

Abstract
Abelisaurids are a clade of large, bizarre predatory dinosaurs, most notable for their high, short skulls and extremely reduced forelimbs. They were common in Gondwana during the Cretaceous, but exceedingly rare in the Northern Hemisphere. The oldest definitive abelisaurids so far come from the late Early Cretaceous of South America and Africa, and the early evolutionary history of the clade is still poorly known. Here, we report a new abelisaurid from the Middle Jurassic of Patagonia, Eoabelisaurus mefi gen. et sp. nov., which predates the so far oldest known secure member of this lineage by more than 40 Myr. The almost complete skeleton reveals the earliest evolutionary stages of the distinctive features of abelisaurids, such as the modification of the forelimb, which started with a reduction of the distal elements. The find underlines the explosive radiation of theropod dinosaurs in the Middle Jurassic and indicates an unexpected diversity of ceratosaurs at that time. The apparent endemism of abelisauroids to southern Gondwana during Pangean times might be due to the presence of a large, central Gondwanan desert. This indicates that, apart from continent-scale geography, aspects such as regional geography and climate are important to reconstruct the biogeographical history of Mesozoic vertebrates.
Keywords: Dinosauria, Ceratosauria, Abelisauridae, Middle Jurassic, Argentina, Gondwanan biogeography
1. Introduction
Abelisaurids are a highly derived lineage of the Ceratosauria, a clade of phylogenetically basal theropod dinosaurs, characterized by a robust, short and high skull, extremely reduced forelimbs, and a highly pneumatized axial skeleton [1]. Abelisaurids were diverse in the Late Cretaceous of Gondwana, where they represent the top predators, but occurrences from the Northern Hemisphere are so far restricted to fragmentary and questionable latest Cretaceous remains from Europe [1]. Although the phylogenetic relationships of the more inclusive abelisauroids indicate that this clade reaches back to the Late Jurassic at least, the oldest secure occurrences of abelisauroids come from the late Early Cretaceous [1–4], and older reports of this clade are based on fragmentary and questionable material that does not provide much information on the early evolution of the group [1,5–7]. Thus, the origin and early history of abelisauroids remains largely unknown.
Here, we report a basal abelisaurid from the early Middle Jurassic of Patagonia, which extends the stratigraphic range of this clade for more than 40 Myr. The discovery of such an early member of this derived clade has important implications for the early evolution of abelisauroids and the origin of their bizarre morphology, as well as for establishing the initial diversification of ceratosaurs in general during Pangean times, which has important biogeographical consequences for understanding the distribution of theropods during the Jurassic.
2. Systematic palaeontology
Dinosauria Owen, 1842
Theropoda Marsh, 1881
Ceratosauria Marsh, 1884
Abelisauridae Bonaparte and Novas, 1985
Eoabelisaurus mefi, gen. et sp. nov.
(a). Etymology
From eos, Greek for dawn, for the early occurrence of the new taxon, and Abelisaurus, type genus of the Abelisauridae. The species name refers to the popular abbreviation of the Museo Paleontológico Egidio Feruglio (MEF).
(b). Holotype
Museo Paleontológico Egidio Feruglio (MPEF) PV 3990, an almost complete skeleton, including the posterior half of the skull, five cervical and nine dorsal vertebrae, complete sacrum, 27 caudal vertebrae, left and right scapulocoracoid, left and right humerus, radius and ulna, left and right distal carpal, left and right metacarpals I–IV, five phalanges (including one ungual) of the left and six phalanges (including one ungual) of the right hand, complete pelvic girdle, left and right femur, tibia, fibula, tarsals and metatarsals, and 12 phalanges (including two uguals) of the left and eight phalanges (including one ungual) of the right pes (figure 1).
Figure 1.

Middle Jurassic abelisaurid Eoabelisaurus mefi. (a) Skeletal reconstruction, showing preserved parts. Skull in left lateral (b) and posterior (c) views. Middle cervical vertebra in left lateral (d) and dorsal (e) views. (f) Anterior mid-dorsal vertebra in left lateral view. (g) Posterior dorsal vertebra in left lateral view. (h) Anterior caudal vertebra in left lateral view. (i) Right ulna and radius in medial view. (j) Right manus in plantar view. (k) Pelvic girdle in left lateral view. (l) Left femur in lateral view. Pedal ungual in lateral (m) and ventral (n) views. Abbreviations: al, additional lamina; ap, ambiens process; at, antitrochanter; boc, basioccipital condyle; bt, basal tubera; d, depression; dc, distal carpal; di, diapophysis; epi, epipophysis; f, frontal; fm, foramen magnum; g, groove; il, ilium; ios, interorbital septum; is, ischium; j, jugal; l, lacrimal; lt, lesser trochanter; Mc, metacarpal; n, notch; ns, neural spine; of, obturator foramen; ol, olecranon process; pa, parietal; pap, paroccipital process; ph, phalanges; pl, pleurocoel; po, postorbital; poz, postzygapophysis; pp, parapophysis; prel, prezygoepipophyseal lamina; ps, parasphenoid; psf, prespinal fossa; pt, pterygoid; ptw, pterygoid wing of the quadrate; pu, pubis; q, quadrate; qj, quadratojugal; r, radius; soc, supraoccipital crest; sq, squamosal; ts, trochanteric shelf; u, ulna; vf, ventral fossa. Scale bars: (a) 500 mm; (b–l) 50 mm; (m,n) 10 mm.
(c). Type locality and horizon
Jugo Loco locality, close to the village of Cerro Cóndor, Chubut, Argentina. The locality is placed in a series of finely bedded mud-, marl- and limestones of the Cañadón Asfalto Formation, early Middle Jurassic (Aalenian-Bajocian; see the electronic supplementary material).
(d). Diagnosis
Medium-sized theropod dinosaur (6–6.5 m total length) with the following autapomorphies: quadrate with thickened medial distal articular end and subparallel articular condyles; mid-dorsal vertebrae with double, V-shaped lamina extending from the parapophysis to the prezygodiapophyseal lamina; ulna with hypertrophied olecranon process, accounting for more than 30 per cent of its length; pubic foramen elongate, more than twice as long as high; ambiens process of pubis developed as a large, anterolaterally directed, convex expansion.
3. Description
Whereas the posterior half of the skeleton was found in articulation, the anterior dorsal and cervical vertebrae and forelimbs had been partially disarticulated prior to burial, and the skull was slightly moved from the vertebral column. Furthermore, the skull and anterior presacrals were exposed at the time of discovery and have thus partially been destroyed by erosion.
Of the snout, only a fragment of the right maxilla has been recovered and shows that the interdental plates are fused, but not striated in Eoabelisaurus. The posterior part of the skull is high, with an oval orbit and an enlarged infratemporal fenestra (figure 1b,c), as in other ceratosaurs [8–10]. The lacrimal antorbital fossa is covered laterally by an expanded lamina. There is a slight step with an anterior fossa on the jugal process of the postorbital, but an expanded suborbital flange is missing. The skull roof is not notably thickened and no cranial ornamentation is present. The frontal is largely fused to the parietal, with only minor parts of the suture visible within the supratemporal fossae. The nuchal crest is raised, but less so than in advanced abelisaurids [8,10]. The occipital ridge is strongly developed and thickens dorsally into a small, horizontal table. A quadrate foramen is absent. As in other ceratosaurs [9,10], the interorbital septum is ossified.
Cervical vertebrae (figure 1d,e) are short and have two pneumatic foramina on either side of the centra. The anteriormost preserved cervicals are amphicoelous, whereas middle and posterior cervical vertebrae are flat anteriorly, but deeply concave posteriorly and have a low ventral keel. Vertebral centra of middle and posterior cervicals are furthermore notably broader than high. Cervical neural arches are strongly pneumatized and have prominent prezygoepipophyseal laminae and large prespinal fossae. Cervical neural spines are anteroposteriorly short and very low. The epipophyses are tabular and about as high as the neural spine. Only the first two dorsal vertebrae have pleurocoels, but the neural arches of all dorsals are extensively pneumatized. Mid-dorsal parapophyses are set on long lateral stalks and the transverse processes and prezygapophyses are connected by a web of bone. The sacrum comprises six vertebrae that form a dorsally arched structure, with only slight swellings marking the border between the fused vertebral centra. Anterior caudal vertebrae (figure 1h) have high, massive centra without any ventral groove. Neural spines are high and anteroposteriorly narrow, and transverse processes are long and strongly dorsolaterally and slightly posteriorly directed, with a marked posterior expansion at their distal end. Hyposphene–hypantrum articulations are present in the anterior caudals, but become indistinct at around caudal 10. The length of vertebral centra remains constant over the preserved portion of the tail, but middle and posterior caudals are considerably lower. Mid-caudals have anteroposteriorly short, vertical neural spines. Distal caudal prezygapophyses are short.
The scapula has a broad shaft without distal expansion. The coracoid is large and semioval. Both humeri are poorly preserved, but show remarkably primitive characters. The articular head is slightly expanded, but far from the globular shape seen in noasaurids [11] and abelisaurids [8,12]. The deltopectoral crest is moderately developed and the distal end shows two rounded condyles. Radius and ulna are short and the ulna has a large olecranon process (figure 1i). The manus is strongly foreshortened, retains four digits and has a stout, blocky metacarpal II (figure 1j). Metacarpal I is about half the length of metacarpal II and considerably more slender than the latter; it bears a stout phalanx that is slightly longer than the metacarpal. Metacarpals II and III have broad extensor grooves with raised proximal margins. The non-terminal manual phalanges are about as long as wide and lack any constriction between the articular ends, and manual unguals are reduced.
Ilium, pubis and ischium are fused without visible sutures (figure 1k). The ilium is low and elongate, with a gently arched dorsal margin. The supra-acetabular crest is continuous with the lateral brevis shelf and the brevis fossa expands posteriorly. The pubis has an elongate obturator foramen and a narrow pubic apron that is V-shaped in cross section, with the apex pointing anteriorly. The distal ends of the pubes lack any anteroposterior expansion. A broad, well-developed antitrochanter is present in the ischium at the suture with the ilium and the ischium has a moderately expanded distal boot. The puboischiadic plate is extensive, with broad incisions adjacent to the shafts of the pubis and ischium. The femur is stout, only slightly bowed, and shows an anteromedially directed head (figure 1l). The lesser trochanter is low, but broad, with a well-developed trochanteric shelf. The fourth trochanter is developed as a stout, high ridge. As in other ceratosaurs, the crista tibiofibularis is expanded and oriented obliquely in relation to the femoral shaft. The tibia has an elevated, rectangular cnemial crest, and the fibular crest is proximally placed and well-developed. The fibula has a deep, posteriorly facing medial depression proximally and the iliofibularis tubercle is developed as a stout, oblique ridge. Astragalus and calcaneum are fused with each other and the tibia, but not the fibula. The metatarsus is long and slender, with metatarsal III reaching almost 50 per cent of the femoral length. As in Ceratosaurus [13], the posterior end of the proximal articular surface of metatarsal III is expanded transversely. Pedal unguals have a broad, triangular depression ventrally (figure 1n), as in other abelisaurids [14].
4. Discussion
In order to assess the phylogenetic position of Eoabelisaurus, we added the codings for this taxon to a combined matrix of two recently published phylogenetic datasets ([1,15]; see the electronic supplementary material). The analysis places Eoabelisaurus as the most basal member of the Abelisauridae (figure 2). Abelisaurid synapomorphies include the laterally covered lacrimal antorbital fossa, broad cervical prespinal fossae, anteroposteriorly short anterior caudal neural spines, absence of a ventral groove in the anterior caudals, presence of rudimentary centrodiapophyseal laminae in the anterior mid-caudals, reduced distal ginglymus in the manual phalanges, and the presence of a flexor depression in the pedal unguals. Alternative phylogenetic placements of Eoabelisaurus are significantly suboptimal, except for a slightly more basal position (as a stem-abelisauroid; see the electronic supplementary material).
Figure 2.

Calibrated phylogeny of ceratosaurian theropods. Thick grey lines indicate major ghost lineages in noasaurids and abelisaurids, resulting from the phylogenetic relationships of Eoabelisaurus. Dashed line in Spinostropheus indicates stratigraphic uncertainty for this taxon [16]. For abelisaurids, outlines of continents indicate geographical distribution.
The so far oldest certain abelisaurids were represented by fragmentary remains from the late Early Cretaceous of South America [2] and Africa [3,4] and older records of abelisauroids in general are fragmentary and questionable ([1,17]; see also the electronic supplementary material). Eoabelisaurus extends the temporal range of this clade for more than 40 Myr into the early Middle Jurassic (figure 2). The occurrence of a derived ceratosaur at that time indicates a rapid diversification of ceratosaurs in the Early to early Middle Jurassic. Indeed, the abelisaurid affinities of the new taxon demonstrate that all major ceratosaurian lineages (those leading to the clades comprising Elaphrosaurus and its closest relatives (‘elaphrosaurs’), ceratosaurids, noasaurids, abelisaurids) were established by this time. The apparent rarity of ceratosaurs in the Jurassic does most probably not reflect true low taxonomic diversity, but is owed to the poor fossil record of Jurassic theropods in the Southern Hemisphere ([18]; see also the electronic supplementary material). Together with the wide array of basal tetanuran taxa known from the Middle Jurassic [19] and the growing recognition that most, if not all coelurosaurian lineages also reach back to this time [20–23], this underlines the rapid and apparently explosive radiation of theropod dinosaurs in the late Early to early Middle Jurassic. Whereas current research on the origin and success of dinosaurs mainly focuses on the Triassic–Jurassic boundary [24,25], these findings lend support to the hypothesis that the rise of dinosaurs was a two-step process, marked by extinctions of basal archosaurs at the Triassic–Jurassic boundary and of several basal dinosaur lineages in the Pliensbachian–Toarcian, followed by an explosive radiation of ‘modern’ clades [26,27]. New discoveries and more research into this topic are needed to establish the timing and patterns of early diversification of dinosaurs in the Jurassic.
So far, abelisaurid anatomy was almost exclusively known from Late Cretaceous taxa that are highly aberrant in their morphology, especially in their unusual skull structure [8,10] and extreme reduction of the forelimbs [8,28,29]. Eoabelisaurus reveals a previously unknown stage in the evolution of abelisaurids, having only some of the cranial modifications and a unique combination of plesiomorphic and apomorphic features in its forelimb. The humerus is plesiomorphic and unreduced, whereas the ulna–radius are shortened, but do not differ significantly from more basal ceratosaurs (e.g. Ceratosaurus). The manus of Eoabelisaurus, however, has a derived morphology, with short and robust metacarpals, non-terminal phalanges that are as broad as long and lack gynglimoidal distal articular surfaces, and reduced manual unguals. Consequently, the modification of the forelimb in the evolution of abelisauroids seems to have occurred in a modular fashion, starting at the distal elements, and only later affected the more proximal elements (figure 3). Eoabelisaurus thus confirms a recently proposed evolutionary scenario for the evolution of abelisaurid limbs based on the incomplete evidence of Cretaceous abelisauroids [29]. The phylogenetic position of the new taxon indicates that modifications similar to those of abelisaurids in the humerus of ‘elaphrosaurs’ [30,31] and noasaurids [11] either arose independently from those in derived abelisaurids, or that these modifications were reversed in Eoabelisaurus. Either way, this suggests a surprisingly high level of homoplasy in such an extreme specialization.
Figure 3.
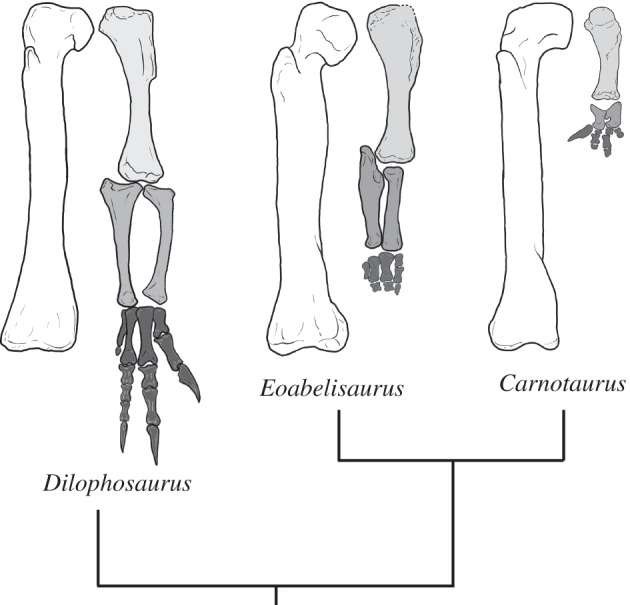
Limb proportions in several theropods, showing reduction of distal elements of the forelimb. Femur and forelimb of the basal theropod Dilophosaurus (left), basal abelisaurid Eoabelisaurus (middle) and the derived abelisaurid Carnotaurus (right). Drawn to same femoral length for comparison.
With a 40 Myr gap, the fossil record of abelisaurids is certainly still too poor to provide a robust biogeographic scenario [32]. Based on the previously available fossil record, abelisaurids were thought to have originated after the separation of Gondwana from the continents of the Northern Hemisphere [1,33], and recent discussions have focused on the possible effects (or lack thereof) of Gondwanan fragmentation on abelisaurid biogeography during the Cretaceous [1,3,4,33–35]. The discovery of Eoabelisaurus demonstrates that there was ample time for a dispersal of abelisaurids over the Gondwanan continents prior to their separation in the course of the ‘middle’ to Late Cretaceous, as it has been argued recently on the basis of geological palaeogeographic evidence [33].
However, the finding of an abelisaurid some 10–15 Myr before the effective separation of the continents of the Northern and Southern Hemispheres begs the question why no abelisauroid remains have been recorded from the Jurassic of Laurasia (a recent record of an abelisauroid from the Middle Jurassic of England [17] cannot be substantiated; see the electronic supplementary material). In contrast, other theropod lineages, which arose at approximately the same time, such as ‘elaphrosaurs’ [31], ceratosaurids [7,9,36] and basal tetanurans [19], achieved a Pangean distribution during this time (see the electronic supplementary material). Interestingly, a similar biogeographic pattern has recently been inferred for other groups of Mesozoic vertebrates, such as basal eusauropods [37], heterodontosaurid ornithischians [38] and mammals [39,40], in which some clades were restricted to southern Gondwana during the Jurassic, whereas other clades had a global distribution. The congruent pattern of restricted distribution for multiple vertebrate clades during Pangean times may be related to a common biogeographic cause. There is growing evidence from climate modelling and geological data for a large, central Gondwanan desert during the Middle and Late Jurassic [41–43], which might provide a possible explanation, acting as a filter for certain groups. However, further work on Jurassic palaeogeography and palaeoclimatology is needed. Nevertheless, this biogeographic pattern present in the above-mentioned clades of vertebrates during the Jurassic indicates that regional geographical and climatic conditions, and differential dispersal capabilities, may be as important to explain the biogeographic history of a group as continental fragmentation.
Acknowledgements
The authors thank the field crews of the January 2009 and February 2010 campaigns, during which the specimen was found and excavated. Mariano Caffa and Santiago Bessone did magic in the rapid preparation of the specimen. Norberto Pfeiffer is also thanked for additional preparation and curation. Jorge Gonzalez conducted the drawings and José María Farfaglia took photographs for the figures of electronic supplementary material. Matthew Carrano is thanked for critical comments on an earlier draft and providing most data on theropod distribution in the Paleobiology Database. The paper also benefitted from reviews by four anonymous reviewers. This project was supported by DFG grant RA 1012/9–1 and VW Foundation grant 84640 to O.W.M.R., FoncyT PICT 2006–1756 grant to D.P. and by the Fundación Egidio Feruglio. Fieldwork was made possible by the authorities of the province of Chubut and with logistic help from the MPEF.
References
- 1.Carrano M. T., Sampson S. D. 2008. The phylogeny of Ceratosauria (Dinosauria: Theropoda). J. Syst. Palaeontol. 6, 183–236 10.1017/S1477201907002246 (doi:10.1017/S1477201907002246) [DOI] [Google Scholar]
- 2.Rauhut O. W. M., Cladera G., Vickers-Rich P., Rich T. H. V. 2003. Dinosaur remains from the Lower Cretaceous of the Chubut Group, Argentina. Cret. Res. 24, 487–497 10.1016/S0195-6671(03)00067-3 (doi:10.1016/S0195-6671(03)00067-3) [DOI] [Google Scholar]
- 3.Sereno P. C., Brusatte S. L. 2008. Basal abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger. Acta Palaeont. Polonica 53, 15–46 10.4202/app.2008.0102 (doi:10.4202/app.2008.0102) [DOI] [Google Scholar]
- 4.Smith J. B., Lamanna M. C., Askar A. S., Bergig K. A., Tshakreen S. O., Abugares M. M., Rasmussen D. T. 2010. A large abelisauroid theropod dinosaur from the Early Cretaceous of Libya. J. Paleontol. 84, 927–934 10.1666/09-152.1 (doi:10.1666/09-152.1) [DOI] [Google Scholar]
- 5.Maganuco S., Cau A., Pasini G. 2005. First description of theropod remains from the Middle Jurassic (Bathonian) of Madagascar. Atti Soc. it. Sci. nat. Museo civ. Stor. nat. Milano. 146, 165–202 [Google Scholar]
- 6.Allain R., Tykoski R. S., Aquesbi N., Jalil N.-E., Monbaron M., Russell D. A., Taquet P. 2007. An abelisauroid (Dinosauria: Theropoda) from the Early Jurassic of the High Atlas Mountains, Morocco, and the radiation of ceratosaurs. J. Vert. Paleontol. 27, 610–624 10.1671/0272-4634(2007)27[610:AADTFT]2.0.CO;2 (doi:10.1671/0272-4634(2007)27[610:AADTFT]2.0.CO;2) [DOI] [Google Scholar]
- 7.Rauhut O. W. M. 2011. Theropod dinosaurs from the Late Jurassic of Tendaguru (Tanzania). Spec. Pap. Palaeontol. 86, 195–239 [Google Scholar]
- 8.Bonaparte J. F., Novas F. E., Coria R. A. 1990. Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia. Contrib. Sci. 416, 1–42 [Google Scholar]
- 9.Madsen J. H. J., Welles S. P. 2000. Ceratosaurus (Dinosauria, Theropoda). A revised osteology. Misc. Publ., Utah Geol. Surv. 00-2, 1–80 [Google Scholar]
- 10.Sampson S. D., Witmer L. M. 2007. Craniofacial anatomy of Majungsaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Soc. Vert. Paleontol., Mem. 8, 32–102 10.1671/0272-4634(2007)27[32:CAOMCT]2.0.CO;2 (doi:10.1671/0272-4634(2007)27[32:CAOMCT]2.0.CO;2) [DOI] [Google Scholar]
- 11.Carrano M. T., Loewen M. A., Sertich J. J. W. 2010. New materials of Masiakasaurus knopfleri Sampson, Carrano and Forster, 2001, and implications for the morphology of the Noasauridae. Smiths. Contrib. Paleobiol. 95, 1–53 [Google Scholar]
- 12.Carrano M. T. 2007. The appendicular skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Soc. Vert. Paleontol., Mem. 8, 163–179 10.1671/0272-4634(2007)27[163:TASOMC]2.0.CO;2 (doi:10.1671/0272-4634(2007)27[163:TASOMC]2.0.CO;2) [DOI] [Google Scholar]
- 13.Gilmore C. W. 1920. Osteology of the carnivorous Dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus. Bull. U.S. Natl. Mus. 110, 1–154 [Google Scholar]
- 14.Novas F. E., Dalla Vecchia F. M., Pais D. F. 2005. Theropod pedal unguals from the Late Cretaceous (Cenomanian) of Morocco, Africa. Rev. Mus. Argent. Cien. Nat. 7, 167–175 [Google Scholar]
- 15.Canale J. I., Scanferla C. A., Agnolin F. L., Novas F. E. 2009. New carnivorous dinosaur from the Late Cretaceous of NW Patagonia and the evolution of abelisaurid theropods. Naturwissenschaften 96, 409–414 10.1007/s00114-008-0487-4 (doi:10.1007/s00114-008-0487-4) [DOI] [PubMed] [Google Scholar]
- 16.Rauhut O. W. M., López-Arbarello A. 2009. Considerations on the age of the Tiouaren Formation (Iullemmeden Basin, Niger, Africa): implications for Gondwanan Mesozoic terrestrial vertebrate faunas. Palaeogeogr. Palaeoclim., Palaeoecol. 271, 259–267 10.1016/j.palaeo.2008.10.019 (doi:10.1016/j.palaeo.2008.10.019) [DOI] [Google Scholar]
- 17.Ezcurra M. D., Agnolín F. L. In press An abelisauroid dinosaur from the Middle Jurassic of Laurasia and its implications on theropod palaeobiogeography and evolution. Proc. Geol. Assoc . (published online) (doi:10.1016/j.pgeola.2011.12.003) [Google Scholar]
- 18.Rauhut O. W. M., López-Arbarello A. 2008. Archosaur evolution during the Jurassic: a southern perspective. Rev. Asoc. Geol. Argent. 63, 557–585 [Google Scholar]
- 19.Benson R. B. J. 2010. A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods. Zool. J. Linn. Soc. 158, 882–935 10.1111/j.1096-3642.2009.00569.x (doi:10.1111/j.1096-3642.2009.00569.x) [DOI] [Google Scholar]
- 20.Hu D., Hou L., Zhang L., Xu X. 2009. A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461, 640–643 10.1038/nature08322 (doi:10.1038/nature08322) [DOI] [PubMed] [Google Scholar]
- 21.Rauhut O. W. M., Milner A. C., Moore-Fay S. C. 2010. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zool. J. Linn. Soc. 158, 155–195 10.1111/j.1096-3642.2009.00591.x (doi:10.1111/j.1096-3642.2009.00591.x) [DOI] [Google Scholar]
- 22.Xu X., Ma Q., Hu D. 2010. Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins. Chin. Sci. Bull. 55, 3971–3977 10.1007/s11434-010-4150-z (doi:10.1007/s11434-010-4150-z) [DOI] [Google Scholar]
- 23.Liu Y.-Q., Kuang H.-W., Jiang X.-J., Peng N., Xu H., Sun H.-Y. 2012. Timing of the earliest known feathered dinosaurs and transitional pterosaurs older than the Jehol Biota. Palaeogeogr. Palaeoclim. Palaeoecol. 323–325, 1–12 10.1016/j.palaeo.2012.01.017 (doi:10.1016/j.palaeo.2012.01.017) [DOI] [Google Scholar]
- 24.Brusatte S. L., Benton M. J., Ruta M., Lloyd G. T. 2008. Superiority, competition, and opportunism in the evolutionary radiation of dinosaurs. Science 321, 1485–1488 10.1126/science.1161833 (doi:10.1126/science.1161833) [DOI] [PubMed] [Google Scholar]
- 25.Brusatte S. L., Nesbitt S. J., Irmis R. B., Butler R. J., Benton M. J., Norell M. A. 2010. The origin and early radiation of dinosaurs. Earth-Sci. Rev. 101, 68–100 10.1016/j.earscirev.2010.04.001 (doi:10.1016/j.earscirev.2010.04.001) [DOI] [Google Scholar]
- 26.Allain R., Aquesbi N. 2008. Tazoudasaurus naimi (Dinosauria, Sauropoda) from the late Early Jurassic of Morocco. Geodiversitas 30, 345–424 [Google Scholar]
- 27.Allain R., Läng É. 2009. Origine et évolution des saurischiens. C. R. Palevol. 8, 243–256 10.1016/j.crpv.2008.09.013 (doi:10.1016/j.crpv.2008.09.013) [DOI] [Google Scholar]
- 28.Coria R. A., Chiappe L. M., Dingus L. 2002. A new close relative of Carnotaurus sastrei Bonaparte 1985 (Theropoda: Abelisauridae) from the Late Cretaceous of Patagonia. J. Vert. Paleontol. 22, 460–465 10.1671/0272-4634(2002)022[0460:ANCROC]2.0.CO;2 (doi:10.1671/0272-4634(2002)022[0460:ANCROC]2.0.CO;2) [DOI] [Google Scholar]
- 29.Burch S. H., Carrano M. T. 2012. An articulated pectoral girdle and forelimb of the abelisaurid theropod Majungasaurus crenatissimus from the Late Cretaceous of Madagascar. J. Vert. Paleontol. 32, 1–16 10.1080/02724634.2012.622027 (doi:10.1080/02724634.2012.622027) [DOI] [Google Scholar]
- 30.Janensch W. 1925. Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas. Palaeontographica 7 (Suppl. I), 1–99 [Google Scholar]
- 31.Xu X., et al. 2009. A Jurassic ceratosaur from China helps to clarify avian digital homologies. Nature 459, 940–944 10.1038/nature08124 (doi:10.1038/nature08124) [DOI] [PubMed] [Google Scholar]
- 32.Turner A. H., Smith N. D., Callery J. A. 2009. Gauging the effects of sampling failure in biogeographic analysis. J. Biogeogr. 36, 612–625 10.1111/j.1365-2699.2008.02020.x (doi:10.1111/j.1365-2699.2008.02020.x) [DOI] [Google Scholar]
- 33.Ali J. R., Krause D. W. 2011. Late Cretaceous bioconnections between Indo-Madagascar and Antarctica: refutation of the Gunnerus Ridge causeway hypothesis. J. Biogeogr. 38, 1855–1872 10.1111/j.1365-2699.2011.02546.x (doi:10.1111/j.1365-2699.2011.02546.x) [DOI] [Google Scholar]
- 34.Sereno P. C., Wilson J. A., Conrad J. L. 2004. New dinosaurs link southern landmasses in the Mid-Cretaceous. Proc. R. Soc. Lond. B 271, 1325–1330 10.1098/rspb.2004.2692 (doi:10.1098/rspb.2004.2692) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mahler L. 2005. Record of Abelisauridae (Dinosauria: Theropoda) from the Cenomanian of Morocco. J. Vert. Paleontol. 25, 236–239 10.1671/0272-4634(2005)025[0236:ROADTF]2.0.CO;2 (doi:10.1671/0272-4634(2005)025[0236:ROADTF]2.0.CO;2) [DOI] [Google Scholar]
- 36.Soto M., Perea D. 2008. A ceratosaurid (Dinosauria, Theropoda) from the Late Jurassic–Early Cretaceous of Uruguay. J. Vert. Paleontol. 28, 439–444 10.1671/0272-4634(2008)28[439:ACDTFT]2.0.CO;2 (doi:10.1671/0272-4634(2008)28[439:ACDTFT]2.0.CO;2) [DOI] [Google Scholar]
- 37.Remes K., Ortega F., Fierro I., Joger U., Kosma R., Ferrer J. M. M., Paldes S. N. H. M., Ide O. A., Maga A. 2009. A new basal sauropod dinosaur from the Middle Jurassic of Niger and the early evolution of Sauropoda. PLoS ONE 4, 1–13 10.1371/journal.pone.0005361 (doi:10.1371/journal.pone.0005361) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Pol D., Rauhut O. W. M., Becerra M. 2011. A Middle Jurassic heterodontosaurid dinosaur from Patagonia and the evolution of heterodontosaurids. Naturwissenschaften 98, 369–379 10.1007/s00114-011-0780-5 (doi:10.1007/s00114-011-0780-5) [DOI] [PubMed] [Google Scholar]
- 39.Rauhut O. W. M., Martin T., Ortiz-Jaureguizar E., Puerta P. 2002. A Jurassic mammal from South America. Nature 416, 165–168 10.1038/416165a (doi:10.1038/416165a) [DOI] [PubMed] [Google Scholar]
- 40.Rougier G. W., Garrido A., Gaetano L., Puerta P., Corbitt C., Novacek M. J. 2007. First Jurassic triconodont from South America. Am. Mus. Novitates 3580, 1–20 10.1206/0003-0082(2007)3580[1:FJTFSA]2.0.CO;2 (doi:10.1206/0003-0082(2007)3580[1:FJTFSA]2.0.CO;2) [DOI] [Google Scholar]
- 41.Rees P. M., Ziegler A. M., Valdes P. J. 2000. Jurassic phytogeography and climates: new data and model comparisons. In Warm climates in Earth history (eds Huber B. T., Macleod K. G., Wing S. L.), pp. 297–318 Cambridge, UK: Cambridge University Press [Google Scholar]
- 42.Sellwood B. W., Valdes P. J. 2008. Jurassic climates. Proc. Geol. Assoc. 119, 5–17 10.1016/S0016-7878(59)80068-7 (doi:10.1016/S0016-7878(59)80068-7) [DOI] [Google Scholar]
- 43.Myers T. S., Tabor N. J., Jacobs L. L. 2011. Late Jurassic paleoclimate of Central Africa. Palaeogeogr., Palaeoclim., Palaeoecol. 311, 111–125 10.1016/j.palaeo.2011.08.013 (doi:10.1016/j.palaeo.2011.08.013) [DOI] [Google Scholar]