

Abstract
Object
Thymosin beta 4 (Tβ4) is a regenerative multifunctional peptide. This study will test the hypothesis that Tβ4 treatment initiated 6 hours post-injury reduces brain damage and improves functional recovery in rats after traumatic brain injury (TBI).
Methods
TBI was induced by controlled cortical impact over the left parietal cortex. Young adult male Wistar rats with TBI were randomly divided into the following groups: 1) Saline group (n=7); 2) Tβ4-6 mg/kg group (n=8), and 3) Tβ4-30 mg/kg group (n=8). Tβ4 or saline was administered intraperitoneally starting at 6 hours post-injury and then repeated daily at 24 and 48 hours. An additional group of sham animals underwent surgery without injury (n=6). Sensorimotor function and spatial learning were assessed using a modified neurological severity score and Morris water maze tests, respectively. Animals were sacrificed 35 days after injury and brain sections processed to assess lesion volume, hippocampal cell loss, cell proliferation and neurogenesis after Tβ4 treatment.
Results
Compared to saline, Tβ4 treatment initiated 6 hours post-injury significantly improved sensorimotor functional recovery and spatial learning, reduced cortical lesion volume and hippocampal cell loss, and enhanced cell proliferation and neurogenesis in the injured hippocampus. The high dose of Tβ4 showed better beneficial effects compared to the low-dose treatment.
Conclusions
Tβ4 treatment initiated 6 hours post-injury provides both neuroprotection and neurorestoration after TBI, indicating that Tβ4 has promising therapeutic potential in TBI patients. These data warrant further investigation of the optimal dose and therapeutic window of Tβ4 treatment for TBI and the associated underlying mechanisms.
Keywords: neuroprotection, neurogenesis, rat, thymosin beta 4, traumatic brain injury
Introduction
Traumatic brain injury (TBI) is the leading cause of death and disability in young people.1 Despite advances in preclinical research as well as improved clinical intensive care in recent years, no effective pharmacological therapy is available to promote functional recovery after TBI.38,45,57 TBI clinical trials targeting a single pathophysiological pathway have failed; a successful therapy may require targeting multiple injury pathways.38,45 There is a compelling need to develop effective therapies to improve functional outcome after TBI.
Thymosin beta 4 (Tβ4), a peptide of 43-amino acids, was first isolated from calf thymus tissue and subsequently found in all mammalian species studied.41 The major intracellular function of Tβ4 is to sequester G-actin monomers, which is necessary for cell motility and organogenesis.12 Tβ4 is also a regenerative multifunctional peptide, which inhibits inflammation and apoptosis,54 and promotes tissue repair in skin,48 cornea,18 and heart.5,56 Tβ4 is an essential paracrine factor of endothelial progenitor cells,23 which mediates cardioprotection.66 Tβ4 promotes angiogenesis after ischemic injury.61,63 The safety profile of Tβ4 has been established in animals and humans.58
Tβ4 exists in the nervous system and is involved in many cellular processes including mobility, axonal path-finding, neurite formation, proliferation and neuronal survival.50,80 Our previous study demonstrates that Tβ4 improves neurological functional recovery in rodents with experimental autoimmune encephalomyelitis and stroke.42,82 Our recent study shows that Tβ4 treatment initiated 24 hours post-TBI does not reduce cortical lesion volume but reduces hippocampal cell loss and promotes endogenous neurorestorative effects, which potentially contribute to improved functional recovery.77 However, the therapeutic efficacy of early Tβ4 treatment for TBI has not been investigated. Although human TBI is a complex and multifaceted disease, pharmacological compounds delivered in a clinically-relevant time window may improve the outcome of TBI patients.37 In the present study, we test the hypothesis that Tβ4 initiated 6 hours post-injury provides neuroprotection by reducing cortical lesion volume and hippocampal cell loss, promotes brain remodeling by enhancing neurogenesis, and improves functional recovery in rats subjected to moderate TBI induced by unilateral controlled cortical impact.
Materials and Methods
All experimental procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Henry Ford Health System. All functional tests were performed by investigators blinded to the treatment status.
TBI Model
A controlled cortical impact (CCI) model of TBI in the rat was utilized for the present study.14,36 Young adult male Wistar rats (336.6 ± 15.2 g) were anesthetized with 35 mg/100 g body weight chloral hydrate intraperitoneally. Rectal temperature was maintained at 37°C using a feedback-regulated water-heating pad. Rats were placed in a stereotactic frame. Two 10-mm-diameter craniotomies were performed adjacent to the central suture, midway between lambda and bregma. The contralateral craniotomy allowed for movement of cortical tissue laterally. The dura mater was kept intact over the cortex. Injury was delivered by impacting the left (ipsilateral) cortex with a pneumatic piston containing a 6-mm-diameter tip at the rate of 4 m/second and 2.5 mm of compression. Velocity was measured with a linear velocity displacement transducer.
Experimental Groups and Treatment
TBI was induced by CCI over the left parietal cortex. Young adult male Wistar rats with TBI were randomly divided into 3 groups: 1) Saline group (n = 7); 2) Tβ4-6 mg/kg group (n = 8); and 3) Tβ4-30 mg/kg group (n = 8). Sham rats underwent surgery without injury (n = 6). Our previous studies show that 6–8 animals per group provide sufficient power to reach significance after TBI induced by CCI.33,36,52,79 Tβ4 used in the present study is a synthetic copy of the naturally-occurring 43-amino acid peptide provided by RegeneRx (Bethesda, MD). Immediately before injection, Tβ4 in lyophilized powder form was weighed and dissolved in saline at a concentration of either 0.6 or 3 mg/ml. The animals received a dose of either 6 or 30 mg/kg with different injection volumes based on body weight. Tβ4 dissolved in saline (RegeneRx, Bethesda, MD) was administered intraperitoneally starting at 6 hours after injury and then repeated one time daily once at 24 and 48 hours. The low doseof Tβ4 (6mg/kg) was selected based on our previous studies.42,77,82 A higher dose group (30 mg/kg) was selected based on a recent Tβ4 clinical trial in healthy humans,58 with a dose translation from humans using the body surface area normalization method.53 Animals in the saline-treated group received an equal volume of saline and were treated using the same protocol as Tβ4. To detect proliferating cells, 5-bromo-2′-deoxyuridine(BrdU, 100 mg/kg; Sigma, St. Louis, MO) was injected intraperitoneally into rats once daily for 10 days, starting 1 day after TBI. Allrats were sacrificed at 35 days after TBI or surgery.
Morris Water Maze Test
To detect spatial learning impairments, a recent version of the Morris water maze (MWM) test was used10 and all animals were tested 5 days (from 31–35 days after TBI or surgery) before sacrifice. The swimming pool was located in a large room, where there were many clues external to the maze (pictures on the walls, lamps and a camera on the ceiling); these were visible from the pool and presumably used by the rats for spatial orientation. The position of the cues remained unchanged throughout the experiment. Data collection was automated by the HVS Image 2020 Plus Tracking System (US HVS Image, San Diego, CA). For data collection, the blue pool (1.8 m diameter) was subdivided into four equal quadrants formed by imaging lines. At the start of a trial, the rat was placed at one of four fixed starting points, randomly facing a wall (designated North, South, East and West) and allowed to swim for 90 seconds or until it found the platform. If the animal found the platform, it was allowed to remain on it for 10 seconds. If the animal failed to find the platform within 90 seconds, it was placed on the platform for 10 seconds. Throughout the test period the platform was located in the northeast (NE) quadrant 2 cm below water in a randomly changing position, including locations against the wall, toward the middle of the pool, or off-center but always within the target quadrant. If the animal was unable to find the platform within 90 seconds, the trial was terminated and a maximum score of 90 seconds was assigned. If the animal reached the platform within 90 seconds, the percentage of time traveled within the NE (correct) quadrant was calculated relative to the total amount of time spent swimming before reaching the platform and employed for statistical analysis. The advantage of this version of the water maze is that each trial takes on the key characteristics of a probe trial because the platform is not in a fixed location within the target quadrant.59
Modified Neurological Severity Score (mNSS) Test
Neurological functional measurement was performed using the mNSS score test.7 The test was carried out on all rats preinjury and on Days 1, 4, 7, 14, 21, 28, and 35 after TBI. The mNSS is a composite of the motor (i.e., muscle status, abnormal movement), sensory (i.e., visual, tactile and proprioceptive), and reflex tests and has been employed in previous studies.33 In this TBI model, injury in the left hemispheric cortex of rats causes sensory and motor functional deficiency with elevated scores on motor, sensory, and Beam Balance Tests in the early phase after injury (Day 1 after injury). Absent reflexes and abnormal movements can be measured in rats with severe injury. Slow recovery in asymmetry deficiency as reflected by Beam Balance Test results has been reported in unilateral brain injuries including TBI33 and ischemia.7 This test is suitable for evaluating long-term neurological function after unilateral brain injury.
Tissue Preparation and Measurement of Lesion Volume
At day 35 after TBI, rats were anesthetized i.p. with chloral hydrate, then perfused transcardially first with saline solution, followed by 4% paraformaldehyde in 0.1 M PBS (pH 7.4). Their brains were removed and post-fixed in 4% paraformaldehyde for 2 days at room temperature. The brain tissue was cut into 7 equally spaced 1-mm coronal blocks and processed for paraffin sectioning. A series of adjacent 6-μm-thick sections were cut from each block in the coronal plane and stained with hematoxylin and eosin (H&E). For lesion volume measurement, the 7 brain sections were traced by a microcomputer imaging device (MCID) (Imaging Research, St. Catharine’s, Ontario, Canada), as previously described.8 The indirect lesion area was calculated (the intact area of the ipsilateral hemisphere is subtracted from the area of the contralateral hemisphere),71 and the lesion volume presented as a volume percentage of the lesion compared with the contralateral hemisphere. H&E sections from Blocks E and F containing hippocampus were used to acquire images of the dentate gyrus and CA3 regions at X 20 magnification. To evaluate the cell loss after TBI, we counted the cell number per millimeter in the dentate gyrus and CA3 regions.
Immunohistochemistry
To examine the effect of Tβ4 on cell proliferation, coronal sections were histochemically stained with mouse anti-BrdU.32 Briefly, 6-μm paraffin-embedded coronal sections were deparaffinized and rehydrated. Antigen retrieval was performed by boiling sections in 10-mM citrate buffer (pH 6.0) for 10 minutes.8 After washing with PBS, sections were incubated with 0.3 % H2O2 in PBS for 10 minutes, blocked with 1% BSA containing 0.3 % Triton-X 100 for 1 hour at room temperature, and incubated with mouse anti-BrdU (1:200; Dako, Carpinteria, CA) at 4°C overnight. After washing, sections were incubated with biotinylated anti-mouse antibody (1:200; Vector Laboratories, Inc., Burlingame, CA) for 30 minutes at room temperature. After an additional washing, sections were incubated with an avidin-biotin-peroxidase system (ABC kit, Vector Laboratories, Inc.). Diaminobenzidine (Sigma, St. Louis, MO) was then used as a sensitive chromogen for light microscopy. Sections were counterstained with hematoxylin. BrdU+ cells in the dentate gyrus and the cortex of ipsilateral hemispheres were examined at X20 magnification and counted.
Immunofluorescent Staining
To identify newly generated neurons in the dentate gyrus, a double labeling for BrdU and NeuN was performed. After dehydration, tissue sections were boiled in 10 mM citric acid buffer (pH 6) for 10 minutes. After washing with PBS, sections were incubated in 2.4 N HCl at 37°C for 20 minutes. Sections were then incubated with 1% BSA containing 0.3% Triton-X-100 in PBS, followed by incubation with mouse anti-NeuN antibody (1:200; Chemicon, Temecula, CA) at 4°C overnight. FITC-conjugated anti-mouse antibody (1:400; Jackson ImmunoResearch, West Grove, PA) was added to sections at room temperature for 2 hours. Sections were then incubated with rat anti-BrdU antibody (1:200; Dako, Glostrup, Denmark) at 4°C overnight, followed by incubation with Cy3-conjugated anti-rat antibody (1:400; Jackson ImmunoResearch) at room temperature for 2 hours. Each of the steps was followed by 3 5-minutes rinses in PBS. Tissue sections were mounted with Vectashield mounting medium (Vector Laboratories). Images were collected with fluorescent microscopy and merged. NeuN/BrdU-colabeled cells in the dentate gyrus and the cortex were counted at a magnification of 40.82
Cell Counting and Quantitation
The methods employed to count cells have been utilized in our previous studies.6,31,84 We did not use stereological counting methods but we counted cells using the principles of stereology.84 This method permits a meaningful comparison of differences among groups. For quantitative measurements of BrdU+, NeuN+, and NeuN+/BrdU+cells, 5 slides from each brain were analyzed, with each slide containing 5 fields of view in the lesion boundary zone from the epicenter of the injury cavity (bregma −3.3 mm), 3 fields of view in the ipsilateral CA3 and 9 fields of view in the ipsilateral dentate gyrus in the same section. The fields were digitized under the light microscope (Nikon, Eclipse 80i, Melville, NY) at either X 200 or X 400 magnification using a CoolSNAP color camera (Photometrics, Tucson, AZ) interfaced with MetaMorph image analysis system (Molecular Devices, Downingtown, PA). The immunopositive cells were calculated and divided by the measured areas, and presented as numbers per mm2. Cell counts were performed by observers blinded to the individual treatment status of the animals. All counting was performed on a computer monitor to improve visualization and in one focal plane to avoid oversampling.83 To evaluate whether Tβ4 administered i.p. reduces neuronal damage after TBI, the number of cells was counted in the dentate gyrus and CA3 region. Although H&E staining is not neuron-specific, the morphological characteristics of neuronal cells in the dentate gyrus and CA3 region aid in counting them. Counts were averaged and normalized by measuring the linear distance (in mm) of the dentate gyrus and CA3 for each section. For cell proliferation, the total number of BrdU+ cells was counted in the lesion boundary zone and dentate gyrus. The cells with BrdU (brown stained) that clearly localized to the nucleus (hematoxylin stained) were counted as BrdU+ cells. For analysis of neurogenesis, additional sections used in the above studies were used to evaluate neurogenesis in the dentate gyrus by calculating the density of BrdU-labeled cells and BrdU/NeuN-colabeled cells. We mainly focused on the ipsilateral dentate gyrus and its subregions, including the subgranular zone, granular cell layer, and the molecular layer. The number of BrdU+ cells (red stained) and NeuN/BrdU-colabeled cells (yellow after merge) were counted in the dentate gyrus and the lesion boundary zone. The percentage of NeuN/BrdU-colabeled cells over the total number of BrdU+ cells in the corresponding regions (dentate gyrus or cortex) was estimated and used as a parameter to evaluate neurogenesis.75
Statistical Analyses
All data are presented as the mean ± standard deviation. Data were analyzed by analysis of variance (ANOVA) for repeated measurements of functional tests (spatial performance and sensorimotor function). For lesion volume, cell loss, cell proliferation and neurogenesis, a one-way ANOVA followed by post hoc Student-Newman-Keuls tests was used to compare the difference between the Tβ4-treated, saline-treated and sham groups. Correlation analysis of functional recovery with the lesion volume and cell counts was conducted by the Pearson linear correlation coefficient. Statistical significance was set at p < 0.05.
Results
Tβ4 Treatment Initiated 6 H Post-Injury Improves Spatial Learning
The modified version of MWM protocol was used to detect spatial learning deficits.10,44 The time spent in the correct quadrant (i.e., NE) by sham rats increased significantly from days 32–35 after surgery, as compared to time spent in the correct quadrant at day 31 (Fig. 1A, F5, 35 = 224.55; p < 0.001). In testing of spatial memory, a statistically significant between-group effect on the time spent in the correct quadrant was noted in the MWM test (at Day 32, F3, 27 = 4.42, p = 0.012; at Day 33, F3, 27 = 12.07, p = 0.004; at Day 34, F3, 27 = 64.09, p < 0.001; and at Day 35, F3, 27 = 20.88, p < 0.001). Relative to the saline group, post-hoc testing demonstrated significantly increased time spent in the correct quadrant in both Tβ4-6 mg/kg group and Tβ4-30 mg/kg group at Day 33 (p < 0.001), Day 34 (p < 0.001) and Day 35 (p < 0.001). The Tβ4-30 mg/kg group significantly differed in time spent in the correct quadrant from the Tβ4-6 mg/kg group at Day 33 (p < 0.001), Day 34 (p < 0.001) and Day 35 (p = 0.024).
Fig. 1.

The effect of Tβ4 on functional recovery after TBI. A: Effect of Tβ4 on spatial learning function 31–35 days after TBI. TBI significantly impairs spatial learning at days 32–35 compared to sham controls (p < 0.05). Tβ4 improves spatial learning performance (i.e., increased percentage of time spent in the correct quadrant of the Morris water maze) measured by a modified version of the Morris water maze test at days 33–35 compared with the saline group (p < 0.05). B: Effect of Tβ4 on sensorimotor function detected on the modified neurological severity scores (mNSS). Tβ4 treatment significantly lowers mNSS scores at days 4–35 compared to the saline group (p < 0.05). Data represent mean ± SD. *p < 0.05 vs saline group. #p < 0.05 vs Tβ4-6 mg/kg group. n (rats/group) = 6 (sham), 7 (saline), 8 (Tβ4-6 mg/kg), and 8 (Tβ4-30 mg/kg).
Tβ4 Treatment Initiated 6 H Post-Injury Reduces Neurological Severity Score
Figure 1B shows that there is no significant difference in the mNSS score between the saline- and Tβ4-treated groups at day 1 post TBI. However, compared to the saline-treated group, significantly improved scores were measured at Days 4–35 after TBI in the Tβ4-treated group (for Tβ4-6 mg/kg group at Days 4, 7, 14, 21, 28, and 35, with p values of < 0.001, = 0.013, < 0.001, < 0.001, = 0.024 and = 0.003, respectively; and Tβ4-30 mg/kg group at Days 4–35, all p values < 0.001). Compared to the Tβ4-6 mg/kg group, the Tβ4-30 mg/kg group showed significantly reduced mNSS scores at Days 7, 14, 21, 28 and Day 35 with p values of < 0.001, < 0.001,. < 0.001, < 0.001 and = 0.018, respectively.
Tβ4 Treatment Initiated 6 Hours Post-Injury Reduces Cortical Lesion Volume in TBI Rats
Rats were sacrificed at 35 days post TBI for histological measurements. In the evaluation of contusion volume (Fig. 2), a significant between-group difference was found (F2, 21 =27.95; p < 0.001). As expected, sham-injured animals did not develop contusions. Tβ4 treatment (6 hours post-injury) at 6 mg/kg and 30 mg/kg reduced cortical lesion volume compared to the saline-treated group (p < 0.001 for the 6 and 30 mg/kg groups). The Tβ4-30 mg/kg group exhibited a significantly reduced lesion volume compared to the Tβ4-6 mg/kg group (p = 0.013).
Fig. 2.
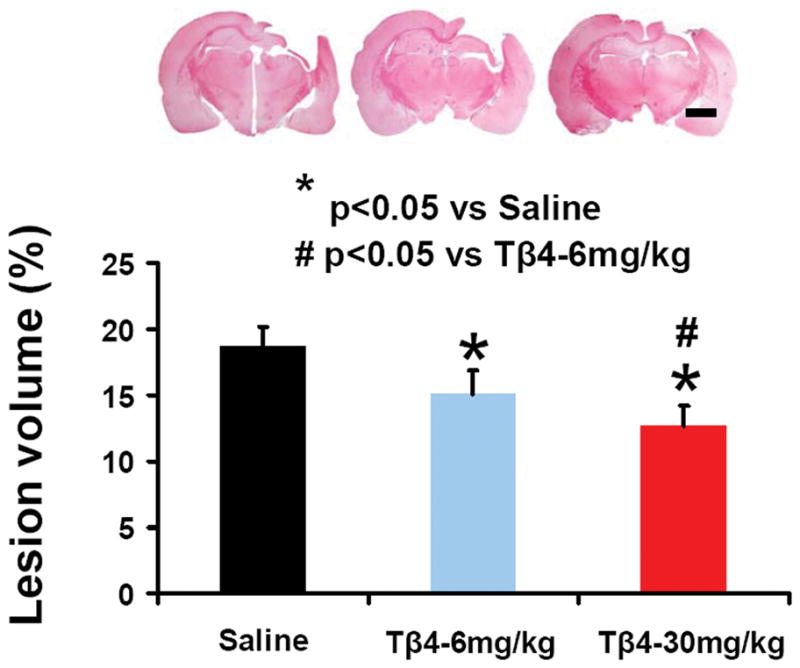
The effect of Tβ4 on cortical lesion volume examined 35 days after TBI. TBI caused significant cortical tissue loss. Tβ4 treatment significantly reduced lesion volume caused by TBI compared to the saline-treated groups (p < 0.05). Scale bar = 3 mm. Data represent mean ± SD. *p < 0.05 vs saline group. #p < 0.05 vs Tβ4-6 mg/kg group. n (rats/group) = 7 (saline), 8 (Tβ4-6 mg/kg), and 8 (Tβ4-30 mg/kg).
Tβ4 Treatment Initiated 6 Hours Post-Injury Reduces Hippocampal Cell Loss in TBI Rats
When examined at 35 days post TBI (Fig. 3), the analysis of selectively vulnerable hippocampal regions adjacent to the contusion revealed significant between-group effects in both the dentate gyrus (F3, 27 = 82.43, p < 0.001) and CA3 (F3, 27 = 26.91; p < 0.001) regions. The neuron counts in the ipsilateral CA3 (p < 0.001) and dentate gyrus (p < 0.001) had significantly decreased after TBI (Fig. 3B and 3) compared to sham controls (Fig. 3A and D). Tβ4 treatment (6-hours post-injury) at 6 mg/kg and 30 mg/kg significantly reduced hippocampal cell loss in the CA3 region and dentate gyrus compared to saline-treated group (p < 0.001 for the 6 and 30 mg/kg groups). A significantly reduced cell loss was observed in the Tβ4-30 mg/kg group in the dentate gyrus (p = 0.007) and CA3 (p = 0.005) regions compared to the Tβ4-6 mg/kg group.
Fig. 3.

The effect of Tβ4 on cell number in the ipsilateral dentate gyrus and CA3 region at 35 days after TBI. H&E staining: a–h. Treatment with Tβ4 (c, d, g, h) significantly reduces cell loss as compared with the saline-treated group (b, f) (p < 0.05). The cell number in the dentate gyrus and CA3 region is shown in (i). Scale bar = 50 μm (h, applicable to a–h). Data represent mean ± SD. *p < 0.05 vs saline group. #p < 0.05 vs Tβ4-6 mg/kg group. n (rats/group) = 6 (sham), 7 (saline), 8 (Tβ4-6 mg/kg), and 8 (Tβ4-30 mg/kg).
Tβ4 Treatment Increases Cell Proliferation in the Injured Cortex and Hippocampus
BrdU, an analog of thymidine, can be incorporated into the newly synthesized DNA of replicating cells during the S phase of the cell cycle, substituting for thymidine during DNA replication. BrdU staining is commonly used to detect proliferating cells. A statistically significant between-group effect on the number of BrdU-positive cells was noted in the dentate gyrus (F3, 27 =176.62, p < 0.001) and cortex (F3, 27 = 1018.75, p < 0.001). The post-hoc testing showed that the number of BrdU+ cells found in the ipsilateral cortex (p < 0.001) and dentate gyrus (p < 0.001) was significantly increased at 35 days after TBI, compared with sham controls (Fig. 4). However, Tβ4 treatment further increased the number of BrdU+ cells in the cortex (p < 0.001) and dentate gyrus (p < 0.011) after TBI compared to saline controls (Fig. 4). Significantly increased cell proliferation was noted in the Tβ4-30 mg/kg group compared to the Tβ4-6 mg/kg group (p< 0.001).
Fig. 4.

The effect of Tβ4 on cell proliferation in the injured cortex and dentate gyrus 35 days after TBI. TBI alone (b, f) significantly increases the number of BrdU+ cells in the ipsilateral cortex and dentate gyrus compared to sham controls (a, e) (p < 0.05). Tβ4 treatment significantly increases the number of BrdU+ cells in these regions (c, d, g, h; arrow as examples showing BrdU+ cell) compared to the saline-treated groups (p < 0.05). The number of BrdU+ cells is shown in (i). Scale bar = 25 μm (h, applicable to a–h). Data represent mean ± SD. *p < 0.05 vs saline group. #p < 0.05 vs Tβ4-6 mg/kg group. n (rats/group) = 6 (sham), 7 (saline), 8 (Tβ4-6 mg/kg), and 8 (Tβ4-30 mg/kg).
Tβ4 Treatment Increases Neurogenesis in the Injured Dentate Gyrus
To identify newly generated neurons, double immunofluorescent staining for BrdU (proliferating marker) and NeuN (mature neuronal marker) was performed. A statistically significant between-group effect on the number of newborn neurons was noted in the dentate gyrus (F3, 27 = 288.72, p < 0.001). The number of NeuN/BrdU-colabeled cells (newborn neurons) was significantly greater in the injured dentate gyrus (p < 0.001) after TBI compared to sham controls. Tβ4 treatment significantly increased the number of newborn neurons in the injured dentate gyrus (p < 0.001 for both doses of Tβ4) compared to saline controls (Fig. 5), while the Tβ4-30 mg/kg group exhibited a significantly increased number of newborn neurons compared to the Tβ4-6 mg/kg group (p < 0.001). As compared to sham animals (as baseline), neurogenesis increased by 2.1-, 4.5-, and 5.6-fold for saline, Tβ4-6 mg/kg, and Tβ4-30 mg/kg groups, respectively.
Fig. 5.

Double immunofluorescent staining for BrdU (red) and NeuN (green) to identify newborn neurons (yellow after merge) in the brain (a–d). The number of NeuN+ cells colabeled with BrdU is shown in (e). Scale bar = 25 μm (d, applicable to a–d). Data represent mean ± SD. *p < 0.05 vs saline group. #p < 0.05 vs Tβ4-6 mg/kg group. n (rats/group) = 6 (sham), 7 (saline), 8 (Tβ4-6 mg/kg), and 8 (Tβ4-30/kg).
Correlation of Functional Outcome with Lesion Volume, Cell Number, and Neurogenesis
To analyze the potential mechanism underlying the difference in functional outcome after Tβ4 treatment, we quantified the correlation of sensorimotor function with cortical lesion volume (Fig. 6A), spatial learning with hippocampal cell counts (Fig. 6B) and with dentate gyrus neurogenesis (Fig. 6C) examined at Day 35 after injury. The sensorimotor functional deficits are highly and significantly correlated with the cortical lesion volume (R2 = 0.7781, p < 0.001). The spatial learning is highly and significantly correlated with the cell number in the dentate gyrus (R2 = 0.7287, p < 0.001) and CA3 region (R2 = 0.7285, p < 0.001). In addition, the improved spatial learning is highly correlated with the number of newborn neurons in the dentate gyrus (R2 = 0.8104, p < 0.001). These data suggest that the Tβ4-improved functional recovery is due in part to its neuroprotection (reduced cortical lesion and hippocampal cell loss) and neurogenesis (increased newborn neurons) in the dentate gyrus.
Fig. 6.

Correlation of functional recovery with lesion volume, cell loss, and cell proliferation and neurogenesis examined 35 days after TBI. A: Significant correlation of mNSS with lesion volume. p < 0.001. B: Significant correlation of spatial learning with cell loss in the DG and CA3 region. p < 0.001. C: Significant correlation of spatial learning with BrdU+ cells in the cortex. D: Significant correlation of mNSS with BrdU+ cells in the DG. p < 0.001. E: Significant correlation of spatial learning with newborn neurons in the DG. n (rats/group) = 6 (sham), 7 (saline), 8 (Tβ4-6 mg/kg), and 8 (Tβ4-30 mg/kg).
Discussion
The present study provides the first evidence that Tβ4 treatment initiated 6 hours post injury provides a neuroprotective effect on reducing cortical lesion volume in rats after TBI. Tβ4 treatment significantly improves long-term behavioral benefits, including spatial learning and sensorimotor functional recovery, as evaluated by MWM and mNSS tests. The improvements in sensorimotor function are significantly correlated with the effect of Tβ4 treatment on reducing cortical lesion volume and increasing cortical cell proliferation. Improved spatial learning is significantly correlated with the effect of Tβ4 treatment on reducing hippocampal cell loss and increasing DG cell proliferation and neurogenesis. Our findings demonstrate that early Tβ4 treatment followed by repeated dosing not only provides neuroprotection but also promotes neurogenesis, indicating that Tβ4 is a potential therapy for TBI patients.
The findings in the present study differ from our previous study with delayed Tβ4 treatment for TBI. Our previous study demonstrates that Tβ4 treatment initiated 24 hours post-injury did not reduce cortical lesion volume but improved long-term functional recovery,77 which is consistent with our recent finding that Tβ4 treatment initiated 24 hours post-stroke improves functional recovery.42 The improved functional recovery may derive from the neurorestorative effects of delayed Tβ4 treatment including angiogenesis, oligodendrogenesis, and neurogenesis.43,77 Our present study shows that Tβ4 treatment initiated 6 hours post-injury significantly reduces cortical lesion volume and improves sensorimotor functional recovery (that is, reduced mNSS), clearly demonstrating that Tβ4 is neuroprotective and promising for treatment of TBI. Our current data are in line with the previous findings that neuroprotective agents significantly reduce brain damage including erythropoietin9 and cyclosporin A67 when administered within 6 hours after TBI. In addition, our present data show that Tβ4 treatment initiated 6 hours post-TBI reduces hippocampal cell loss. Similar neuroprotective effect (reduced hippocampal cell loss) was also seen with Tβ4 treatment initiated 24 hours post-injury.77 Our previous data demonstrate that apoptosis induced by CCI occurs as early as 2 hours after injury and persists up to 2 weeks afterwards.27 This finding supports the hypothesis that the early therapy combined with late repeated dosing for treatment of TBI may be required to provide maximum efficacy. The early treatment provides neuroprotection and the repeated delayed treatment maintains neuroprotection and promotes neurorestorative effects including neurogenesis as demonstrated in the present study and our previous study.77 Although we did not investigate apoptosis in the present study, Tβ4 provides the neuroprotection likely through its anti-apoptotic effect. Tβ4 reduces apoptosis in many cells including human corneal epithelial cells24 and gingival fibroblasts,54 rat cortical astrocytes81 and neurons.11 Intracerebroventricular administration of Tβ4 significantly reduces hippocampal neuronal loss induced by kainic acid.50 Several other Tβ4-derived peptides have been screened for biological activity.64 The first 15 amino acids of Tβ4 have been found to be major sites for cytoprotection and for antiapoptotic activity.64 In the present study, the rats were sacrificed at 35 days after TBI, which may not be suitable for evaluating the early antiapoptotic effect of Tβ4 treatment. Clearly, a more in-depth study evaluating the antiapoptotic effect of Tβ4 treatment with animals sacrificed at early and different time points after TBI is warranted. In addition, whether a single dose of Tβ4 treatment initiated 6 hours after TBI can afford a similar effect on neuroprotection and neurogenesis needs further investigation.
Neurogenesis occurs in the subventricular and subgranular zones of mammalian adult brains.20,85 TBI alone enhances neurogenesis in the dentate gyrus and subventricular zone.34,55,68,73,76 Newborn neurons in the dentate gyrus after TBI are capable of projecting axons into the CA3 region and may integrate in the existing neuronal circuitry.16,69 Injury-induced neurogenesis may partially contribute to spontaneous functional recovery after TBI. However, this endogenous neurogenesis is limited in the adult brain after injury. Previous studies from us and others demonstrate that increasing neurogenesis by therapeutic treatments may contribute to improved functional recovery after TBI.35,39,46,55,68,69,78 In the present study, although the causal link was not investigated, our correlation data indicate that the improvement in spatial learning is highly correlated with increased neurogenesis and cell counts at day 35 after TBI. Electrophysiological maturation of newborn neurons in the dentate gyrus progresses over a period of 2–7 weeks after cell division in rodents, which is accompanied by changes in expression of the immature neuron markers for the rst 3 weeks, with a subsequent transition to the mature neuronal marker NeuN by 4–6 weeks.29,86 In the present study, Tβ4 treatment further increases neurogenesis in the dentate gyrus after TBI compared to saline treatment. Neurogenesis plays a critical role in spatial learning and memory.28,35,39,69 In rodents, it is well established that the newborn neurons are involved in olfaction as well as in certain forms of memory and learning under normal and disease conditions.60 Eriksson et al. published the first pivotal study on hippocampal neural progenitor cells and neurogenesis in the human brain.17 Neurogenesis is involved in processes such as learning, memory, and depression, and may also be involved in regenerative attempts after injuries to the central nervous system while the contribution of neurogenesis to these phenomena remains to be elucidated, especially in humans.40 Collectively, our data suggest that Tβ4-induced increased neurogenesis of the dentate gyrus and reduced neuronal loss of the CA3 region and dentate gyrus may contribute to improvement in spatial learning after Tβ4 treatment.
Our present study indicates that the higher dose of Tβ4 (30 mg/kg) provides better functional and histological outcomes compared to the lower dose (6 mg/kg). A pharmacokinetic study of Tβ4 demonstrates that peak concentration is found in the brain at 40 minutes following intraperitoneal administration of Tβ4 in mice.41 In addition, the intravenous administration of Tβ4 over a dose range of 0.6–60 mg/kg is safe and tolerated without evidence of dose-limiting toxicity in all study animals.12 A recent clinical trial shows that intravenous administration of Tβ4 as a single dose or in multiple daily doses for 14 days over a dose range of 42–1260 mg is well tolerated with no evidence of dose-limiting toxicity in healthy humans.58 The mean half-life of Tβ4 in plasma increased with a single intravenous dose from 0.95 hours at 42 mg/subject, to 1.2 hours at 140 mg/subject, to 1.9 hours at 420 mg/subject, and to 2.1 hours at 1260 mg/subject.58 Whether the higher dose of Tβ4 (30 mg/kg) used in the present study is optimal remains unknown. This rat dose is equivalent to a human equivalent dose of 4.86 mg/kg, assuming 60 kg human, with a dose translation from rats using the body surface area normalization method.53 Our recent studies demonstrate that intraperitoneal injection of Tβ4 at 6 mg/kg starting at 24 hours after brain injury every 3 days improves neurological function in rat models of embolic stroke and TBI.42,77 To date, there is no effective pharmacological therapy available to promote functional recovery after TBI.30,45,57 Inflammation, oxidative injury, excitotoxicity, apoptosis, and hypoxia/ischemia contribute to secondary brain injury following TBI.72,74 Clinical TBI trials targeting a single pathophysiological pathway have failed; a successful therapy may require targeting multiple injury pathways.45,72 Tβ4 protects human corneal epithelial cells against oxidative injury,25 induces hair growth via stem cell migration and differentiation,49 promotes epicardium-derived neovascularization in the adult heart,56,62 reduces rat hippocampal neuronal loss by kainic acid excitotoxicity,50 reduces apoptotic death of chick and rat neurons,11 and inhibits inflammation.65 These studies provide convincing data that Tβ4 is a regenerative multifunctional peptide (it has anti-inflammatory, anti-oxidative, anti-apoptotic, pro-angiogenic, and tissue-protective effects), which could be a safe and promising agent for treatment of TBI.
Tβ4 is mainly recognized as a regulator of actin polymerization by its ability to form a complex with G-actin.64 This peptide has been implicated in angiogenesis, apoptosis, blood coagulation, carcinogenesis, lymphocyte maturation, and wound healing.26,70 Tβ4 is also involved in lesion-induced neuroplasticity through upregulation of microglia.15,47 The signaling pathways beyond actin sequestration remain largely unknown. Tβ4 lacks stable folded structure in aqueous solution and acquires specific folded structures only on binding to a partner protein; Tβ4 offers many possibilities for interaction with multiple partners.4,13,64 A candidate extracellular receptor of high affinity for Tβ4 is the β subunit of cell surface-located ATP synthase, which would allow extracellular Tβ4 to signal via a purinergic receptor to promote endothelial cell migration.19 Tβ4 induces endothelial progenitor cell migration via the PI3K/Akt/eNOS signal transduction pathway.51 Internalization of exogenous Tβ4 may be responsible for the antiapoptotic effects of exogenous Tβ4 on human corneal epithelial cells24 Internalization of the exogenous Tβ4 followed by activation of integrin-linked kinase and Akt seem to be the major mechanisms that promote the survival of cardiomyocytes.3,21 The Ku80 subunit of ATP-dependent DNA helicase II is associated with Tβ4 and functions as a novel receptor for Tβ4, mediating its intracellular activity.2 Ku80 has been implicated in multiple nuclear processes, e.g., DNA repair, telomere maintenance and apoptosis.22 Whether these signaling pathways mediate neuroprotection and neurorestoration of Tβ4 is unclear. Further investigation on these aspects is required.
In conclusion, the therapeutic efficacy of early Tβ4 treatment for TBI demonstrated in the present study suggests that Tβ4 is a potential therapy for TBI patients. Further investigation of the optimal dose and therapeutic window of Tβ4 treatment for TBI and the associated underlying mechanisms is warranted.
Acknowledgments
Sources of financial support: NINDS grants RO1 NS62002 (Ye Xiong) and PO1 NS023393 (Michael Chopp).
This work was supported by NIH Grant Nos. R01 NS62002 (to Dr. Xiong) and P01 NS023393 (to Dr. Chopp). Drs. ZG Zhang, Morris, and Chopp have applied for a patent for the use of Tβ4 as a treatment of stroke, neurological disease, and injury.
Footnotes
Disclosure
Author contributions to the study and manuscript preparation include the following. Conception and design: Xiong, Mahmood, ZG Zhang, Morris, Chopp. Acquisition of data: Xiong, Meng, Y Zhang. Analysis and interpretation of data: Xiong, Mahmood, ZG Zhang, Chopp. Drafting the article: Xiong, Chopp. Critically revising the article: Xiong, Chopp. Reviewed final version of the manuscript and approved it for submission: all authors.
References
- 1.Beauchamp K, Mutlak H, Smith WR, Shohami E, Stahel PF. Pharmacology of traumatic brain injury: where is the “golden bullet”? Mol Med. 2008;14:731–740. doi: 10.2119/2008-00050.Beauchamp. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bednarek R, Boncela J, Smolarczyk K, Cierniewska-Cieslak A, Wyroba E, Cierniewski CS. Ku80 as a novel receptor for thymosin beta4 that mediates its intracellular activity different from G-actin sequestering. J Biol Chem. 2008;283:1534–1544. doi: 10.1074/jbc.M707539200. [DOI] [PubMed] [Google Scholar]
- 3.Bock-Marquette I, Saxena A, White MD, Dimaio JM, Srivastava D. Thymosin beta4 activates integrin-linked kinase and promotes cardiac cell migration, survival and cardiac repair. Nature. 2004;432:466–472. doi: 10.1038/nature03000. [DOI] [PubMed] [Google Scholar]
- 4.Bubb MR. Thymosin beta 4 interactions. Vitam Horm. 2003;66:297–316. doi: 10.1016/s0083-6729(03)01008-2. [DOI] [PubMed] [Google Scholar]
- 5.Cavasin MA. Therapeutic potential of thymosin-beta4 and its derivative N-acetyl-seryl-aspartyl-lysyl-proline (Ac-SDKP) in cardiac healing after infarction. Am J Cardiovasc Drugs. 2006;6:305–311. doi: 10.2165/00129784-200606050-00003. [DOI] [PubMed] [Google Scholar]
- 6.Chen J, Li Y, Zhang R, Katakowski M, Gautam SC, Xu Y, et al. Combination therapy of stroke in rats with a nitric oxide donor and human bone marrow stromal cells enhances angiogenesis and neurogenesis. Brain Res. 2004;1005:21–28. doi: 10.1016/j.brainres.2003.11.080. [DOI] [PubMed] [Google Scholar]
- 7.Chen J, Sanberg PR, Li Y, Wang L, Lu M, Willing AE, et al. Intravenous administration of human umbilical cord blood reduces behavioral deficits after stroke in rats. Stroke. 2001;32:2682–2688. doi: 10.1161/hs1101.098367. [DOI] [PubMed] [Google Scholar]
- 8.Chen J, Zhang C, Jiang H, Li Y, Zhang L, Robin A, et al. Atorvastatin induction of VEGF and BDNF promotes brain plasticity after stroke in mice. J Cereb Blood Flow Metab. 2005;25:281–290. doi: 10.1038/sj.jcbfm.9600034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cherian L, Goodman JC, Robertson C. Neuroprotection with erythropoietin administration following controlled cortical impact injury in rats. J Pharmacol Exp Ther. 2007;322:789–794. doi: 10.1124/jpet.107.119628. [DOI] [PubMed] [Google Scholar]
- 10.Choi SH, Woodlee MT, Hong JJ, Schallert T. A simple modification of the water maze test to enhance daily detection of spatial memory in rats and mice. J Neurosci Methods. 2006;156:182–193. doi: 10.1016/j.jneumeth.2006.03.002. [DOI] [PubMed] [Google Scholar]
- 11.Choi SY, Noh MR, Kim DK, Sun W, Kim H. Neuroprotective function of thymosin-beta and its derivative peptides on the programmed cell death of chick and rat neurons. Biochem Biophys Res Commun. 2007;362:587–593. doi: 10.1016/j.bbrc.2007.08.031. [DOI] [PubMed] [Google Scholar]
- 12.Crockford D. Development of thymosin beta4 for treatment of patients with ischemic heart disease. Ann N Y Acad Sci. 2007;1112:385–395. doi: 10.1196/annals.1415.051. [DOI] [PubMed] [Google Scholar]
- 13.Crockford D, Turjman N, Allan C, Angel J. Thymosin beta4: structure, function, and biological properties supporting current and future clinical applications. Ann N Y Acad Sci. 2010;1194:179–189. doi: 10.1111/j.1749-6632.2010.05492.x. [DOI] [PubMed] [Google Scholar]
- 14.Dixon CE, Clifton GL, Lighthall JW, Yaghmai AA, Hayes RL. A controlled cortical impact model of traumatic brain injury in the rat. J Neurosci Methods. 1991;39:253–262. doi: 10.1016/0165-0270(91)90104-8. [DOI] [PubMed] [Google Scholar]
- 15.Dong JH, Ying GX, Liu X, Wang WY, Wang Y, Ni ZM, et al. Expression of thymosin beta4 mRNA by activated microglia in the denervated hippocampus. Neuroreport. 2005;16:1629–1633. doi: 10.1097/01.wnr.0000183326.21241.48. [DOI] [PubMed] [Google Scholar]
- 16.Emery DL, Fulp CT, Saatman KE, Schutz C, Neugebauer E, McIntosh TK. Newly born granule cells in the dentate gyrus rapidly extend axons into the hippocampal CA3 region following experimental brain injury. J Neurotrauma. 2005;22:978–988. doi: 10.1089/neu.2005.22.978. [DOI] [PubMed] [Google Scholar]
- 17.Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, et al. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4:1313–1317. doi: 10.1038/3305. [DOI] [PubMed] [Google Scholar]
- 18.Fine JD. Epidermolysis bullosa. a genetic disease of altered cell adhesion and wound healing, and the possible clinical utility of topically applied thymosin beta4. Ann N Y Acad Sci. 2007;1112:396–406. doi: 10.1196/annals.1415.017. [DOI] [PubMed] [Google Scholar]
- 19.Freeman KW, Bowman BR, Zetter BR. Regenerative protein thymosin beta-4 is a novel regulator of purinergic signaling. FASEB J. 2011;25:907–915. doi: 10.1096/fj.10-169417. [DOI] [PubMed] [Google Scholar]
- 20.Gage FH. Neurogenesis in the adult brain. J Neurosci. 2002;22:612–613. doi: 10.1523/JNEUROSCI.22-03-00612.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gnecchi M, He H, Noiseux N, Liang OD, Zhang L, Morello F, et al. Evidence supporting paracrine hypothesis for Akt-modified mesenchymal stem cell-mediated cardiac protection and functional improvement. Faseb J. 2006;20:661–669. doi: 10.1096/fj.05-5211com. [DOI] [PubMed] [Google Scholar]
- 22.Gullo C, Au M, Feng G, Teoh G. The biology of Ku and its potential oncogenic role in cancer. Biochim Biophys Acta. 2006;1765:223–234. doi: 10.1016/j.bbcan.2006.01.001. [DOI] [PubMed] [Google Scholar]
- 23.Hinkel R, El-Aouni C, Olson T, Horstkotte J, Mayer S, Muller S, et al. Thymosin beta4 is an essential paracrine factor of embryonic endothelial progenitor cell-mediated cardioprotection. Circulation. 2008;117:2232–2240. doi: 10.1161/CIRCULATIONAHA.107.758904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ho JH, Chuang CH, Ho CY, Shih YR, Lee OK, Su Y. Internalization is essential for the antiapoptotic effects of exogenous thymosin beta-4 on human corneal epithelial cells. Invest Ophthalmol Vis Sci. 2007;48:27–33. doi: 10.1167/iovs.06-0826. [DOI] [PubMed] [Google Scholar]
- 25.Ho JH, Tseng KC, Ma WH, Chen KH, Lee OK, Su Y. Thymosin beta-4 upregulates anti-oxidative enzymes and protects human cornea epithelial cells against oxidative damage. Br J Ophthalmol. 2008;92:992–997. doi: 10.1136/bjo.2007.136747. [DOI] [PubMed] [Google Scholar]
- 26.Huff T, Muller CS, Otto AM, Netzker R, Hannappel E. beta-Thymosins, small acidic peptides with multiple functions. Int J Biochem Cell Biol. 2001;33:205–220. doi: 10.1016/s1357-2725(00)00087-x. [DOI] [PubMed] [Google Scholar]
- 27.Kaya SS, Mahmood A, Li Y, Yavuz E, Goksel M, Chopp M. Apoptosis and expression of p53 response proteins and cyclin D1 after cortical impact in rat brain. Brain Res. 1999;818:23–33. doi: 10.1016/s0006-8993(98)01204-9. [DOI] [PubMed] [Google Scholar]
- 28.Kleindienst A, McGinn MJ, Harvey HB, Colello RJ, Hamm RJ, Bullock MR. Enhanced hippocampal neurogenesis by intraventricular S100B infusion is associated with improved cognitive recovery after traumatic brain injury. J Neurotrauma. 2005;22:645–655. doi: 10.1089/neu.2005.22.645. [DOI] [PubMed] [Google Scholar]
- 29.Kohler SJ, Williams NI, Stanton GB, Cameron JL, Greenough WT. Maturation time of new granule cells in the dentate gyrus of adult macaque monkeys exceeds six months. Proc Natl Acad Sci U S A. 2011;108:10326–10331. doi: 10.1073/pnas.1017099108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Loane DJ, Faden AI. Neuroprotection for traumatic brain injury. translational challenges and emerging therapeutic strategies. Trends Pharmacol Sci. 2010;31:596–604. doi: 10.1016/j.tips.2010.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lu D, Li Y, Wang L, Chen J, Mahmood A, Chopp M. Intraarterial administration of marrow stromal cells in a rat model of traumatic brain injury. J Neurotrauma. 2001;18:813–819. doi: 10.1089/089771501316919175. [DOI] [PubMed] [Google Scholar]
- 32.Lu D, Mahmood A, Qu C, Goussev A, Schallert T, Chopp M. Erythropoietin enhances neurogenesis and restores spatial memory in rats after traumatic brain injury. J Neurotrauma. 2005;22:1011–1017. doi: 10.1089/neu.2005.22.1011. [DOI] [PubMed] [Google Scholar]
- 33.Lu D, Mahmood A, Qu C, Hong X, Kaplan D, Chopp M. Collagen scaffolds populated with human marrow stromal cells reduce lesion volume and improve functional outcome after traumatic brain injury. Neurosurgery. 2007;61:596–602. doi: 10.1227/01.NEU.0000290908.38438.B2. discussion 602–593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lu D, Mahmood A, Zhang R, Copp M. Upregulation of neurogenesis and reduction in functional deficits following administration of DEtA/NONOate, a nitric oxide donor, after traumatic brain injury in rats. J Neurosurg. 2003;99:351–361. doi: 10.3171/jns.2003.99.2.0351. [DOI] [PubMed] [Google Scholar]
- 35.Lu D, Qu C, Goussev A, Jiang H, Lu C, Schallert T, et al. Statins increase neurogenesis in the dentate gyrus, reduce delayed neuronal death in the hippocampal CA3 region, and improve spatial learning in rat after traumatic brain injury. J Neurotrauma. 2007;24:1132–1146. doi: 10.1089/neu.2007.0288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mahmood A, Lu D, Chopp M. Marrow stromal cell transplantation after traumatic brain injury promotes cellular proliferation within the brain. Neurosurgery. 2004;55:1185–1193. doi: 10.1227/01.neu.0000141042.14476.3c. [DOI] [PubMed] [Google Scholar]
- 37.Marklund N, Bakshi A, Castelbuono DJ, Conte V, McIntosh TK. Evaluation of pharmacological treatment strategies in traumatic brain injury. Curr Pharm Des. 2006;12:1645–1680. doi: 10.2174/138161206776843340. [DOI] [PubMed] [Google Scholar]
- 38.Marklund N, Hillered L. Animal Modeling of Traumatic Brain Injury in Pre-clinical Drug Development - Where do we go from here? Br J Pharmacol. 2010 doi: 10.1111/j.1476-5381.2010.01163.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Meng Y, Xiong Y, Mahmood A, Zhang Y, Qu C, Chopp M. Dose-dependent neurorestorative effects of delayed treatment of traumatic brain injury with recombinant human erythropoietin in rats. J Neurosurg. 2011 doi: 10.3171/2011.3.JNS101721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ming GL, Song H. Adult neurogenesis in the mammalian brain: significant answers and significant questions. Neuron. 2011;70:687–702. doi: 10.1016/j.neuron.2011.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mora CA, Baumann CA, Paino JE, Goldstein AL, Badamchian M. Biodistribution of synthetic thymosin beta 4 in the serum, urine, and major organs of mice. Int J Immunopharmacol. 1997;19:1–8. doi: 10.1016/s0192-0561(97)00005-2. [DOI] [PubMed] [Google Scholar]
- 42.Morris DC, Chopp M, Zhang L, Lu M, Zhang ZG. Thymosin beta4 improves functional neurological outcome in a rat model of embolic stroke. Neuroscience. 2010;169:674–682. doi: 10.1016/j.neuroscience.2010.05.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Morris DC, Chopp M, Zhang L, Zhang ZG. Thymosin beta4: a candidate for treatment of stroke? Ann N Y Acad Sci. 2010;1194:112–117. doi: 10.1111/j.1749-6632.2010.05469.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Morris R. Developments of a water-maze procedure for studying spatial learning in the rat. J Neurosci Methods. 1984;11:47–60. doi: 10.1016/0165-0270(84)90007-4. [DOI] [PubMed] [Google Scholar]
- 45.Narayan RK, Michel ME, Ansell B, Baethmann A, Biegon A, Bracken MB, et al. Clinical trials in head injury. J Neurotrauma. 2002;19:503–557. doi: 10.1089/089771502753754037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ning R, Xiong Y, Mahmood A, Zhang Y, Meng Y, Qu C, et al. Erythropoietin promotes neurovascular remodeling and long-term functional recovery in rats following traumatic brain injury. Brain Res. 2011;1384:140–150. doi: 10.1016/j.brainres.2011.01.099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Paulussen M, Landuyt B, Schoofs L, Luyten W, Arckens L. Thymosin beta 4 mRNA and peptide expression in phagocytic cells of different mouse tissues. Peptides. 2009;30:1822–1832. doi: 10.1016/j.peptides.2009.07.010. [DOI] [PubMed] [Google Scholar]
- 48.Philp D, Goldstein AL, Kleinman HK. Thymosin beta4 promotes angiogenesis, wound healing, and hair follicle development. Mech Ageing Dev. 2004;125:113–115. doi: 10.1016/j.mad.2003.11.005. [DOI] [PubMed] [Google Scholar]
- 49.Philp D, St-Surin S, Cha HJ, Moon HS, Kleinman HK, Elkin M. Thymosin beta 4 induces hair growth via stem cell migration and differentiation. Ann N Y Acad Sci. 2007;1112:95–103. doi: 10.1196/annals.1415.009. [DOI] [PubMed] [Google Scholar]
- 50.Popoli P, Pepponi R, Martire A, Armida M, Pezzola A, Galluzzo M, et al. Neuroprotective effects of thymosin beta4 in experimental models of excitotoxicity. Ann N Y Acad Sci. 2007;1112:219–224. doi: 10.1196/annals.1415.033. [DOI] [PubMed] [Google Scholar]
- 51.Qiu FY, Song XX, Zheng H, Zhao YB, Fu GS. Thymosin beta4 induces endothelial progenitor cell migration via PI3K/Akt/eNOS signal transduction pathway. J Cardiovasc Pharmacol. 2009;53:209–214. doi: 10.1097/FJC.0b013e318199f326. [DOI] [PubMed] [Google Scholar]
- 52.Qu C, Mahmood A, Lu D, Goussev A, Xiong Y, Chopp M. Treatment of traumatic brain injury in mice with marrow stromal cells. Brain Res. 2008;1208:234–239. doi: 10.1016/j.brainres.2008.02.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Reagan-Shaw S, Nihal M, Ahmad N. Dose translation from animal to human studies revisited. FASEB J. 2008;22:659–661. doi: 10.1096/fj.07-9574LSF. [DOI] [PubMed] [Google Scholar]
- 54.Reti R, Kwon E, Qiu P, Wheater M, Sosne G. Thymosin beta4 is cytoprotective in human gingival fibroblasts. Eur J Oral Sci. 2008;116:424–430. doi: 10.1111/j.1600-0722.2008.00569.x. [DOI] [PubMed] [Google Scholar]
- 55.Richardson RM, Sun D, Bullock MR. Neurogenesis after traumatic brain injury. Neurosurg Clin N Am. 2007;18:169–181. xi. doi: 10.1016/j.nec.2006.10.007. [DOI] [PubMed] [Google Scholar]
- 56.Riley PR, Smart N. Thymosin beta4 induces epicardium-derived neovascularization in the adult heart. Biochem Soc Trans. 2009;37:1218–1220. doi: 10.1042/BST0371218. [DOI] [PubMed] [Google Scholar]
- 57.Royo NC, Schouten JW, Fulp CT, Shimizu S, Marklund N, Graham DI, et al. From cell death to neuronal regeneration: building a new brain after traumatic brain injury. J Neuropathol Exp Neurol. 2003;62:801–811. doi: 10.1093/jnen/62.8.801. [DOI] [PubMed] [Google Scholar]
- 58.Ruff D, Crockford D, Girardi G, Zhang Y. A randomized, placebo-controlled, single and multiple dose study of intravenous thymosin beta4 in healthy volunteers. Ann N Y Acad Sci. 2010;1194:223–229. doi: 10.1111/j.1749-6632.2010.05474.x. [DOI] [PubMed] [Google Scholar]
- 59.Schallert T. Behavioral tests for preclinical intervention assessment. NeuroRx. 2006;3:497–504. doi: 10.1016/j.nurx.2006.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Sierra A, Encinas JM, Maletic-Savatic M. Adult human neurogenesis: from microscopy to magnetic resonance imaging. Front Neurosci. 2011;5:47. doi: 10.3389/fnins.2011.00047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Smart N, Risebro CA, Melville AA, Moses K, Schwartz RJ, Chien KR, et al. Thymosin beta4 induces adult epicardial progenitor mobilization and neovascularization. Nature. 2007;445:177–182. doi: 10.1038/nature05383. [DOI] [PubMed] [Google Scholar]
- 62.Smart N, Risebro CA, Melville AA, Moses K, Schwartz RJ, Chien KR, et al. Thymosin beta-4 is essential for coronary vessel development and promotes neovascularization via adult epicardium. Ann N Y Acad Sci. 2007;1112:171–188. doi: 10.1196/annals.1415.000. [DOI] [PubMed] [Google Scholar]
- 63.Smart N, Rossdeutsch A, Riley PR. Thymosin beta4 and angiogenesis: modes of action and therapeutic potential. Angiogenesis. 2007;10:229–241. doi: 10.1007/s10456-007-9077-x. [DOI] [PubMed] [Google Scholar]
- 64.Sosne G, Qiu P, Goldstein AL, Wheater M. Biological activities of thymosin beta4 defined by active sites in short peptide sequences. FASEB J. 2010;24:2144–2151. doi: 10.1096/fj.09-142307. [DOI] [PubMed] [Google Scholar]
- 65.Sosne G, Qiu P, Kurpakus-Wheater M. Thymosin beta-4 and the eye: I can see clearly now the pain is gone. Ann N Y Acad Sci. 2007;1112:114–122. doi: 10.1196/annals.1415.004. [DOI] [PubMed] [Google Scholar]
- 66.Srivastava D, Saxena A, Michael Dimaio J, Bock-Marquette I. Thymosin beta4 is cardioprotective after myocardial infarction. Ann N Y Acad Sci. 2007;1112:161–170. doi: 10.1196/annals.1415.048. [DOI] [PubMed] [Google Scholar]
- 67.Sullivan PG, Sebastian AH, Hall ED. Therapeutic window analysis of the neuroprotective effects of cyclosporine a after traumatic brain injury. J Neurotrauma. 2011;28:311–318. doi: 10.1089/neu.2010.1646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Sun D, Bullock MR, McGinn MJ, Zhou Z, Altememi N, Hagood S, et al. Basic fibroblast growth factor-enhanced neurogenesis contributes to cognitive recovery in rats following traumatic brain injury. Exp Neurol. 2009;216:56–65. doi: 10.1016/j.expneurol.2008.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sun D, McGinn MJ, Zhou Z, Harvey HB, Bullock MR, Colello RJ. Anatomical integration of newly generated dentate granule neurons following traumatic brain injury in adult rats and its association to cognitive recovery. Exp Neurol. 2007;204:264–272. doi: 10.1016/j.expneurol.2006.11.005. [DOI] [PubMed] [Google Scholar]
- 70.Sun W, Kim H. Neurotrophic roles of the beta-thymosins in the development and regeneration of the nervous system. Ann N Y Acad Sci. 2007;1112:210–218. doi: 10.1196/annals.1415.013. [DOI] [PubMed] [Google Scholar]
- 71.Swanson RA, Morton MT, Tsao-Wu G, Savalos RA, Davidson C, Sharp FR. A semiautomated method for measuring brain infarct volume. J Cereb Blood Flow Metab. 1990;10:290–293. doi: 10.1038/jcbfm.1990.47. [DOI] [PubMed] [Google Scholar]
- 72.Vink R, Nimmo AJ. Multifunctional drugs for head injury. Neurotherapeutics. 2009;6:28–42. doi: 10.1016/j.nurt.2008.10.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Xiong Y, Lu D, Qu C, Goussev A, Schallert T, Mahmood A, et al. Effects of erythropoietin on reducing brain damage and improving functional outcome after traumatic brain injury in mice. J Neurosurg. 2008;109:510–521. doi: 10.3171/JNS/2008/109/9/0510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Xiong Y, Mahmood A, Chopp M. Emerging treatments for traumatic brain injury. Expert Opin Emerg Drugs. 2009;14:67–84. doi: 10.1517/14728210902769601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Xiong Y, Mahmood A, Lu D, Qu C, Kazmi H, Goussev A, et al. Histological and functional outcomes after traumatic brain injury in mice null for the erythropoietin receptor in the central nervous system. Brain Res. 2008;1230:247–257. doi: 10.1016/j.brainres.2008.06.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Xiong Y, Mahmood A, Meng Y, Zhang Y, Qu C, Schallert T, et al. Delayed administration of erythropoietin reducing hippocampal cell loss, enhancing angiogenesis and neurogenesis, and improving functional outcome following traumatic brain injury in rats: comparison of treatment with single and triple dose. J Neurosurg. 2009 doi: 10.3171/2009.9.JNS09844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Xiong Y, Mahmood A, Meng Y, Zhang Y, Zhang ZG, Morris DC, et al. Treatment of traumatic brain injury with thymosin beta in rats. J Neurosurg. 2011;114:102–115. doi: 10.3171/2010.4.JNS10118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Xiong Y, Mahmood A, Zhang Y, Meng Y, Zhang ZG, Qu C, et al. Effects of posttraumatic carbamylated erythropoietin therapy on reducing lesion volume and hippocampal cell loss, enhancing angiogenesis and neurogenesis, and improving functional outcome in rats following traumatic brain injury. J Neurosurg. 2011;114:549–559. doi: 10.3171/2010.10.JNS10925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Xiong Y, Qu C, Mahmood A, Liu Z, Ning R, Li Y, et al. Delayed transplantation of human marrow stromal cell-seeded scaffolds increases transcallosal neural fiber length, angiogenesis, and hippocampal neuronal survival and improves functional outcome after traumatic brain injury in rats. Brain Res. 2009;1263:183–191. doi: 10.1016/j.brainres.2009.01.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Yang H, Cheng X, Yao Q, Li J, Ju G. The promotive effects of thymosin beta4 on neuronal survival and neurite outgrowth by upregulating L1 expression. Neurochem Res. 2008;33:2269–2280. doi: 10.1007/s11064-008-9712-y. [DOI] [PubMed] [Google Scholar]
- 81.Yang H, Cui GB, Jiao XY, Wang J, Ju G, You SW. Thymosin-beta4 attenuates ethanol-induced neurotoxicity in cultured cerebral cortical astrocytes by inhibiting apoptosis. Cell Mol Neurobiol. 2010;30:149–160. doi: 10.1007/s10571-009-9439-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zhang J, Zhang ZG, Morris D, Li Y, Roberts C, Elias SB, et al. Neurological functional recovery after thymosin beta4 treatment in mice with experimental auto encephalomyelitis. Neuroscience. 2009;164:1887–1893. doi: 10.1016/j.neuroscience.2009.09.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zhang R, Wang Y, Zhang L, Zhang Z, Tsang W, Lu M, et al. Sildenafil (Viagra) induces neurogenesis and promotes functional recovery after stroke in rats. Stroke. 2002;33:2675–2680. doi: 10.1161/01.str.0000034399.95249.59. [DOI] [PubMed] [Google Scholar]
- 84.Zhang RL, Zhang ZG, Zhang L, Chopp M. Proliferation and differentiation of progenitor cells in the cortex and the subventricular zone in the adult rat after focal cerebral ischemia. Neuroscience. 2001;105:33–41. doi: 10.1016/s0306-4522(01)00117-8. [DOI] [PubMed] [Google Scholar]
- 85.Zhao C, Deng W, Gage FH. Mechanisms and functional implications of adult neurogenesis. Cell. 2008;132:645–660. doi: 10.1016/j.cell.2008.01.033. [DOI] [PubMed] [Google Scholar]
- 86.Zhao C, Teng EM, Summers RG, Jr, Ming GL, Gage FH. Distinct morphological stages of dentate granule neuron maturation in the adult mouse hippocampus. J Neurosci. 2006;26:3–11. doi: 10.1523/JNEUROSCI.3648-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]