

Abstract
Four different parts, hypocotyl and radicle (HR), inner cotyledon (IC), outer cotyledon (OC), seed coat and endosperm (SE), were sampled from mature rapeseed (Brassica napus L.) by laser microdissection. Subsequently, major secondary metabolites, glucosinolates and sinapine, as well as three minor ones, a cyclic spermidine conjugate and two flavonoids, representing different compound categories, were qualified and quantified in dissected samples by high-performance liquid chromatography with diode array detection and mass spectrometry. No qualitative and quantitative difference of glucosinolates and sinapine was detected in embryo tissues (HR, IC and OC). On the other hand, the three minor compounds were observed to be distributed unevenly in different rapeseed tissues. The hypothetic biological functions of the distribution patterns of different secondary metabolites in rapeseed are discussed.
Introduction
Seeds, the reproductive organs of plants, generally consist of seed coat, endosperm and embryo. Seed coats protect seeds during dormancy; endosperms normally provide nutrients during germination and, in the initial growth phase of the developing seedling; while embryos, which consist of cotyledons, hypocotyl and radicle, develop into different organs of the seedlings. According to the requirements of different physiological processes, nutrients and other metabolites are distributed and deposited in various seed organs. The embryo – which in the case of rapeseed (Brassica napus L.) refers especially to the cotyledons – is a storage site for lipids. In rapeseed, the oil contents reach approximately 50% (w/w) [1], making rape a major oil crop; worldwide it contributes up to 15% of global oil production [2]. Glucosinolates, which account for 3–8% of the rapeseed meal of conventional cultivars and 0.5–1.0% of low-glucosinolate cultivars, may have a depot function for nitrogen, as cyanogenic glucosides do [3]. Phenolic choline esters, mainly sinapate choline esters, are the other major class of secondary metabolites in rapeseed. Sinapine, the choline ester of sinapic acid (sinapate), is the predominant compound of that type, constituting 1–2% (w/w) of the rapeseed meal [4]. Although the sinapine biosynthesis pathway has been well investigated in Brassicaceae plants [5], the biological functions of sinapate choline esters are barely known. Sinapine was thought to be stored in Raphanus sativus seeds as a supply of choline, a compound that aids phosphatidylcholine biosynthesis in young seedlings [6]. From a nutritional point of view, the presence of the major secondary metabolites, glucosinolates and sinapates, are unwanted because of their antinutritive properties [1]. However, these compounds are very important for helping plants adapt to their biotic and abiotic environments [7], [8], and in plants different classes of secondary metabolites play specific ecological functions.
The glucosinolate-myrosinase system found in rape and other Brassicales is one of the best-explored plant chemical defense systems against herbivores [9]. Glucosinolate-derived indolics are also involved in antifungal defense [10]. Flavonoids, sinapates and other phenolics have been found in rapeseed and protect plants from ultraviolet-B (UV-B) stress [11]–[13]. Because different classes of secondary metabolites possess individual biological functions, it is reasonable to speculate that diverse secondary metabolites in rapeseed accumulate separately in specific tissues and play different roles in physiological processes or ecological interactions.
A recent study, in which laser microdissection (LMD) was successfully used to harvest specific tissues from developing rapeseed [14], encouraged us to apply LMD to sample different tissues of mature rapeseed and map the distribution of diverse secondary metabolites in the seed tissues. Insights gained from understanding how secondary metabolites are distributed in rapeseed can help us to conceive the biosynthesis and function of these metabolites in the plant.
LMD has been successfully used to harvest specific tissues or cells from plant material for transcript and protein analyses [15]–[17], and micro-spatial metabolic profiling studies [18]–[22]. In this study, LMD was used to sample four different parts, namely, hypocotyl and radicle (HR), inner cotyledon (IC), outer cotyledon (OC), seed coat and endosperm (SE) (Figure 1) from mature rapeseed. Secondary metabolites of different classes found in rapeseed cv. “Emerald,” namely glucosinolates, sinapine, a cyclic spermidine conjugate and flavonoids (unpublished data), were quantified in the extracts of dissected tissues by high-performance liquid chromatography - diode array detection and mass spectrometry (HPLC-DAD/MS). Here we report the distribution patterns of the above secondary metabolites in different rapeseed tissues and discuss their potential physiological and ecological relevance.
Figure 1. Work flow of laser microdissection of rapeseed.

(A) Progress of laser microdissection workflow applied to rapeseed. Hypocotyl and radicle (HR), inner cotyledon (IC), outer cotyledon (OC), seed coat and endosperm (SE) were successively dissected from rapeseed. (B) Micrographs of dissected tissues. Bar represents 1 mm.
Results and Discussion
Laser Microdissection of Rapeseed
The progress of LMD workflow applied to rapeseed is shown in Figure 1A. Four tissue parts, hypocotyl and radicle (HR), inner cotyledon (IC), outer cotyledon (OC), seed coat and endosperm (SE) (Figure 1B), were successively dissected from rapeseed cryosections and collected for analysis. HR, IC, and OC constitute the rapeseed embryo, and SE is material from the seed hull. The sampling was performed on four individual seeds. The weights of the four parts from each seed are listed in Table 1. The weights include the supporting polyethylene terephthalate (PET) membrane of the frame slide, which was unavoidably cut along with the seed tissues. The dissected materials were prepared for further analysis according to procedures described in the Materials and methods section.
Table 1. Weights (mg) of laser microdissected samples obtained from four individual seeds.
| Seed | HR | IC | OC | SE |
| 1 | 0.50 | 1.19 | 2.05 | 0.69 |
| 2 | 0.46 | 1.11 | 1.59 | 0.57 |
| 3 | 0.64 | 1.00 | 1.43 | 0.57 |
| 4 | 0.58 | 0.98 | 1.39 | 0.47 |
The samples include the supporting polyethylene terephthalate (PET) membrane of frame slides, which was cut together with the seed material. HR: hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; SE, seed coat and endosperm.
Glucosinolates in Rapeseed
Glucosinolates were determined in their desulfated form by HPLC-DAD/MS at 229 nm. Figure 2A shows chromatograms of the extracts of four seed tissues, HR, IC, OC and SE, dissected from rapeseed. Altogether, 11 desulfated glucosinolates, which have been recently identified in the “Emerald” cultivar of rapeseed (unpublished data), were determined by comparing MS data and retention times with those of references. The concentrations of identified glucosinolates (Figure 2B) from different seed tissues were calculated relative to the internal standard sinalbin. The concentration of glucosinolates in this cultivar is relatively high. Total glucosinolate concentrations in embryo tissues (HR, IC and OC) are higher than 100 µmol/g DW, and they are not statistically different between embryo tissues. Progoitrin (1) and gluconapin (6) are the predominant glucosinolates in this cultivar as they are in other rapeseed cultivars [23]. In the three embryo parts (HR, IC and OC), glucosinolate profiles are the same, and the individual glucosinolate concentrations are not significantly different. The concentrations of detected glucosinolates in SE samples are significantly lower than those in embryo tissues. Glucosinolates, glucoraphanin (3), gluconapoleiferin (4), glucoalyssin (5), glucoerucin (9), glucoberteroin (10) and gluconasturtiin (11) could not be detected in SE tissues, probably because of the very small amounts of dissected material available for analysis (Table 1), and the SE tissue is dominated by a hard seed coat.
Figure 2. Glucosinolate profiles and distribution in different rapeseed tissues.

(A) HPLC chromatograms of glucosinolate profiling in laser-microdissected samples from rapeseed detected at 229 nm. ▾ contamination peaks. (B) Total glucosinolate concentration and concentrations of individual glucosinolates 1–11 in four dissected samples. HR, hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; and SE, seed coat and endosperm. Each column shows the mean of four replicates with standard error. *means not detectable. Peaks: 1, progoitrin; 2, epiprogoitrin; 3, glucoraphanin; 4, gluconapoleiferin; 5, glucoalyssin; 6, gluconapin; 7, 4-hydroxyglucobrassicin; 8, glucobrassicanapin; 9, glucoerucin; 10, glucoberteroin; and 11, gluconasturtiin.
The even distribution of glucosinolates in mature rapeseed embryo tissues (HR, IC and OC) is consistent with the observation that myrosinase is expressed in all embyo tissues of developing rapeseed [24]. Glucosinolates of brassicaceous plants are well-known defense compounds, effective against herbivores and pathogens [25]–[27]. The evenly distributed glucosinolates in HR, IC and OC seem to provide protection for the entire embryo during seed dormancy. Glucosinolate levels decrease during germination of rapeseed [28] and Arabidopsis thaliana seeds [29], and the degradation products affect the interaction of plant roots with microorganisms [30]–[36], nematodes [37]–[40], other plants [41]–[43] and animals [39]. These evidences strongly indicate a depot function of glucosinolates in mature rapeseed as precursors of allelochemicals, which help the seedlings to establish the ecosystem in the rhizosphere.
Sinapine in Rapeseed
Sinapine, 12 (Figure 3A), the choline ester of sinapate, represents the dominant phenolic compound in rapeseed. The concentration of sinapine in four tested seeds of the “Emerald” cultivar averaged 20.36 µmol/g. Average sinapine concentrations (Figure 3B) found in three embryo tissues (HR, IC and OC) are close to each other, and all of them are higher than 22 µmol/g. The concentration detected in SE (0.72 µmol/g) is significantly lower than that in the embryo tissues. This finding is in accordance with the reported occurrence of sinapine mainly in rapeseed embryo [44].
Figure 3. Distribution of sinapine in rapeseed.
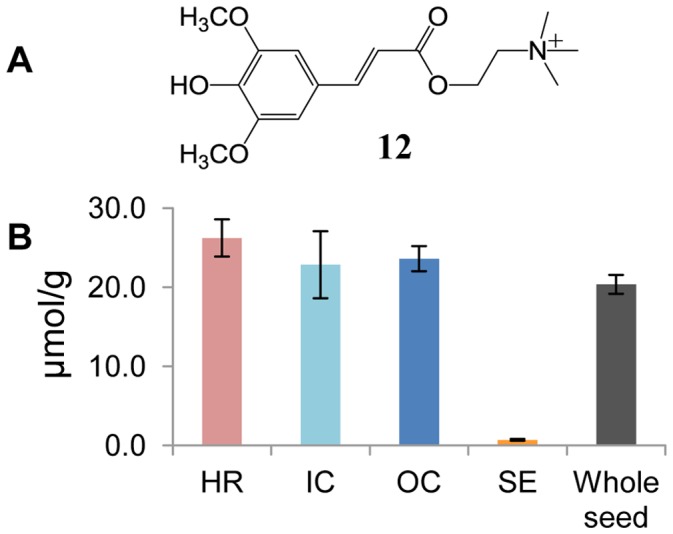
(A) Structure of sinapine (12). (B) Sinapine concentrations in different rapeseed tissues and whole rapeseed. HR, hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; and SE, seed coat and endosperm. Each column shows the mean of four replicates with standard error.
Sinapates, which are biosynthesized through the phenylpropanoid pathway, are chemotaxonomic markers of brassicaceous plants [45]. Sinapine is the major compound of that type in mature seeds. During early stages of seedling development sinapine is converted to sinapoylmalate via sinapate and sinapoylglucose [46], [47]. Sinapoylmalate protects plant leaves from UV-B irradiation [12], [48]–[51] and is involved in UV-B-induced defense against fungi in A. thaliana leaves [52]. On the other hand, much experimental evidence suggests that the sinapine stored in rapeseed provides a supply of sinapate and choline, both of which serve as important precursors for essential plant components. Sinapine (12) degrades into sinapate and choline during early stages of seed germination [6], [53], [54], and the two components are used in later biosynthetic processes [53]. In Raphanus sativus seedlings, choline released from sinapine was proven to be processed biosynthetically to phosphatidylcholine [6], and the sinapic acid moiety was hypothesized as the precursor for the biosynthesis of further phenolic compounds, such as flavonoids [53]. Thus, all products released or converted from sinapine during early steps of seed germination (sinapoylglucose, sinapoylmalate, sinapate and choline) play essential physiological and ecological roles for the seedling and plant [5]. The even distribution of sinapine in rapeseed embryo tissue supports its depot function.
Cyclic Spermidine Conjugates in Rapeseed
Cyclic spermidine conjugates in non-glucosinolate (NG) fractions of laser-microdissected rapeseed tissues were detected by HPLC-ESIMS in positive ionization mode (see Materials and methods). The major peak in extracted ion chromatogram (EIC) for ions at m/z 496.4 ([M+H]+) (Figure S1) was identified as the major cyclic spermidine conjugate (13) (Figure 4A), based on its molecular mass of 495 Da and comparing the retention time with the compound recently isolated from rapeseed (unpublished data). Based on the same molecular mass in the EIC and the same fragmentation patterns in MS/MS analysis compared to those of the major peak, several minor peaks (Figure S1) were suggested to be isomeric cyclic spermidine conjugates. However, structural details remained unassigned because nuclear magnetic resonance (NMR) data are lacking. The average concentration of compound 13 in the whole rapeseed is 1.94 µmol/g, as calculated from a calibration curve. Interestingly, the cyclic spermidine conjugates were found only in HR, where the average concentration of 13 is as high as 13.48 µmol/g. Compound 13 and minor cyclic spermidines are absent in SE, IC and OC tissues (Figures 4B, S1). No free spermidine was detected in any sample.
Figure 4. Distribution of the major cyclic spermidine in rapeseed.
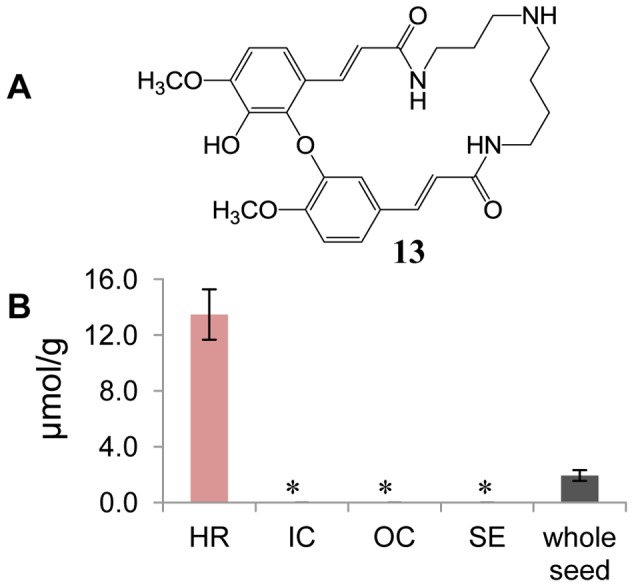
(A) Structure of the major cyclic spermidine conjugate (13) identified from rapeseed. (B) The concentration of 13 in different tissues and whole rapeseed. HR, hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; and SE, seed coat and endosperm. Each column shows the mean of four replicates with standard error, and *means not detectable.
Polyamines (PAs) and phenylpropanoid-polyamine conjugates (PPCs) are widely distributed in plants [55], including seeds [56], and play important roles in plant growth, abiotic stress tolerance and defense against insect herbivores [57]–[59]. Compound 13 (Figure 4A) was previously identified as the sole PPC from the same plant material, rapeseed [47], [60]. Nevertheless, this is the first time that the distribution of PPCs in seeds has been directly demonstrated. Our results showed that PPCs in rapeseed accumulate only in HR. This is consistent with the expression of PPC biosynthetic genes in Arabidopsis seeds [56]. The same authors also demonstrated that PPCs degrade at an early stage of seed germination [56]. Seeds of an Arabidopsis spermidine synthase-deficient double mutant contain a reduced level of spermidine and showed an abnormal phenotype [61]. The results indicated that spermidine, and probably other PAs as well, is essential for seed development in plants. Based on this evidence, PPCs that have accumulated in rapeseed are proposed to be sources of PAs and involved in diverse processes of plant growth and development [57], [58]. Although there is increasing interest on PAs functions in seed germination and seedling growth [62], [63], further experiments are needed to establish the precise roles of PPCs distributed in hypocotyl and/or radicle in rapeseed. Degradation products derived from PPCs also contain phenylpropanoids, which are universal precursors for condensed phenolics in plants.
Flavonoids in Rapeseed
Two major flavonoids, kaempferol-3-O-β-D-glucopyranosyl-(1→2)-β-D-glucopyranoside-7-O-β-D-glucopyranoside (14) and kaempferol-3-O-(2-O-sinapoyl)-β-D-glucopyranosyl-(1→2)-β-D-glucopyranoside-7-O-β-D-glucopyranoside (15) (Figure 5A), are known from the rape cultivar “Emerald” (unpublished data). Using calibration curves, the two flavonoids in dissected rapeseed samples were quantified by HPLC-ESIMS in negative mode. The average concentrations of flavonoids 14 and 15 in the whole seed are 0.23 and 0.42 µmol/g, respectively (Figure 5B). The distribution pattern of flavonoids in different rapeseed tissues is contrary to that of PPCs. Compounds 14 and 15 were mainly detected in cotyledon parts (IC and OC) (Figure S2), where their concentrations are similar. Meanwhile, the two flavonoids are not detectable in SE and almost undetectable in HR (Figure 5B). In fact, a trace of flavonoid 15 was detected in only one of the four HR samples. No kaempferol derivative was detectable in the other three HR samples.
Figure 5. Distribution of the two major flavonoids in rapeseed.
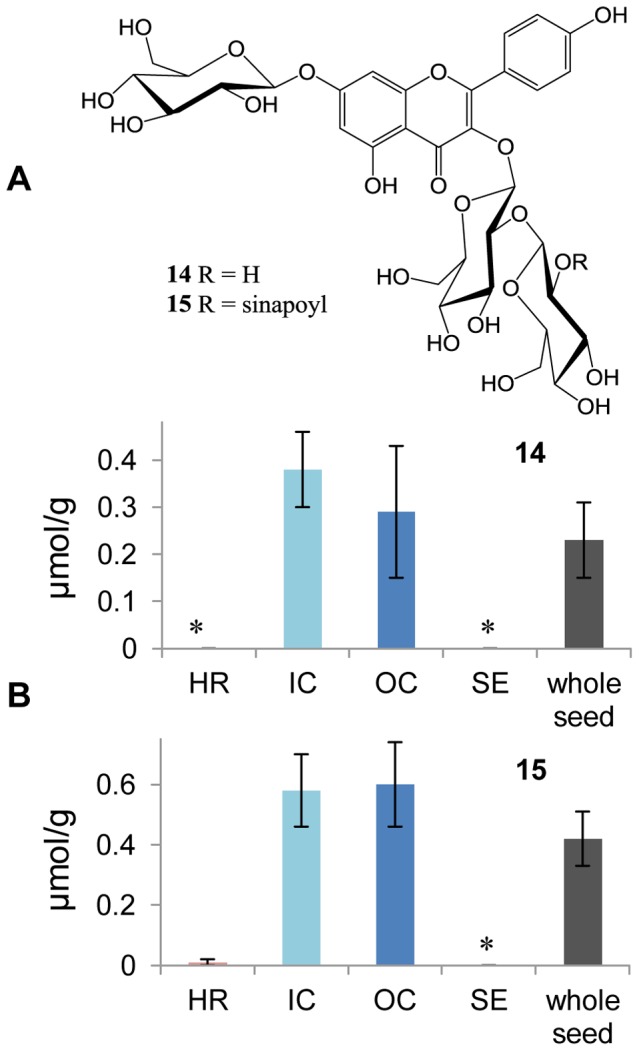
(A) Structures of two major flavonoids found in rapeseed: 14, kaempferol-3-O-β-D-glucopyranosyl-(1→2)-β-D-glucopyranoside-7-O-β-D-glucopyranoside; and 15, kaempferol-3-O-(2-O-sinapoyl)-β-D-glucopyranosyl-(1→2)-β-D-glucopyranoside-7-O-β-D-glucopyranoside. (B) Concentrations of 14 and 15 in different rapeseed tissues and whole rapeseed. HR, hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; and SE, seed coat and endosperm. Each column shows the mean of four replicates with standard error, and *means not detectable.
Flavonoids, which constitute an enormously diverse class of phenolic secondary metabolites, are involved in various physiological and ecological processes in plants [64]. A common function of flavonoids is protecting plants from UV-B irradiation [65], which was also demonstrated in rape [66], [67]. Here, the finding of flavonoid accumulation in the primordial tissue of the cotyledons (IC and OC) of mature rapeseed leads to the hypothesis that these compounds are preformed for protecting the chlorophyll and other light-sensitive components from UV-B irradiation in cotyledons emerging during germination. Flavonoids were clearly demonstrated to inhibit root formation [68], [69] by interfering with the transport of auxins from shoot to root [70]–[73]. Our finding that flavonoids are absent in hypocotyl and radicle (HR) fraction is consistent with this physiological phenomenon. Flavonoids also accumulate in seed coats to protect seeds against diverse biotic and abiotic stresses [74]. As in other seeds, proanthocyanidins accumulate in rapeseed coats. Responsible for the seed color, they are normally insoluble [75]. Oligomers and polymers are the probable reason why monomeric flavonoids were not detected in rapeseed hull tissue.
Tissue-specific Secondary Metabolites Biosynthesis in Rapeseed
The present results and previously reported metabolic profiling data on rapeseed [2], [23], [44], [45], [60], [67], [75], [79] suggest that expression of genes encoding enzymes of secondary metabolites biosynthetic pathways is different among rapeseed tissues. While the glucosinolates are evenly distributed in embryo tissues, and also occur in the seed hull, the phenolics, which all originate from the phenylpropanoid pathway, show tissue-specific distribution patterns disclosing diverse gene expression in rapeseed tissues. The biosynthetic pathways of major phenolics in rapeseed tissues are outlined in Figure 6. Sinapine is synthesized in the entire rapeseed, meanwhile, each tissue pursues its own biosynthetic pathway. Kaempferol glucosides accumulate in cotyledons, suggesting their biosynthesis in this tissue. Another class of flavonoids, the proanthocyanidins are produced in the seed coat [75], [76], the same site as in seeds of other plants [74]. The spermidine conjugate, which is exclusively accumulated in HR, implies that the corresponding biosynthetic pathway occurs only in HR part. The data presented here corroborate the working hypothesis, namely that different classes of secondary metabolites possessing individual biological functions indeed exist in specific tissues in rapeseed.
Figure 6. Hypothetical compartementation of phenolics biosynthetic pathways in different rapeseed tissues.

HR, hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; and SE, seed coat and endosperm.
Conclusion
Recent studies on the tissue-specific distribution of soluble primary metabolites such as lipids, amino acids, carbohydrates and polymers (starch) demonstrated the feasibility of the LMD-based chemical analysis of rapeseed organs [14]. The major primary metabolites in rapeseed embryo tissues are quantitatively but not qualitatively different, because these components are storage products and are involved in essential life cycles of plant growth and development. Unlike primary components, secondary metabolites help plants adapt to their biotic and abiotic environments [7], [8]. Seed tissues play different roles before and during germination, and develop into individual plant organs after germination. Therefore, secondary metabolites are speculated to accumulate unevenly in different seed tissues. The finding that some of the secondary metabolites detected in this work have different tissue-specific distribution patterns not only solidly supports this hypothesis but also offers the first clue to the biological functions of the secondary metabolites in the mature seed and probably during germination. The knowledge about the specific localization may be used to study the regulation of the biosynthesis and metabolic modification of secondary metabolites. On the other hand, the described sampling methodology, LMD, can be adjusted to facilitate the tissue-specific detection of metabolites, proteins and RNA in other plant materials.
Materials and Methods
Plant Material
Rapeseed (winter cultivar “Emerald”) used in this study was purchased from Raps GbR (Langballig, Germany). Entire seeds were used for analysis.
Laser Microdissection
The basic work flow of LMD and its application to plant tissue has been reported [15], [77]. Mature rapeseed was embedded vertically in Jung tissue freezing medium (Leica Microsystems GmbH, Nussloch, Germany), and immediately frozen in liquid nitrogen. Serial cryosections (60 µm thickness) were prepared at –24°C using a cryostat microtome (Leica CM1850, Bensheim, Germany) and directly mounted on PET-Membrane FrameSlides (MicroDissect GmbH, Herborn, Germany). LMD was performed on the Leica LMD 6000 laser microdissection system (Leica Microsystems GmbH, Wetzlar, Germany) equipped with a nitrogen solid state diode laser of a short pulse duration (355 nm). The cutting settings were as follows: 20×magnification, laser intensity of 128 (the strongest), laser moving speed of 1 (the slowest). The cut materials were collected in the cap of 0.5 ml centrifuge tubes by gravity and then transferred to an HPLC vial. The pictures were taken by a microscope-integrated camera HV-D20P (Hitachi, Tokyo, Japan). Rapeseed was dissected into four parts, HR, IC, OC, and SE (Figure 1), and weights, including the supporting PET membrane of the frame slide, which was unavoidably cut along with the plant tissue, are listed in Table 1.
Sample Preparation
Generally, each sample was separated into glucosinolate fraction and non-glucosinolate (NG) fraction for further analysis through the procedure adapted from the literature [78]. The four dissected tissue groups (HR, IC, OC, and SE) were extracted separately in an ultrasonic bath for 10 min with 1 ml 80% (v/v) MeOH, which contains 10 µM sinalbin as an internal standard for glucosinolates and 10 µM cinnamic acid choline ester (synthesized according to [79]) as an internal standard for sinapine. The weak anion exchange DEAE Sephadex cartridges (Sigma, Steinheim, Germany), which were conditioned with 800 µl H2O and equilibrated with 500 µl 80% (v/v) MeOH before use, were used to separate glucosinolates from the other compounds. Each sample (800 µl extract) was loaded to the cartridge and eluted with 500 µl 80% (v/v) MeOH. Eluate (1300 µl) was collected as an NG fraction and dried in a vacuum centrifuge evaporator Genevac HT-4X (Genevac Ltd, Suffolk, UK). Samples were reconstituted in 200 µl 20% (v/v) MeCN for NG analysis. The DEAE Sephadex cartridges were further eluted by 1 ml H2O twice and 500 µl 0.02 M 2-(N-morpholino)ethanesulfonic acid (MES) buffer (pH 5.2). Sulfatase (30 µl solution) (Sigma, Steinheim, Germany) was prepared as described in [80] and loaded onto the cartridge. The cartridges were capped, incubated at ambient temperature overnight, and eluted with 500 µl H2O for desulfated glucosinolate analysis.
Identification and Quantification of Glucosinolates
Desulfated glucosinolates were identified with HPLC-DAD/MS by comparing their mass spectrometric data and retention times with those of references [81]. The compounds were quantified based on an internal standard with DAD. HPLC was conducted on an Agilent series HP1100 (binary pump G1312A, autosampler G1367A, diode array detector G1315A; Agilent Technologies, Waldbronn, Germany). Chromatographic separation was performed on a LiChrospher RP18 column (5 µm, 250×4.6 mm, Merck, Darmstadt, Germany) with a guard column (5 µm, 4×4 mm) using a linear binary gradient of H2O (solvent A) containing 0.2% (v/v) formic acid (FA) and MeCN (solvent B), with a flow rate of 1.0 ml min−1 at 25°C as follows: 0 min: 1.5% B, 1 min: 1.5% B, 6 min: 5% B, 8 min: 7% B, 18 min: 21% B, 23 min: 29% B, 23.1 min: 100% B, 24 min: 100% B, 24.1 min: 1.5% B, and 28 min: 1.5% B. The injection volume was 50 µl. The absorption of HPLC eluate was monitored by DAD at 229 nm.
Identification and Quantification of Phenolics in the NG Fractions
HPLC-ESIMS was applied to quantify phenolics in laser-microdissected samples in NG fractions. The chromatographic separation was performed on a Nucleodur Sphinx RP column (5 µm, 250×4.6 mm; Macherey-Nagel GmbH, Düren, Germany) using the above-mentioned separation conditions (HPLC system, flow rate, temperature, and eluent) except a linear gradient, which was as follows: 0 min: 10% B, 20 min: 30% B, 25 min: 70% B, 25.1 min: 100% B, 28 min: 100% B, 28.1 min: 10% and 32 min: 10% B. The injection volume was 10 µl. Electrospray ionization mass spectra of HPLC eluate were monitored on an Esquire 6000 ion trap mass spectrometer (Bruker Daltonics, Bremen, Germany). Both positive and negative modes were used in the range m/z 150–1200 with skimmer voltage +/−40 V, capillary exit voltage +/−150 V, capillary voltage −/+4000 V, nebulizer pressure 35 psi, drying gas 10 L min−1, and gas temperature 350°C. Phenolics were identified based on their MS data and comparing the chromatographic retention times to those of compounds reported for rapeseed of cv. “Emerald” (unpublished data). The concentration of sinapine was calculated relative to the internal standard cinnamic acid choline ester in positive mode. The cyclic spermidine conjugate and two flavonoids were quantified using calibration curves in positive and negative modes, respectively.
Data Analysis
The experiments were performed in four replicates. Data are reported as means ± standard deviation (SD). Analyses of variance and significant differences among means were tested by one-way ANOVA using SPSS Statistics 17.0. The least significant difference at P = 0.05 level was calculated.
Supporting Information
Extracted ion chromatograms for the cyclic spermidine in different rapeseed tissues. Extracted ion chromatograms (EIC) for ions at m/z 496.4±0.5 measured in positive ionization mode of samples from different laser-microdissected rapeseed tissues. 13: Major cyclic spermidine conjugate (for structure, see Figure 4A ). ▾ major cyclic spermidine conjugate (13) peak.HR, hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; and SE, seed coat and endosperm.
(TIF)
Extracted ion chromatograms for the two major flavonoids in different rapeseed tissues. Extracted ion chromatograms (EIC) of samples from different rapeseed tissues measured in negative ionization mode for (A) ions at m/z 771.4±0.5 of flavonoid 14; and (B) ions at m/z 977.5±0.5 of flavonoid 15. For structures, see Figure 5A. HR, hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; and SE, seed coat and endosperm. ▾ peaks of flavonoid 14 in (A) and peaks of flavonoid 15 in (B).
(TIF)
Acknowledgments
We thank Emily Wheeler for editorial help.
Funding Statement
JF received a PhD scholarship by the International Max Planck Research School (IMPRS) “Exploration of Ecological Interactions with Molecular and Chemical Techniques”. The research was financially supported by the Max Planck Society (MPG). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Nesi N, Delourme R, Brégeon M, Falentin C, Renard M (2008) Genetic and molecular approaches to improve nutritional value of Brassica napus L. seed. CR Biol 331: 763–771. [DOI] [PubMed] [Google Scholar]
- 2. Wolfram K, Schmidt J, Wray V, Milkowski C, Schliemann W, et al. (2010) Profiling of phenylpropanoids in transgenic low-sinapine oilseed rape (Brassica napus). Phytochemistry 71: 1076–1084. [DOI] [PubMed] [Google Scholar]
- 3. Bones AM, Rossiter JT (1996) The myrosinase-glucosinolate system, its organisation and biochemistry. Physiol Plantarum 97: 194–208. [Google Scholar]
- 4. Fenwick GR (1982) The assessment of a new protein source - Rapeseed. Proc Nutr Soc 41: 277–288. [DOI] [PubMed] [Google Scholar]
- 5. Milkowski C, Strack D (2010) Sinapate esters in brassicaceous plants: biochemistry, molecular biology, evolution and metabolic engineering. Planta 232: 19–35. [DOI] [PubMed] [Google Scholar]
- 6. Strack D (1981) Sinapine as a supply of choline for the biosynthesis of phosphatidylcholine in Raphanus sativus seedlings. Z Naturforsch C 36: 215–221. [Google Scholar]
- 7. Wink M (2003) Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 64: 3–19. [DOI] [PubMed] [Google Scholar]
- 8. Hartmann T (2007) From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry 68: 2831–2846. [DOI] [PubMed] [Google Scholar]
- 9. Winde I, Wittstock U (2011) Insect herbivore counteradaptations to the plant glucosinolate-myrosinase system. Phytochemistry 72: 1566–1575. [DOI] [PubMed] [Google Scholar]
- 10. Bednarek P, Piślewska-Bednarek M, Svatoš A, Schneider B, Doubský J, et al. (2009) A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 323: 101–106. [DOI] [PubMed] [Google Scholar]
- 11. Li JY, Ou-Lee TM, Raba R, Amundson RG, Last RL (1993) Arabidopsis flavonoid mutants are hypersensitive to UV-B irradiation. Plant Cell 5: 171–179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Landry LG, Chapple CCS, Last RL (1995) Arabidopsis mutants lacking phenolic sunscreens exhibit enhanced ultraviolet-B injury and oxidative damage. Plant Physiol 109: 1159–1166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Li X, Bergelson J, Chapple C (2010) The ARABIDOPSIS accession Pna-10 is a naturally occurring sng1 deletion mutant. Mol Plant 3: 91–100. [DOI] [PubMed] [Google Scholar]
- 14. Schiebold S, Tschiersch H, Borisjuk L, Heinzel N, Radchuk R, et al. (2011) A novel procedure for the quantitative analysis of metabolites, storage products and transcripts of laser microdissected seed tissues of Brassica napus . Plant Methods 7: 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hölscher D, Schneider B (2008) Application of laser-assisted microdissection for tissue and cell-specific analysis of RNA, proteins, and metabolites. In: Lüttge UE, Beyschlag W, Murata J, editors. Progress in Botany. Berlin, Heidelberg: Springer. 141–167.
- 16. Nelson T, Gandotra N, Tausta SL (2008) Plant cell types: reporting and sampling with new technologies. Curr Opin Plant Biol 11: 567–573. [DOI] [PubMed] [Google Scholar]
- 17. Schmid MW, Schmidt A, Klostermeier UC, Barann M, Rosenstiel P, et al. (2012) A powerful method for transcriptional profiling of specific cell types in eukaryotes: Laser-assisted microdissection and RNA sequencing. PLoS One 7: e29685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Li SH, Schneider B, Gershenzon J (2007) Microchemical analysis of laser-microdissected stone cells of Norway spruce by cryogenic nuclear magnetic resonance spectroscopy. Planta 225: 771–779. [DOI] [PubMed] [Google Scholar]
- 19. Hölscher D, Schneider B (2007) Laser microdissection and cryogenic nuclear magnetic resonance spectroscopy: an alliance for cell type-specific metabolite profiling. Planta 225: 763–770. [DOI] [PubMed] [Google Scholar]
- 20. Nakashima J, Chen F, Jackson L, Shadle G, Dixon RA (2008) Multi-site genetic modification of monolignol biosynthesis in alfalfa (Medicago sativa): effects on lignin composition in specific cell types. New Phytol 179: 738–750. [DOI] [PubMed] [Google Scholar]
- 21. Obel N, Erben V, Schwarz T, Kuhnel S, Fodor A, et al. (2009) Microanalysis of plant cell wall polysaccharides. Mol Plant 2: 922–932. [DOI] [PubMed] [Google Scholar]
- 22. Abbott E, Hall D, Hamberger B, Bohlmann J (2010) Laser microdissection of conifer stem tissues: isolation and analysis of high quality RNA, terpene synthase enzyme activity and terpenoid metabolites from resin ducts and cambial zone tissue of white spruce (Picea glauca). BMC Plant Biol 10: 106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. El-Din Saad El-Beltagi H, Amin Mohamed A (2010) Variations in fatty acid composition, glucosinolate profile and some phytochemical contents in selected oil seed rape (Brassica napus L.) cultivars. Grasas Aceites 61: 143–150. [Google Scholar]
- 24. Thangstad OP, Gilde B, Chadchawan S, Seem M, Husebye H, et al. (2004) Cell specific, cross-species expression of myrosinases in Brassica napus, Arabidopsis thaliana and Nicotiana tabacum . Plant Mol Biol 54: 597–611. [DOI] [PubMed] [Google Scholar]
- 25. Agerbirk N, Olsen CE (2012) Glucosinolate structures in evolution. Phytochemistry 77: 16–45. [DOI] [PubMed] [Google Scholar]
- 26. Halkier BA, Gershenzon J (2006) Biology and biochemistry of glucosinolates. Annu Rev Plant Biol 57: 303–333. [DOI] [PubMed] [Google Scholar]
- 27. Kliebenstein DJ, Kroymann J, Mitchell-Olds T (2005) The glucosinolate-myrosinase system in an ecological and evolutionary context. Curr Opin Plant Biol 8: 264–271. [DOI] [PubMed] [Google Scholar]
- 28. Clossais-Besnard N, Larher F (1991) Physiological role of glucosinolates in Brassica napus. Concentration and distribution pattern of glucosinolates among plant organs during a complete life cycle. J Sci Food Agr 56: 25–38. [Google Scholar]
- 29. Petersen BL, Chen SX, Hansen CH, Olsen CE, Halkier BA (2002) Composition and content of glucosinolates in developing Arabidopsis thaliana . Planta 214: 562–571. [DOI] [PubMed] [Google Scholar]
- 30. Brown PD, Morra MJ, Mccaffrey JP, Auld DL, Williams L (1991) Allelochemicals produced during glucosinolate degradation in soil. J Chem Ecol 17: 2021–2034. [DOI] [PubMed] [Google Scholar]
- 31. Smolinska U, Morra MJ, Knudsen GR, Brown PD (1997) Toxicity of glucosinolate degradation products from Brassica napus seed meal toward Aphanomyces euteiches f. sp. pisi. Phytopathology 87: 77–82. [DOI] [PubMed] [Google Scholar]
- 32. Bending GD, Lincoln SD (2000) Inhibition of soil nitrifying bacteria communities and their activities by glucosinolate hydrolysis products. Soil Biol Biochem 32: 1261–1269. [Google Scholar]
- 33. Ishimoto H, Fukushi Y, Yoshida T, Tahara S (2000) Rhizopus and Fusarium are selected as dominant fungal genera in rhizospheres of Brassicaceae. J Chem Ecol 26: 2387–2399. [Google Scholar]
- 34. Lazzeri L, Manici LM (2001) Allelopathic effect of glucosinolate-containing plant green manure on Pythium sp and total fungal population in soil. Hortscience 36: 1283–1289. [Google Scholar]
- 35. Cohen MF, Yamasaki H, Mazzola M (2005) Brassica napus seed meal soil amendment modifies microbial community structure, nitric oxide production and incidence of Rhizoctonia root rot. Soil Biol Biochem 37: 1215–1227. [Google Scholar]
- 36. Bressan M, Roncato MA, Bellvert F, Comte G, Haichar FE, et al. (2009) Exogenous glucosinolate produced by Arabidopsis thaliana has an impact on microbes in the rhizosphere and plant roots. ISME J 3: 1243–1257. [DOI] [PubMed] [Google Scholar]
- 37. Lazzeri L, Curto G, Leoni O, Dallavalle E (2004) Effects of glucosinolates and their enzymatic hydrolysis products via myrosinase on the root-knot nematode Meloidogyne incognita (Kofoid et White) Chitw. J Agric Food Chem 52: 6703–6707. [DOI] [PubMed] [Google Scholar]
- 38. Zasada IA, Meyer SLF, Morra MJ (2009) Brassicaceous seed meals as soil amendments to suppress the plant-parasitic nematodes Pratylenchus penetrans and Meloidogyne incognita . J Nematol 41: 221–227. [PMC free article] [PubMed] [Google Scholar]
- 39. Kabouw P, van der Putten WH, van Dam NM, Biere A (2010) Effects of intraspecific variation in white cabbage (Brassica oleracea var. capitata) on soil organisms. Plant Soil 336: 509–518. [Google Scholar]
- 40. Rahman L, Whitelaw-Weckert MA, Orchard B (2011) Consecutive applications of brassica green manures and seed meal enhances suppression of Meloidogyne javanica and increases yield of Vitis vinifera cv Semillon. Appl Soil Ecol 47: 195–203. [Google Scholar]
- 41. Brown PD, Morra MJ (1996) Hydrolysis products of glucosinolates in Brassica napus tissues as inhibitors of seed germination. Plant Soil 181: 307–316. [Google Scholar]
- 42. Bainard LD, Brown PD, Upadhyaya MK (2009) Inhibitory effect of tall hedge mustard (Sisymbrium loeselii) allelochemicals on rangeland plants and arbuscular mycorrhizal fungi. Weed Sci 57: 386–393. [Google Scholar]
- 43. Hu P, Wang AS, Engledow AS, Hollister EB, Rothlisberger KL, et al. (2011) Inhibition of the germination and growth of Phymatotrichopsis omnivora (cotton root rot) by oilseed meals and isothiocyanates. Appl Soil Ecol 49: 68–75. [Google Scholar]
- 44. Liu Q, Wu L, Pu HM, Li CY, Hu QH (2012) Profile and distribution of soluble and insoluble phenolics in Chinese rapeseed (Brassica napus). Food Chem 135: 616–622. [DOI] [PubMed] [Google Scholar]
- 45. Bouchereau A, Hamelin J, Lamour I, Renard M, Larher F (1991) Distribution of sinapine and related compounds in seeds of Brassica and allied genera. Phytochemistry 30: 1873–1881. [Google Scholar]
- 46. Clauß K, Baumert A, Nimtz M, Milkowski C, Strack D (2008) Role of a GDSL lipase-like protein as sinapine esterase in Brassicaceae. Plant J 53: 802–813. [DOI] [PubMed] [Google Scholar]
- 47. Clauß K, von Roepenack-Lahaye E, Böttcher C, Roth MR, Welti R, et al. (2011) Overexpression of sinapine esterase BnSCE3 in oilseed rape seeds triggers global changes in seed metabolism. Plant Physiol 155: 1127–1145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Sheahan JJ (1996) Sinapate esters provide greater UV-B attenuation than flavonoids in Arabidopsis thaliana (Brassicaceae). Am J Bot 83: 679–686. [Google Scholar]
- 49. Jin HL, Cominelli E, Bailey P, Parr A, Mehrtens F, et al. (2000) Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in Arabidopsis . EMBO J 19: 6150–6161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Meißner D, Albert A, Böttcher C, Strack D, Milkowski C (2008) The role of UDP-glucose: hydroxycinnamate glucosyltransferases in phenylpropanoid metabolism and the response to UV-B radiation in Arabidopsis thaliana . Planta 228: 663–674. [DOI] [PubMed] [Google Scholar]
- 51. Lake JA, Field KJ, Davey MP, Beerling DJ, Lomax BH (2009) Metabolomic and physiological responses reveal multi-phasic acclimation of Arabidopsis thaliana to chronic UV radiation. Plant Cell Environ 32: 1377–1389. [DOI] [PubMed] [Google Scholar]
- 52. Demkura PV, Ballaré CL (2012) UVR8 mediates UV-B-induced Arabidopsis defense responses against Botrytis cinerea by controlling sinapate accumulation. Mol Plant 5: 642–652. [DOI] [PubMed] [Google Scholar]
- 53. Tzagoloff A (1963) Metabolism of sinapine in mustard plants. I. Degradation of sinapine into sinapic acid and choline. Plant Physiol 38: 202–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Bopp M, Ludicke W (1975) Degradation of sinapine during early development of Sinapis alba L. Z Naturforsch C. 30: 663–667. [Google Scholar]
- 55. Bienz S, Bisegger P, Guggisberg A, Hesse M (2005) Polyamine alkaloids. Nat Prod Rep 22: 647–658. [DOI] [PubMed] [Google Scholar]
- 56. Luo J, Fuell C, Parr A, Hill L, Bailey P, et al. (2009) A novel polyamine acyltransferase responsible for the accumulation of spermidine conjugates in Arabidopsis seed. Plant Cell 21: 318–333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Kusano T, Berberich T, Tateda C, Takahashi Y (2008) Polyamines: essential factors for growth and survival. Planta 228: 367–381. [DOI] [PubMed] [Google Scholar]
- 58. Alcázar R, Altabella T, Marco F, Bortolotti C, Reymond M, et al. (2010) Polyamines: molecules with regulatory functions in plant abiotic stress tolerance. Planta 231: 1237–1249. [DOI] [PubMed] [Google Scholar]
- 59. Kaur H, Heinzel N, Schöttner M, Baldwin IT, Gális I (2010) R2R3-NaMYB8 regulates the accumulation of phenylpropanoid-polyamine conjugates, which are essential for local and systemic defense against insect herbivores in Nicotiana attenuata . Plant Physiol 152: 1731–1747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Baumert A, Milkowski C, Schmidt J, Nimtz M, Wray V, et al. (2005) Formation of a complex pattern of sinapate esters in Brassica napus seeds, catalyzed by enzymes of a serine carboxypeptidase-like acyltransferase family? Phytochemistry 66: 1334–1345. [DOI] [PubMed] [Google Scholar]
- 61. Imai A, Matsuyama T, Hanzawa Y, Akiyama T, Tamaoki M, et al. (2004) Spermidine synthase genes are essential for survival of Arabidopsis. Plant Physiol 135: 1565–1573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Gill SS, Tuteja N (2010) Polyamines and abiotic stress tolerance in plants. Plant Signal Behav 5: 26–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Wimalasekera R, Tebartz F, Scherer GFE (2011) Polyamines, polyamine oxidases and nitric oxide in development, abiotic and biotic stresses. Plant Sci 181: 593–603. [DOI] [PubMed] [Google Scholar]
- 64.Croteau R, Kutchan TM, Lewis NG (2000) Natural products (Secondary metabolites); Buchanan BB, Gruissem W, Jones RL, editors. Rochville: American society of plant physiologist.
- 65. Harborne JB, Williams CA (2000) Advances in flavonoid research since 1992. Phytochemistry 55: 481–504. [DOI] [PubMed] [Google Scholar]
- 66. Wilson MI, Greenberg BM (1993) Protection of the D1 photosystem-II reaction center protein from degradation in ultraviolet-radiation following adaptation of Brassica napus L. to growth in ultraviolet-B. Photochem Photobiol 57: 556–563. [Google Scholar]
- 67. Wilson KE, Wilson MI, Greenberg BM (1998) Identification of the flavonoid glycosides that accumulate in Brassica napus L. cv. Topas specifically in response to ultraviolet B radiation. Photochem Photobiol 67: 547–553. [Google Scholar]
- 68. Imin N, Nizamidin M, Wu T, Rolfe BG (2007) Factors involved in root formation in Medicago truncatula . J Exp Bot 58: 439–451. [DOI] [PubMed] [Google Scholar]
- 69. Rusak G, Cerni S, Polancec DS, Ludwig-Müller J (2010) The responsiveness of the IAA2 promoter to IAA and IBA is differentially affected in Arabidopsis roots and shoots by flavonoids. Biol Plantarum 54: 403–414. [Google Scholar]
- 70. Brown DE, Rashotte AM, Murphy AS, Normanly J, Tague BW, et al. (2001) Flavonoids act as negative regulators of auxin transport in vivo in Arabidopsis . Plant Physiol 126: 524–535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Murphy A, Peer WA, Taiz L (2000) Regulation of auxin transport by aminopeptidases and endogenous flavonoids. Planta 211: 315–324. [DOI] [PubMed] [Google Scholar]
- 72. Peer WA, Bandyopadhyay A, Blakeslee JJ, Makam SI, Chen RJ, et al. (2004) Variation in expression and protein localization of the PIN family of auxin efflux facilitator proteins in flavonoid mutants with altered auxin transport in Arabidopsis thaliana . Plant Cell 16: 1898–1911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Peer WA, Murphy AS (2007) Flavonoids and auxin transport: modulators or regulators? Trends Plant Sci 12: 556–563. [DOI] [PubMed] [Google Scholar]
- 74. Lepiniec L, Debeaujon I, Routaboul JM, Baudry A, Pourcel L, et al. (2006) Genetics and biochemistry of seed flavonoids. Annu Rev Plant Biol 57: 405–430. [DOI] [PubMed] [Google Scholar]
- 75. Auger B, Marnet N, Gautier V, Maia-Grondard A, Leprince F, et al. (2010) A detailed survey of seed coat flavonoids in developing seeds of Brassica napus L. J Agric Food Chem. 58: 6246–6256. [DOI] [PubMed] [Google Scholar]
- 76. Debeaujon I, Nesi N, Perez P, Devic M, Grandjean O, et al. (2003) Proanthocyanidin-accumulating cells in Arabidopsis testa: Regulation of differentiation and role in seed development. Plant Cell 15: 2514–2531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Moco S, Schneider B, Vervoort J (2009) Plant micrometabolomics: The analysis of endogenous metabolites present in a plant cell or tissue. J Proteome Res 8: 1694–1703. [DOI] [PubMed] [Google Scholar]
- 78. Burow M, Müller R, Gershenzon J, Wittstock U (2006) Altered glucosinolate hydrolysis in genetically engineered Arabidopsis thaliana and its influence on the larval development of Spodoptera littoralis . J Chem Ecol 32: 2333–2349. [DOI] [PubMed] [Google Scholar]
- 79. Böttcher C, von Roepenack-Lahaye E, Schmidt J, Clemens S, Scheel D (2009) Analysis of phenolic choline esters from seeds of Arabidopsis thaliana and Brassica napus by capillary liquid chromatography/electrospray-tandem mass spectrometry. J Mass Spectrom 44: 466–476. [DOI] [PubMed] [Google Scholar]
- 80. Graser G, Oldham NJ, Brown PD, Temp U, Gershenzon J (2001) The biosynthesis of benzoic acid glucosinolate esters in Arabidopsis thaliana . Phytochemistry 57: 23–32. [DOI] [PubMed] [Google Scholar]
- 81. Reichelt M, Brown PD, Schneider B, Oldham NJ, Stauber E, et al. (2002) Benzoic acid glucosinolate esters and other glucosinolates from Arabidopsis thaliana . Phytochemistry 59: 663–671. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Extracted ion chromatograms for the cyclic spermidine in different rapeseed tissues. Extracted ion chromatograms (EIC) for ions at m/z 496.4±0.5 measured in positive ionization mode of samples from different laser-microdissected rapeseed tissues. 13: Major cyclic spermidine conjugate (for structure, see Figure 4A ). ▾ major cyclic spermidine conjugate (13) peak.HR, hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; and SE, seed coat and endosperm.
(TIF)
Extracted ion chromatograms for the two major flavonoids in different rapeseed tissues. Extracted ion chromatograms (EIC) of samples from different rapeseed tissues measured in negative ionization mode for (A) ions at m/z 771.4±0.5 of flavonoid 14; and (B) ions at m/z 977.5±0.5 of flavonoid 15. For structures, see Figure 5A. HR, hypocotyl and radicle; IC, inner cotyledon; OC, outer cotyledon; and SE, seed coat and endosperm. ▾ peaks of flavonoid 14 in (A) and peaks of flavonoid 15 in (B).
(TIF)