
Figure 1. Summarizing intrinsic differences between PV- and CCK-containing basket cells and their synapses.
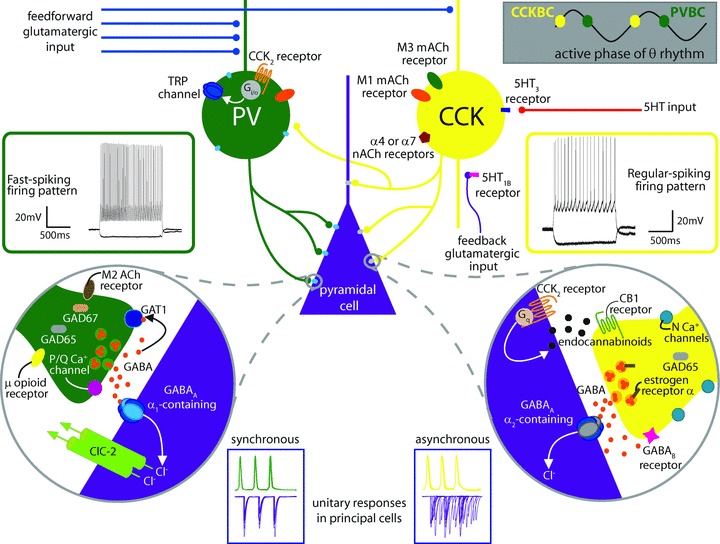
As described in the text, CCK BCs and PV BCs both target the perisomatic region of postsynaptic principal cells, but the distribution of CCK BC synapses is somewhat shifted toward the proximal dendrite, indicating a slight domain specificity. PV BCs (shown in green) have a fast spiking firing pattern (left, green inset) and are primarily activated by feedforward glutamatergic input (blue inputs) with high temporal fidelity while CCK BCs (shown in yellow) have regular spiking firing patterns (right, yellow inset) and because of their intrinsic proclivity for temporal summation are best activated by convergent feedforward and feedback (purple) glutamatergic input. PV BCs and CCK BCs are active at different phases of the theta cycle (upper right grey inset). CCK BCs express M1 (orange oval) and M3 (green oval) muscarinic acetylcholine (mACh) receptors as well as α7 or α4 nicotinic acetylcholine receptors (nACh, maroon pentagon) and 5HT3 receptors (dark blue rectangle). 5HT input (red input) can also affect 5HT1B receptors (pink rectangle) on CA1 pyramidal cell boutons to decrease feedforward excitation of CCK BCs. PV BCs express M1 receptors, extrasynaptic α1-containing GABAA receptors (light blue ovals) and CCK2 receptors (orange lines) coupled to a Gi/o pathway resulting in the opening of TRP channels (dark blue ovals) – non-selective cation channels that depolarize the cell (also see Fig. 2). At the same time, CCK2 receptors on pyramidal cells (orange lines, right circular inset) are coupled to Gq proteins which activate a molecular cascade leading to the synthesis of endocannabinoids (small black octagons). Endocannabinoids synthesized in response to this or other signals travel in a retrograde fashion to the CB1 receptors (green lines) on presynaptic CCK BC terminals to suppress release of GABA (small orange circles). As noted in the text, CCK BCs utilise N-type Ca2+ channels (light blue circles) located at a distance from the GABA release sites, express more GAD65 (grey rounded rectangle) than GAD 67, couple to mainly α2 subunit-containing GABAA receptors (grey oval), and express α oestrogen receptors (brown rectangle) and GABAB receptors (pink star) presynaptically. PV BC synapses (left circular inset) contain abundant GAD65 and GAD67 (tan rounded rectangle) and the GABA transporter GAT1 (blue rounded square), utilise P/Q-type Ca2+ channels (pink circle) located close to GABA release sites, and express μ opioid receptors (yellow oval) and M2 mACh receptors (brown oval), both of which inhibit release. PV BCs couple with mainly α1 subunit-containing GABAA receptors, and their inputs are modulated by the hyperpolarization-activated chloride channel, ClC-2 (green cylinders). Unitary responses in pyramidal cells (purple traces in blue insets, bottom) upon PV BC stimulation (green action potentials) are highly synchronous while responses to CCK BC stimulation (yellow action potentials) exhibit asynchronous release. Electrophysiological traces adapted by permission from Journal of Neuroscience; Lee SY, Földy C, Szabadics J & Soltesz I (2011). Cell-type-specific CCK2 receptor signalling underlies the cholecystokinin-mediated selective excitation of hippocampal parvalbumin-positive fast-spiking basket cells. J Neurosci31,10993–11002, copyright 2011.