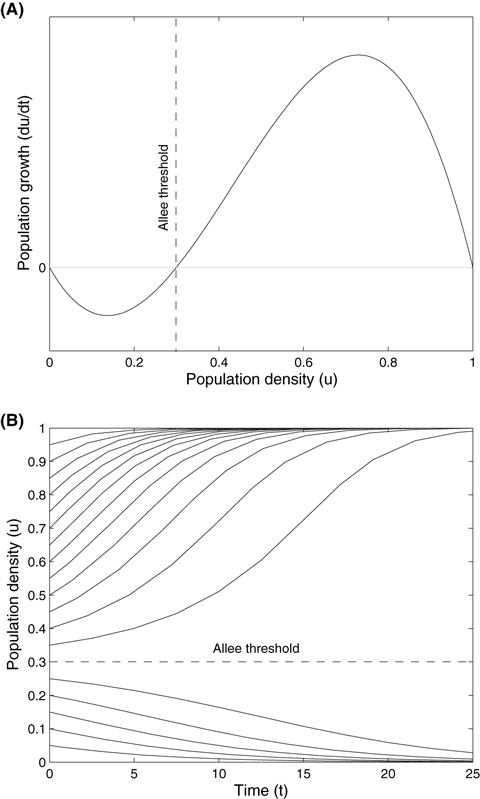
Growth dynamics of the model population (A), and the solution of Eqn. (1) without diffusion (B) with reproductive rate, r = 0.6, and Allee threshold, a2 = 0.3. Trajectories for population size (B) are given for initial densities from 0 to 1 in increments of 0.05.
There are many variations of single-species models of population dynamics that exhibit Allee effects (see table 1 of Boukal and Berec 2002), however the growth function of Eqn. (1) is widely used and flexible (Boukal and Berec 2002). The behavior of this Verhulst (1838) logistic model modified to include a nonlinear cubic term (based on the Fitzhugh-Nagumo equations; Fitzhugh 1960; Nagumo et al. 1962), is bistable, with equilibria at u = 0 (extinction), u = a2 (unstable threshold), and u = 1 (carrying capacity). Figure 1A shows this behavior in terms of the growth of the population (change in population density with respect to time) versus the population density. At densities below the critical threshold (a2) there is negative population growth declining to extinction (from here on the population is considered extinct below a cutoff density of 0.0001, as a declining population trajectory will only asymptotically approach zero in a deterministic model; Gomulkiewicz and Holt 1995); otherwise the population will reach carrying capacity. This is clearly shown in Fig. 1B with the graph of the solution of the growth function (population size versus time) at various initial densities.
When diffusion is added to this model of population growth, there are two critical elements that emerge based on the solution to the partial differential equation (PDE). The first is the wave speed, which is determined by the Allee threshold (a2). As we are considering a strong Allee effect in this model (i.e., 0 < a2 < 1, where the population below this threshold exhibits negative growth versus reduced positive growth from a weak Allee effect), there exists a unique wave speed of the invasion front that is a result of being ‘pushed’ from the inside out, as opposed to being ‘pulled’ by the leading edge (Lewis and Kareiva 1993). This velocity can be derived through the solution of the PDE (1): 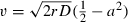 (Lewis and Kareiva 1993; Murray 1993). This result suggests that in order for a wave to maintain a positive velocity of advance, the magnitude of the Allee threshold (a2) must be less than half of the maximum value of the population density relative to the carrying capacity. In addition to the velocity of the wave front, the region occupied by the invading population must exceed a certain critical size for positive growth to occur (Skellam 1951; Kierstead and Slobodkin 1953). This phenomenon is clearly explained by Okubo (1980) by noting that whereas reproduction takes place within a region or patch, diffusion takes place at the boundaries resulting in a loss of organisms, thus reducing the density within the patch. This tradeoff in the ratio of inner region volume to outer surface area will either allow a population to grow or decline with an inverse relationship of diffusivity to rate of growth. This relationship gives a minimum region within which reproduction cannot compensate for loss due to diffusion, especially when Allee effects influence population growth. Thus, Lewis and Kareiva (1993) derive a minimum size condition (i.e., the radius of the initial beachhead) based on the wave speed that is required for the population to establish and radially expand. We address this critical size threshold qualitatively, as the analytical solution (i.e.,
(Lewis and Kareiva 1993; Murray 1993). This result suggests that in order for a wave to maintain a positive velocity of advance, the magnitude of the Allee threshold (a2) must be less than half of the maximum value of the population density relative to the carrying capacity. In addition to the velocity of the wave front, the region occupied by the invading population must exceed a certain critical size for positive growth to occur (Skellam 1951; Kierstead and Slobodkin 1953). This phenomenon is clearly explained by Okubo (1980) by noting that whereas reproduction takes place within a region or patch, diffusion takes place at the boundaries resulting in a loss of organisms, thus reducing the density within the patch. This tradeoff in the ratio of inner region volume to outer surface area will either allow a population to grow or decline with an inverse relationship of diffusivity to rate of growth. This relationship gives a minimum region within which reproduction cannot compensate for loss due to diffusion, especially when Allee effects influence population growth. Thus, Lewis and Kareiva (1993) derive a minimum size condition (i.e., the radius of the initial beachhead) based on the wave speed that is required for the population to establish and radially expand. We address this critical size threshold qualitatively, as the analytical solution (i.e., 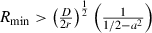 ; Lewis and Kareiva 1993) is for two-dimensional spread, while we work with a simpler one-dimensional model. The minimum critical radius is proportional to the ratio of diffusivity (i.e., diffusion coefficient, D) to the reproductive rate (controlled by r). The inclusion of diffusion in the model provides a spatially explicit understanding of how all of the components interact to affect invasion/establishment success. The diffusion process has been extensively analyzed in invasion processes (e.g., Fisher 1937; Skellam 1951; Okubo 1980).
; Lewis and Kareiva 1993) is for two-dimensional spread, while we work with a simpler one-dimensional model. The minimum critical radius is proportional to the ratio of diffusivity (i.e., diffusion coefficient, D) to the reproductive rate (controlled by r). The inclusion of diffusion in the model provides a spatially explicit understanding of how all of the components interact to affect invasion/establishment success. The diffusion process has been extensively analyzed in invasion processes (e.g., Fisher 1937; Skellam 1951; Okubo 1980).
The evolutionary subsystem
In order to incorporate evolutionary factors that may influence invasion success, we develop a quantitative genetic subsystem. This genetic subsystem is coupled to the ecological model to explore the effects of selection and genetic variance on traits that may increase a population's likelihood of survival. Specifically, we allow the Allee threshold to become a dynamic parameter that is considered to be a fitness related trait (e.g., a trait impacting the component Allee effect). From here on, except in the absence of evolution, referring to the Allee threshold implies that that value is the initial value, as it changes over time. This quantitative trait influences an organism's ability to survive and reproduce in a small population. The results reveal the possibility that an introduced population that would fail to persist in the ecological context of this model has the potential to succeed through evolutionary means. Including evolution within the context of ecological invasions can serve to provide more robust predictions for management strategies. Therefore, it is important to investigate the possibility of evolution in the analysis of invasions.
The framework that is used to link evolutionary change with ecological processes involves developing a relationship between the fast, ecological and slow, evolutionary timescales in order to make these rates comparable (Kondrashov and Khibnik 1996). In the coupled evolutionary ecology model, the reaction-diffusion Eqn. (1) describes the change in the population density over time and is tied into a genetic subsystem that allows the organismal response to population density to evolve in terms of the selection gradient and genetic variance. As the population dynamics vary across space, the genetic subsystem describes the rate of change of the trait mean (i.e., the Allee parameter) at each location x by:
| (2) |
(Pease et al. 1989; García-Ramos and Kirkpatrick 1997; Kirkpatrick and Barton 1997; Hare et al. 2005). The first term on the right hand side reflects the force of local directional selection, where the selection gradient for frequency-independent selection is the rate of change of the mean Malthusian fitness function (i.e., per-capita growth rate: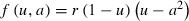 ) with respect to the trait, a (Lande 1976; Falconer 1989). Thus,
) with respect to the trait, a (Lande 1976; Falconer 1989). Thus,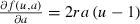 , where we assume that individual fitness approaches the population mean fitness, as most individuals are close to the average phenotype (Webb 2003). This suggests that the genetic variance (ε) is small (and constant in this model). This small parameter for the genetic variance can be used to couple the fast ecological timescale, t, with the slow evolutionary timescale, τ = εt (Kondrashov and Khibnik 1996; Webb 2003). Combing these two components of genetic variance and selection, quantifies the effect of natural selection on the local mean value of the quantitative trait (the Allee parameter; Lande 1976; Falconer 1989).
, where we assume that individual fitness approaches the population mean fitness, as most individuals are close to the average phenotype (Webb 2003). This suggests that the genetic variance (ε) is small (and constant in this model). This small parameter for the genetic variance can be used to couple the fast ecological timescale, t, with the slow evolutionary timescale, τ = εt (Kondrashov and Khibnik 1996; Webb 2003). Combing these two components of genetic variance and selection, quantifies the effect of natural selection on the local mean value of the quantitative trait (the Allee parameter; Lande 1976; Falconer 1989).
In order to account for the influence of migration on the trait's local mean, the latter two terms in Eqn. (2) incorporate space. The middle term takes into account asymmetrical gene flow caused by the variation of density across space (Pease et al. 1989; García-Ramos and Kirkpatrick 1997; Kirkpatrick and Barton 1997; Hare et al. 2005). This captures the influence of the mean trait value (i.e., genetic contribution) from more abundant populations to less abundant neighboring locations due to the spatial gradient, as more individuals migrate from areas with relatively high population densities. The last term mirrors the diffusion term from the ecological model, and describes the homogenizing effect of random dispersal.
We solved the spatially explicit system numerically using Matlab 7.0 (2004, The MathWorks, Natick, MA, USA) using a finite difference method to incorporate diffusion and gene flow (adapted from Garvie 2007). By iterating Eqns (1) and (2) forward in time, the population density and Allee threshold at each location are updated with diffusion following growth and selection, respectively, while incorporating the spatial gradient. The simulated populations, with and without evolution, behaved as we expected from the model Eqns (1) and (2), and adequately approximate/represent the critical conditions that govern this dynamical system.
Results
The dynamics of the evolutionary ecology model can be interpreted using the idea of fast and slow timescales (Kondrashov and Khibnik 1996; Webb 2003). Earlier, we assumed that the genetic variation (ε) was small (to use mean fitness as a proxy for individual fitness), which can subsequently be taken advantage of for our analysis of the coupled evolutionary ecological dynamics. When ε = 0, the situation without evolution, the genetic subsystem is frozen and the population moves towards a stable equilibrium of the ecological subsystem (carrying capacity or extinction) depending on its initial density (greater than or less than the Allee threshold respectively; Fig. 1B) and initial radius (spatial extent) in the spatially explicit model. When ε > 0 but small, the Allee threshold evolves relatively slowly and influences the ecological system. Whenever the population is below its carrying capacity (u = 1 for each spatial coordinate x when space is explicit), Eqn. (2) is negative, and decreases the mean Allee threshold (a2), as the intensity of selection is density dependent. Thus, fitness increases as Allee effects are suppressed, and selection drives the Allee threshold towards zero. If the population density is greater than the Allee threshold, but still below the carrying capacity, it will progress towards carrying capacity more rapidly than it would without evolution as a2 decreases; as the rate at which the population density changes (Eqn. 1) is proportional to the difference between u and a2. The ecological dynamics are reversed when the population density is below the Allee threshold as the population declines towards extinction, but more slowly than it does without evolution. When u < a2, Eqn. (1) is negative, and the population density approaches extinction more rapidly with a2 constant (as the difference between u and a2 increases), than it does with evolution as a2 decreases (revealing a more pronounced time lag to extinction). During this time lag, as the population slowly declines, the opportunity for evolution to overcome inverse density dependent effects occurs. If the rate of evolution is fast enough, the Allee threshold can fall below the population density, causing the rate of change of population density to become positive (where u > a2) and the population grows and can successfully invade. The chance that evolution can rescue the population from extinction depends on the relative rates of genetic change in the quantitative trait (i.e., Allee threshold) and of population decline (Gomulkiewicz and Holt 1995); hence the amount of genetic variance greatly impacts the ability to adapt and survive.
A nonspatial example of this process, referred to as evolutionary rescue (Gomulkiewicz and Holt 1995), is shown in Fig. 2, where a population is introduced below the Allee threshold. Without evolution, the population declines to extinction (Fig. 2A, solid line) as the Allee threshold remains constant (Fig. 2B, solid line). When the population can evolve (Fig. 2, dotted line), it declines at first until it can overcome the magnitude of inverse density dependence, and is then able to successfully establish. As it is difficult to measure the Allee effect empirically (Tobin et al. 2007), we use an extreme value that exaggerates density dependent effects in order to investigate the ‘worst case scenario’ (a2 = 0.3, where the population exhibits deterministic decline when its density is less than 30% of its carrying capacity). When evolution is included, we used a small value for the genetic variance, ε = 0.02, in order to remain consistent with fast-slow dynamics, unless otherwise indicated.
Figure 2.
Comparison of an invading population introduced at a density below the Allee threshold, a2 = 0.3 (u = 0.25, r = 0.6). The solid line represents the nonspatial system (D = 0) described by Eqns (1) and (2) without evolution (ε = 0) which results in extinction (A) and a constant Allee threshold (B). The dotted line indicates population growth (A) when evolution (ε = 0.02) acts to reduce the Allee threshold (B).
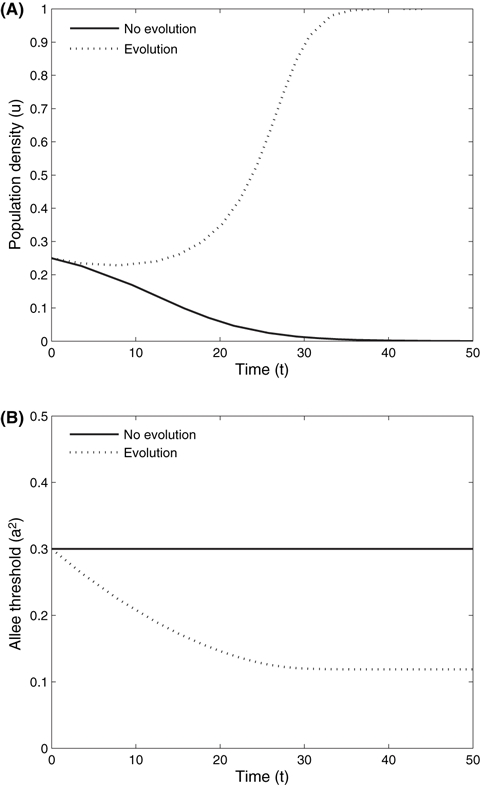
In general, there is a range of parameter space that permits persistence for a population below the Allee threshold in the nonspatial model with evolution (instead of simple decline to extinction). We explored this behavior while varying the genetic variance from zero to 0.1. As genetic variance increases, we are essentially relaxing the assumption of fast-slow timescales and allow evolution to occur more rapidly. These dynamics are shown in Fig. 3 where initial population densities below the Allee threshold require a minimum amount of genetic variance in order to avoid extinction. In this case, the rate of reproduction, r, also influences the potential for evolutionary rescue, as it impacts both population growth and rate of evolution (Eqns 1 and 2, respectively). As we relax the assumption of fast-slow timescales, the behavior remains qualitatively the same as that described analytically under a strict fast-slow timescale assumption.
Figure 3.
Parameter combinations of reproductive rate: r; genetic variance: ε, and initial population density: u, that result in extinction or evolutionary rescue. In this nonspatial scenario, initial population densities greater than the Allee threshold (a2 = 0.3) always succeed, thus the focus is on the parameter space that allows for evolutionary rescue (i.e., where the population growth changes from negative to positive). As the reproductive rate increases from 0.1 to 1, there is less genetic variance needed for a population to evolve to overcome inverse density dependence as increased reproduction will contribute to suppressing Allee effects.
Including more realistic population dynamics through spatial structure provides further invasion criteria. Nonetheless, the additional complexities result in qualitatively similar behavior to the nonspatial model. In this case, not only will evolution influence population growth, it affects the wave speed and the critical size threshold, Rmin. As the population overcomes Allee effects with a decreasing Allee threshold, the wave speed accelerates and the critical patch size becomes smaller. Thus, in addition to the initial density of the introduced population and the genetic variance, the initial radius or patch size of the initial invasion area, the ratio of diffusion to reproduction, and gene flow will factor into successful establishment and give rise to a wider range of interactions between the ecology and evolution of this system.
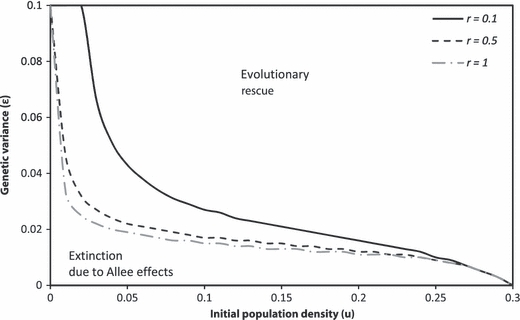
The numerical solution of Eqn. (1) (without the evolutionary subsystem) in one-dimensional space, with an initial population density below the Allee threshold, declines to extinction (Fig. 4A). This is contrasted by the results when the evolutionary subsystem is included. With the initial population density below the Allee threshold, Fig. 4B shows that the population rebounds from decline. The same type of rescue occurs for a population that starts near carrying capacity, but occupies an initial spatial size below that which is necessary for a population to successfully establish. Figure 5A shows a rapidly declining population that goes extinct. Under the same circumstance, but where evolution of the Allee threshold occurs, Fig. 5B shows the population density at first beginning to shrink and then growing and expanding. In addition to the time evolution of population density across space in Figs 4 and 5, the evolution of the mean trait value across space illustrates how gene flow and the density dependent selection gradient influences its rate of change and distribution (Figs 4C and 5C). As the intensity of selection is density dependent (and we assume constant genetic variance), locations with smaller populations can evolve the trait value more rapidly compared to other areas where Allee effects may not be as strong and experience weaker selection. The trait distribution over time, Figs 4C and 5C, therefore reflect the population density distribution, but are also influenced by the trait values of the migrants. As individuals disperse out to new locations and push the boundaries of the species range, their trait values are averaged to determine the demographic Allee threshold for that spatial coordinate. This demographic Allee threshold combines with their local population density to influence individual fitness and population growth (where the distance between the density and mean trait value is the initial degree of maladaptation).
Figure 4.
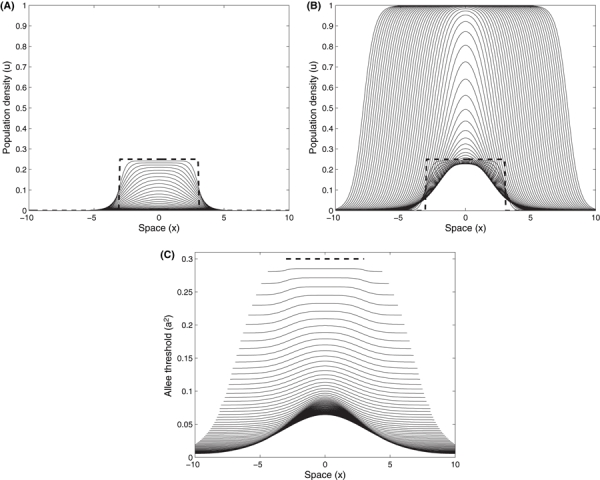
Diffusive dispersal of an introduced population at an initial density (bold dashed line) below the initial Allee threshold, a2 = 0.3 (u = 0.25, r = 1, D = 0.1) across a linear, one dimensional habitat. The population collapses over time to extinction (A) where there is no evolution (ε = 0), and succeeds (B) after an initial decline with evolution (ε = 0.02). (C) The evolution of the mean value of the Allee threshold across space (where the initial distribution is given by the bold dashed line). The population density distribution and corresponding trait values (i.e., Allee threshold) are plotted at equal time increments (every 20 of 1200 model iterations).
Figure 5.
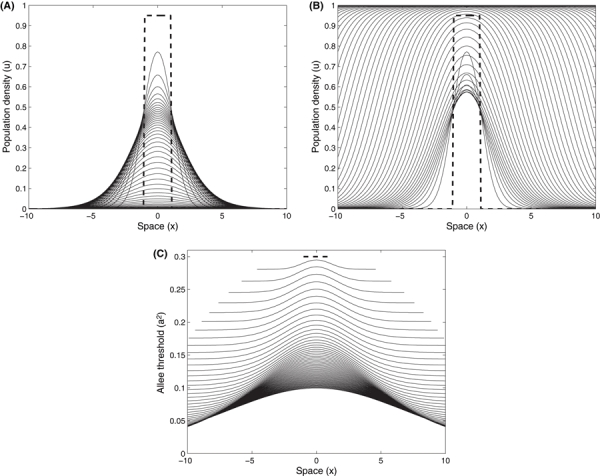
Population density of a diffusion dispersed population across one dimensional space. The initial population density (bold dashed line) is near carrying capacity (u = 0.95, a2 = 0.3, r = 1, D = 0.5), but introduced below the minimum radius of area determined to be critical for invasion success. (A) is collapsing to extinction without evolution (ε = 0), whereas (B) shows success of an invader with evolution (ε = 0.02) after initial decline. (C) The evolution of the mean value of the Allee threshold across space (where the initial distribution is given by the bold dashed line). The population density distribution and corresponding trait values (i.e., Allee threshold) are plotted at equal time increments (every 20 of 1200 model iterations).
We explored when evolutionary rescue occurred across a range of parameter values for the spatially explicit model. According to Drake et al. (2005), variability among locations and over time makes it unreasonable to determine precise estimates for the diffusion coefficient, D. We therefore explored a range of values, and present those that best illustrate breadth of behavior. The parameter that controls the reproductive rate, r, was also varied substantially, but as the spatial dynamics depend on the ratio of diffusion to rate of reproduction (resulting in a measure of length); we fixed r and varied D, unless otherwise noted. This was justified as the results of the spatial simulations are qualitatively identical for equivalent ratios. The effects of the critical patch size, initial population density, ratio of diffusion to growth and genetic variation on evolutionary rescue and population dynamics are shown in Fig. 6.
Figure 6.
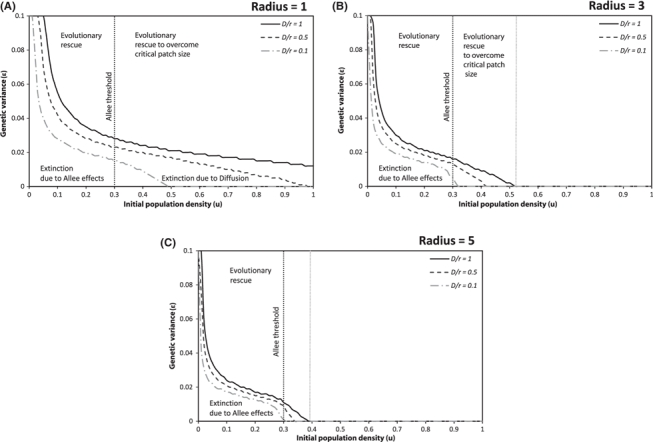
The sensitivity of population growth and expansion based on the combination of parameter values. The ratio of the diffusion coefficient (D) to the reproductive rate (r) determines whether the population will expand or collapse according to the initial radius of the introduced population. The areas under the curves denote combinations of genetic variance and initial population density that result in extinction. Areas above the curves are combinations of genetic and/or demographic conditions that produce inevitable persistence. The parameter space between the vertical dashed lines refers to the different ways population survival is influenced. To the left of the initial Allee threshold, the initial population density will either go extinct due to density dependent effects (below the D/r curve), or given enough genetic variation, will be evolutionarily rescued (above the D/r curve). The area to the right of the initial Allee threshold [and between the dashed lines in (B) and (C)] is the case where the initial population density is greater than the Allee threshold but due to the initial spatial size and the ratio of diffusion to reproduction, the population may go extinct without sufficient genetic variance (below the D/r curve), otherwise it will evolve to overcome the critical patch size effect. For initial population densities greater than the rightmost dotted line, populations persist solely because of ecological effects. Thus, the area between the dashed lines in (B) and (C) truly delineates evolutionary rescue when D/r = 1. The rightmost vertical line moves slightly to the left to the point of intersection of the D/r curve and the x-axis for other values of D/r. Graphs (A), (B), and (C) represent different radii of the linear habitat that the introduced population initially occupies.
When the size of the initial population is too small (i.e., a radius of 1), a population at carrying capacity (i.e., u = 1) will go extinct without evolution due to the relative effect of diffusion to reproduction (Fig. 6A). If evolution occurs rapidly enough (i.e., ε > 0.02), the population can overcome inverse density dependent effects and compensate for the loss due to diffusion and rebound from low densities. When the initial radius of the population is increased (Fig. 6B,C), the chance of survival and establishment (growth and expansion) of populations above or below the Allee threshold increases with initial density and genetic variance. Therefore, the initial radius of the population can significantly impact the likelihood of evolutionary rescue for populations with the same amount of genetic variance.
This is demonstrated further in Fig. 7A, where the rate of recovery (i.e., the inverse of the time lag before growth becomes positive and the population reaches carrying capacity) for a population near carrying capacity depends on its initial size/radius and genetic variance. Where size and variance are small, rescue never occurs. As these parameters increase, the rate of recovery gradually becomes faster until it essentially plateaus (although with greater variance and initial radius, the rate of recovery may slow slightly if the initial spatial extent is large enough for the population to experience early growth before diffusion causes decline prior to recovery). If the population occupies a large enough spatial extent, it will succeed without evolution (where the genetic variance is zero), however the lag time may be more pronounced depending on the ratio of diffusion to reproduction through the tradeoff between growth and spread (e.g., if spread is relatively fast compared to reproduction, D/r = 1). The population density may thus initially decline across space until reproduction can sufficiently overcome the loss due to diffusion, and the population can grow to carrying capacity. Similar to the nonspatial case, a population (greater than the Allee threshold) above the spatial threshold will grow to carrying capacity more rapidly with evolution than without.
Figure 7.
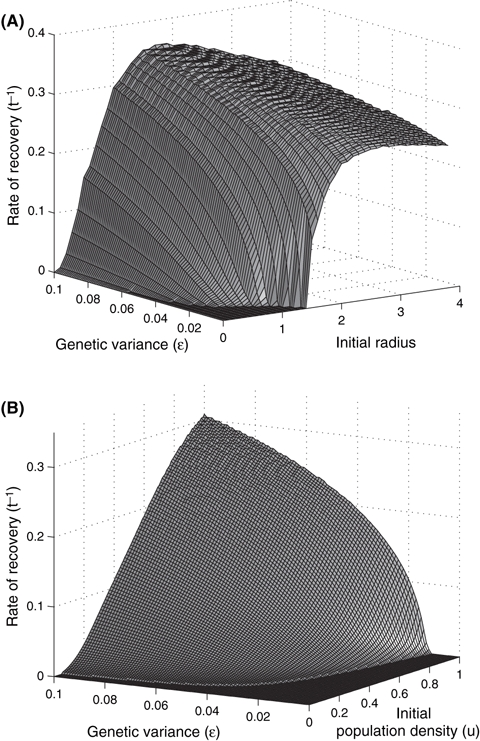
Rate of recovery in terms of the inverse of the time lag before population growth becomes positive, where one ‘timestep’ equals 24 iterations of the model. In (A), the initial population density is near carrying capacity (u = 0.95, a2 = 0.3, D/r = 1), and the initial radius and genetic variance, ε, varies. Where the rate of recovery is zero, the population goes extinct as it initially occupies an area smaller than the critical patch size (in this case, a radius of 1.4) or does not have sufficient genetic variance to evolve quickly enough to be rescued prior to extinction. Increasing the genetic variance and initial radius will decrease this time lag until the population no longer experiences any negative growth (in this case, for initial radii ≥ 3.8 and ε ≥ 0.036; for initial radii > 2.7, the rate of recovery slows slightly due to early growth followed by a transient decline that precedes ultimate recovery). When the initial population density varies (indicating the initial degree of maladaptation where a2 = 0.3), (B) shows the rate of recovery with the initial radius fixed (as in Fig. 5A where the radius = 1 and D/r = 1). In this case, extinction will occur without evolution not only for an initial density below the initial Allee threshold, but for any density as the initial radius is below the critical patch size. Hence, a nonzero rate implies evolutionary rescue and a zero rate means extinction.
As shown in Fig. 7B, when evolutionary rescue is possible, the initial level of maladaptation (a02−u0) and the genetic variance (ε) also determine the rate at which evolutionary rescue proceeds. Figure 7B uses parameters (i.e., radius and ratio of D to r) for a population that would decline and go extinct without evolution regardless of the initial density. Hence, it is clear that the amount of time required for a population to begin growing depends on its initial level of maladaptation (to both the critical density and spatial thresholds) and/or genetic variance. As the rate at which this rescue occurs depends on the amount of genetic variance (Eqn. 2), it may take an extremely long time (as ε→ 0, the rate of recovery → 0) for the Allee threshold to fall below the population density. In this circumstance, as the population density becomes very close to zero, the rate of change of the Allee threshold is greater than that of the population density (as u → 0, ∂u/∂t→ 0 and ∂a/∂t → -2εra). Thus, theoretically, rescue would always occur (Gomulkiewicz and Holt 1995). However, to maintain biological realism, when solving this system numerically, we always considered the population extinct when the maximum density (across space, when diffusion is included) becomes reasonably close to zero (i.e., u = 0.0001; we chose this protocol instead of the total population across space due to the diffusion dynamics based on the Gaussian dispersal kernel and the ‘pushed’ wave front behavior).
Overall, the numerical results qualitatively hold for a wide range of dimensional parameter values and initial conditions with and without diffusion and in one- and two-dimensional space. Results for two-dimensional space are not shown as they are qualitatively similar to the simpler, one dimensional model.
Discussion
From these results, it is apparent that adaptations that enable organisms to overcome the negative effects of low densities can allow the population to rebound from a trajectory toward extinction to grow to reach carrying capacity. Current management strategies (e.g., reducing population density or size) are based on ecological theory (e.g. Drake et al. 2005), but this evolutionary ecology model suggests that adaptive evolution can enable successful establishment and that ecological considerations alone may not be sufficient.
Under the assumptions of an Allee effect and diffusive dispersal, the idea of ecological size thresholds fits well with the ecological evidence that a large founding population is a primary cause of successful establishment (Lockwood et al. 2005; Colautti et al. 2006). However, by incorporating evolution, we see that the situation is not quite this simple because ecological size thresholds and genetic variance can interact to determine successful establishment. As the ratio of diffusion to reproduction decreases, the spatial constraint on population growth becomes weaker, and less genetic variance is needed to rescue populations with densities below the Allee threshold. As the initial spatial radius of introduction increases, population persistence is more likely with less genetic variance for selection to act on. Furthermore, the rate of this rescue depends on the initial genetic load or maladaptation (i.e., how far the population density is from the Allee threshold), as well as the amount of genetic variance. Because bottlenecks during founding events do not always result in highly reduced genetic variability, even small founding populations may have sufficient genetic variation to evolve to overcome Allee effects and establish, contrary to solely ecologically based predictions.
Additionally, we can draw several general insights about how dispersal impacts selection and evolution of Allee effects in an invasion context. As species are transported from their native environment into novel habitats or simply disperse on their own, it is clear that the genetic composition of the local population can influence the rate of evolution and adaptation to the new local conditions. Given enough genetic diversity, local populations can adapt to their local environment, but dispersal may hinder survival across ecological clines as dispersers tend to be maladapted to the new local environment. Essentially, local population persistence depends on the race between the rate of evolution and the degree of maladaptation (Gomulkiewicz and Holt 1995). In this case, gene flow will play a major role in determining the outcome. As Kirkpatrick and Barton (1997) and García-Ramos and Kirkpatrick (1997) demonstrate, individuals moving from one selection regime from the center of their species’ range to the periphery can introduce enough maladaptation that the new area becomes a sink environment. On the other hand, Holt et al. (2003, 2004) show that immigration can have a positive influence on adaptation to sink environments, in some circumstances. Resolving the disparity between these perspectives requires understanding what is contributing to the severity of maladaptation and the population's ability to overcome it.
In our model, dispersal impacts survival ecologically through the critical patch size, and genetically, as individuals may move from areas where they are well adapted (i.e., the population density is greater than the Allee threshold or mean trait value) to sink regions, where they are maladapted. As individuals disperse across space, they may be contributing positively in an ecological sense to the quality of their new local environment (by increasing the local population density). However, dispersers are more likely to come from higher density areas where Allee effects, and hence selection, are locally weak. These dispersers potentially introduce more maladaptation to their new location, because they increase the average phenotype (Allee threshold) in the new location where density is likely to be lower.
Interestingly, the evolutionary impacts of migration in this model do not dramatically influence the dynamics. Changes to the local mean phenotype through local selection and simple mixing (i.e., diffusion) actually slightly hastens the evolutionary rescue effect over a model that considers only the impact of local selection. As the selection intensity is density dependent and proportional to u− 1 for each point in space, the peripheral individuals faced with stronger selection with lower trait values have a small positive influence on the more dense neighboring populations. The gradient term accounts for asymmetric gene flow due to differential migration from areas of relatively high population densities. However, this term does not alter the overall evolutionary dynamics based on local selection any more than adding the diffusion term, as the negative effects of gene flow and the local rate of evolution (which is relatively fast, based on the selection intensity) essentially cancel each other out. In this context, similar to that of Holt et al. (2003, 2004), the immigrants simply contribute to the local population density, which helps prevent extinction long enough for evolutionary rescue to occur locally (i.e., positive population growth; note that whereas Holt et al. (2003) attribute the main effect of immigration to the contribution of variation, this is not the case in our model, as we assume constant genetic variance). Overall, our results are similar to that of Holt et al. (2004) where immigration has a demographic effect on increasing fitness that can essentially outweigh the ‘swamping’ effect of gene flow.
In general, the primary determinant of invasion success depends on positive population growth at the center of the introduced range. This result comes from the Allee effect [and the solution to the PDE (1)] by forcing a ‘pushed’ travelling wave front (Lewis and Kareiva 1993), where the wave speed causes population expansion, contraction, or propagation failure (i.e., pinning; Keitt et al. 2001). Intuitively, aggregation-like behavior emerges based on the strength of the Allee effects. Individuals that disperse too far from the whole are likely to die before they can ‘pull’ others in their vicinity. In this regard, growth occurs from the inside out, where the population seemingly spills out and overflows to expand its range. Consequently, in this study, and for biological invasions that exhibit similar dynamics, it is more important to focus on the center of the invader's range and whether the initial beachhead can survive (through evolutionary rescue), than the fate of peripheral populations at the wave front when determining the importance of evolution on invasion success. This is also understood by comparing the nonspatial (Fig. 3) with the spatial (Fig. 5) sensitivity analysis. The overall dynamics are qualitatively similar in the parameter space that allows for evolutionary rescue to occur.
Even though gene flow and spatial structure do not dramatically influence the establishment of an introduced population, additional invasion criteria need to be considered. When analyzing the model behavior in a spatially explicit context, there is an ecological tradeoff between growth and spread that affects establishment and the rate of recovery. As previously mentioned, reproduction needs to compensate for the loss due to diffusion. Including evolution and suppressing Allee effects, actually contributes to the acceleration of the wave front (i.e., enhancing dispersal speed). A population then can more rapidly disperse as it evolves, and may become more of an invasion threat as long as this range expansion does not reduce their density too quickly. Whereas this increasing wave speed can lead to a slightly longer lag phase prior to positive population growth, the population will likely be inevitably rescued because this effect primarily influences the dynamics at the periphery and is offset by the reduction in the critical invasion area (Rmin). Although there is no range contraction (as there is always a positive wave velocity with unbounded expansion due to the parameter values and absence of environmental heterogeneity or range limitations; Filin et al. 2008), as the critical patch size (Rmin) becomes smaller with the decreasing Allee threshold, rescue occurs more readily at the range center as the critical patch size threshold criteria weakens and the behavior approaches that of the nonspatial model. This may seem like an oversimplification of the global dynamics; however these conclusions are valid in the context of this investigation which focuses on the establishment phase rather than subsequent range expansion and spread.
Recognizing that evolution can significantly affect the establishment success of invasive species is becoming more widely accepted, influencing the ways in which invasion biologists conduct their research (see the other articles in this issue). Specifically, adaptations that diminish Allee effects and evolutionary responses to density dependence are beginning to emerge as viable explanations for sustaining vulnerable populations at low density and size (Gascoigne et al. 2009). As it is difficult to conclusively support this claim empirically (as the origin of the adaptation or the associated cost may be unknown; Courchamp et al. 2008; Gascoigne et al. 2009), mathematical models that incorporate evolution and compare the effects of various strategies (e.g., mitigating component Allee effects) can help decipher the mechanisms that both limit and facilitate population growth. Two such models that incorporate adaptations to component mate-finding Allee effects compare the efficiency and survival of populations at various densities that attract mates with or without a sexual pheromone (Jonsson et al. 2003) or by increasing mate detection distance (Berec et al. 2001). Another study suggests that broadcast spawners that evolve their gamete morphology and performance under sperm limitation (at low density) bear a cost of decreased fitness at high density due to hybridization and competition (Levitan 2002). In these cases, particular strategies are shown to influence population viability in addition to an associated tradeoff, whereas our investigation provides broad, albeit simplistic, results dealing with generalized demographic Allee effects and evolution.
In order to understand how the results of this simplistic model extend to more realistic and complex evolutionary scenarios, spatially explicit, individually-based stochastic simulation of the introduced populations should be developed to investigate more closely the mechanisms that allow these population level dynamics to emerge. In particular, tracking the mean value of a component Allee effect is sufficient to illustrate how evolution can overcome inverse density dependence and result in invasion. However, this approach may not be sufficient to make the specific quantitative predictions necessary for management of invasive species. This is due to the simplifying assumption of constant genetic variance based on mutation-selection balance (Lande 1976). Complex simulations could relax this assumption and permit genetic variation to change via mutation, selection, and drift, in tandem with the demographic processes in a heterogeneous environment, and explicitly investigate the costs associated with avoiding Allee effects. Hence, future models should incorporate how propagule pressure (size and frequency of introduction events) impacts genetic variation and how more realistic genetic architectures contribute to the evolutionary trajectory of invasive species.
Although there is still much more work to be done to elucidate the factors that determine establishment success of founder populations, this theoretical approach has the promise to provide evidence in support of our working hypothesis that adaptive evolution can mitigate Allee effects and be an important driver of biological invasions.
Acknowledgments
We thank Michael Buhnerkempe, Greg Ames, Cameron Ghalambor, Ruth Hufbauer, and N. LeRoy Poff for discussions and advice on this research and this paper. We also thank two anonymous reviewers and the Associate Editor, Robert Holt, for helpful comments. The authors gratefully acknowledge the financial support for this work including the Global Invasions Network NSF-RCN DEB-#0541673; NSF-IGERT Grant DGE-#0221595, administered by the PRIMES program at Colorado State University for ARK; and NSF Grant DEB-0618097 for CTW.
Literature cited
- Allee WC. Animal Aggregations: A Study in General Sociology. Chicago: University of Chicago Press; 1931. [Google Scholar]
- Allendorf FW, Lundquist LL. Population biology, evolution, and control of invasive species. Conservation Biology. 2003;17:24–30. [Google Scholar]
- Berec L, Boukal DS, Berec M. Linking the Allee effect, sexual reproduction, and temperature-dependent sex determination via spatial dynamics. The American Naturalist. 2001;157:217–230. doi: 10.1086/318626. [DOI] [PubMed] [Google Scholar]
- Boukal DS, Berec L. Single-species models of the Allee effect: extinction boundaries, sex ratios and mate encounters. Journal of Theoretical Biology. 2002;210:375–394. doi: 10.1006/jtbi.2002.3084. [DOI] [PubMed] [Google Scholar]
- Briggs WH, Goldman IL. Genetic variation and selection response in model breeding populations of Brassica rapa following a diversity bottleneck. Genetics. 2006;172:457–465. doi: 10.1534/genetics.105.040899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bryant EH, McCommas SA, Combs LM. The effect of an experimental bottleneck upon quantitative genetic variation in the housefly. Genetics. 1986;114:1191–1211. doi: 10.1093/genetics/114.4.1191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carroll SP, Hendry AP, Reznick DN, Fox CW. Evolution on ecological time-scales. Functional Ecology. 2007;21:387–393. [Google Scholar]
- Cheverud JM, Routman EJ. Epistasis as a source of increased additive genetic variance at population bottlenecks. Evolution. 1996;50:1042–1051. doi: 10.1111/j.1558-5646.1996.tb02345.x. [DOI] [PubMed] [Google Scholar]
- Colautti RI, Grigorovich IA, MacIsaac HJ. Propagule pressure: a null model for biological invasions. Biological Invasions. 2006;8:1023–1037. [Google Scholar]
- Courchamp F, Clutton-Brock T, Grenfell B. Inverse density dependence and the Allee effect. Trends in Ecology and Evolution. 1999;14:405–410. doi: 10.1016/s0169-5347(99)01683-3. [DOI] [PubMed] [Google Scholar]
- Courchamp F, Berec L, Gascoigne J. Allee Effects in Ecology and Conservation. New York: Oxford University Press; 2008. [Google Scholar]
- Drake JM, Lodge DM, Lewis M. Theory and preliminary analysis of species invasions from ballast water: controlling discharge volume and location. American Midland Naturalist. 2005;154:459–470. [Google Scholar]
- Drury KLS, Drake JM, Lodge DM, Dwyer G. Immigration events dispersed in space and time: factors affecting invasion success. Ecological Modelling. 2007;206:63–78. [Google Scholar]
- Elton CS. The Ecology of Invasions by Animals and Plants. London, UK: Methuen; 1958. [Google Scholar]
- Falconer DS. Introduction to Quantitative Genetics. 3rd edn. New York: Wiley; 1989. [Google Scholar]
- Filin I, Holt RD, Barfield M. The relation of density regulation to habitat specialization, evolution of a species’ range, and the dynamics of biological invasions. The American Naturalist. 2008;172:233–247. doi: 10.1086/589459. [DOI] [PubMed] [Google Scholar]
- Fisher RA. The wave of advance of advantageous genes. Annals of Eugenics. 1937;7:355–369. [Google Scholar]
- Fitzhugh R. Thresholds and plateaus in the Hodgkin-Huxley nerve equations. Journal of General Physiology. 1960;43:867–896. doi: 10.1085/jgp.43.5.867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- García-Ramos G, Kirkpatrick M. Genetic models of adaptation and gene flow in peripheral populations. Evolution. 1997;51:21–28. doi: 10.1111/j.1558-5646.1997.tb02384.x. [DOI] [PubMed] [Google Scholar]
- Garvie MR. Finite difference schemes for reaction-diffusion equations modeling predator-prey interactions in MATLAB. Bulletin of Mathematical Biology. 2007;69:931–956. doi: 10.1007/s11538-006-9062-3. [DOI] [PubMed] [Google Scholar]
- Gascoigne J, Berec L, Gregory S, Courchamp F. Dangerously few liaisons: a review of mate-finding Allee effects. Population Ecology. 2009;51:355–372. [Google Scholar]
- Gomulkiewicz R, Holt RD. When does evolution by natural selection prevent extinction? Evolution. 1995;49:201–207. doi: 10.1111/j.1558-5646.1995.tb05971.x. [DOI] [PubMed] [Google Scholar]
- Goodnight CJ. Epistasis and the effect of founder events on the additive genetic variance. Evolution. 1988;42:441–454. doi: 10.1111/j.1558-5646.1988.tb04151.x. [DOI] [PubMed] [Google Scholar]
- Hare MP, Guenther C, Fagan WF. Nonrandom larval dispersal can steepen marine clines. Evolution. 2005;59:2509–2517. [PubMed] [Google Scholar]
- Holt RD, Gomulkiewicz R, Barfield M. The phenomenology of niche evolution via quantitative traits in a ‘black-hole’ sink. Proceedings of the Royal Society of London, B. 2003;270:215–224. doi: 10.1098/rspb.2002.2219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holt RD, Knight TM, Barfield M. Allee effects, immigration, and the evolution of species’ niches. The American Naturalist. 2004;163:253–262. doi: 10.1086/381408. [DOI] [PubMed] [Google Scholar]
- Jonsson M, Kindvall O, Jonsell M, Nordlander G. Modelling mating success of saproxylic beetles in relation to search behavior, population density and substrate abundance. Animal Behaviour. 2003;65:1069–1076. [Google Scholar]
- Keitt TH, Lewis MA, Holt RD. Allee effects invasion pinning and species borders. The American Naturalist. 2001;157:203–216. doi: 10.1086/318633. [DOI] [PubMed] [Google Scholar]
- Kierstead H, Slobodkin LB. The size of water masses containing plankton bloom. Journal of Marine Research. 1953;12:141–147. [Google Scholar]
- Kinnison MT, Hairston NG., Jr Eco-evolutionary conservation biology: contemporary evolution and the dynamics of persistence. Functional Ecology. 2007;21:444–454. [Google Scholar]
- Kirkpatrick M, Barton NH. Evolution of a species’ range. American Naturalist. 1997;150:1–23. doi: 10.1086/286054. [DOI] [PubMed] [Google Scholar]
- Kondrashov AS, Khibnik AI. Ecogenetical models as fast-slow systems. In: Schnol EE, editor. Studies in Mathematical Biology. Pushchino: Russian Academy of Science; 1996. pp. 88–123. [Google Scholar]
- Lande R. Natural selection and random genetic drift in phenotypic evolution. Evolution. 1976;30:314–334. doi: 10.1111/j.1558-5646.1976.tb00911.x. [DOI] [PubMed] [Google Scholar]
- Lee CE. Evolutionary genetics of invasive species. Trends in Ecology and Evolution. 2002;17:386–391. [Google Scholar]
- Levitan DR. Density-dependent selection on gamete traits in three congeneric sea urchins. Ecology. 2002;83:464–479. [Google Scholar]
- Lewis MA, Kareiva P. Allee dynamics and the spread of invading organisms. Theoretical Population Biology. 1993;43:141–158. [Google Scholar]
- Lockwood JL, Cassey P, Blackburn T. The role of propagule pressure in explaining species invasions. Trends in Ecology and Evolution. 2005;20:223–228. doi: 10.1016/j.tree.2005.02.004. [DOI] [PubMed] [Google Scholar]
- Mayr E. The nature of colonization in birds. In: Baker HG, Stebbins GL, editors. The Genetics of Colonizing Species. New York: Academic Press; 1965. pp. 29–47. [Google Scholar]
- Murray JD. Mathematical Biology. 2nd edn. New York: Springer-Verlag; 1993. [Google Scholar]
- Nagumo J, Arimoto S, Yoshizawa S. An active pulse transmission line simulating nerve axon. Proceedings of the Institute of Radio Engineers. 1962;50:2061–2070. [Google Scholar]
- Nei M, Maruyama T, Chakraborty R. The bottleneck effect and genetic variability in populations. Evolution. 1975;29:1–10. doi: 10.1111/j.1558-5646.1975.tb00807.x. [DOI] [PubMed] [Google Scholar]
- Okubo A. Diffusion and Ecological Problems: Mathematical Models. New York: Springer-Verlag; 1980. [Google Scholar]
- Pease CM, Lande R, Bull JJ. A model of population growth, dispersal and evolution in a changing environment. Ecology. 1989;70:1657–1664. [Google Scholar]
- Reznick DN, Ghalambor CK. The population ecology of contemporary adaptations: what empirical studies reveal about the conditions that promote adaptive evolution. Genetica. 2001;112:183–198. [PubMed] [Google Scholar]
- Roman J, Darling JA. Paradox lost: genetic diversity and the success of aquatic invasions. Trends in Ecology and Evolution. 2007;22:454–464. doi: 10.1016/j.tree.2007.07.002. [DOI] [PubMed] [Google Scholar]
- Sakai AK, Allendorf FW, Holt JS, Lodge DM, Molofsky J, With KA, Baughman S, et al. The population biology of invasive species. Annual Review of Ecology and Systematics. 2001;32:305–332. [Google Scholar]
- Shigesada N, Kawasaki K. Biology Invasions: Theory and Practice. New York: Oxford University Press; 1997. [Google Scholar]
- Skellam J. Random dispersal in theoretical populations. Biometrika. 1951;38:196–218. [PubMed] [Google Scholar]
- Stephens PA, Sutherland WJ, Freckleton RP. What is the Allee effect? Oikos. 1999;87:185–190. [Google Scholar]
- Tobin PC, Whitmire SL, Johnson DM, Bjørnstad ON, Liebhold AM. Invasion speed is affected by geographical variation in the strength of Allee effects. Ecology Letters. 2007;10:36–43. doi: 10.1111/j.1461-0248.2006.00991.x. [DOI] [PubMed] [Google Scholar]
- Turelli M, Barton NH. Will population bottlenecks and multilocus epistasis increase additive genetic variance? Evolution. 2006;60:1763–1776. [PubMed] [Google Scholar]
- Verhulst PF. Notice sur la loi que la population poursuit dans son accroissement. Correspondance Mathématique et Physique. 1838;10:113–121. [Google Scholar]
- Webb C. A complete classification of Darwinian extinction in ecological interactions. The American Naturalist. 2003;161:181–205. doi: 10.1086/345858. [DOI] [PubMed] [Google Scholar]
- Willis JH, Orr HA. Increased heritable variation following population bottlenecks: the role of dominance. Evolution. 1993;47:949–957. doi: 10.1111/j.1558-5646.1993.tb01249.x. [DOI] [PubMed] [Google Scholar]