

Abstract
Global biodiversity losses provide an immediate impetus to elucidate the relationships between biodiversity, productivity and stability. In this study, we quantified the effects of species richness and species combination on the productivity and stability of phytoplankton communities subject to predation by a single rotifer species. We also tested one mechanism of the insurance hypothesis: whether large, slow-growing, potentially-defended cells would compensate for the loss of small, fast-growing, poorly-defended cells after predation. There were significant effects of species richness and species combination on the productivity, relative yield, and stability of phytoplankton cultures, but the relative importance of species richness and combination varied with the response variables. Species combination drove patterns of productivity, whereas species richness was more important for stability. Polycultures containing the most productive single species, Dunaliella, were consistently the most productive. Yet, the most species rich cultures were the most stable, having low temporal variability in measures of biomass. Polycultures recovered from short-term negative grazing effects, but this recovery was not due to the compensation of large, slow-growing cells for the loss of small, fast-growing cells. Instead, polyculture recovery was the result of reduced rotifer grazing rates and persisting small species within the polycultures. Therefore, although an insurance effect in polycultures was found, this effect was indirect and unrelated to grazing tolerance. We hypothesize that diverse phytoplankton assemblages interfered with efficient rotifer grazing and that this “interference effect” facilitated the recovery of the most productive species, Dunaliella. In summary, we demonstrate that both species composition and species richness are important in driving patterns of productivity and stability, respectively, and that stability in biodiverse communities can result from an alteration in consumer functioning. Our findings underscore the importance of predator-prey dynamics in determining the relationships between biodiversity, productivity and stability in producer communities.
Introduction
With unprecedented species extinction rates [1], [2], [3] and concomitant changes in ecosystem functioning [4], [5], [6] worldwide, there is a need to understand the relationships between biodiversity, productivity and stability at different trophic levels. Although there is no simple relationship between productivity and species richness [7], it is generally accepted that productivity and stability increase with biodiversity at the community level [8], [9], [10], [11], [12]. One mechanism that has been proposed to explain positive biodiversity-productivity relationships is niche partitioning, whereby species with different morphological or physiological characteristics can use different resources thus increasing overall productivity in species rich systems [13], [14], [15], [16]. Extending this concept from productivity to stability, a functionally diverse group of taxa may be more resistant or resilient to environmental or biological perturbations because different species exhibit different tolerances and thus responses to changes in environmental and biological factors. This idea, termed the “insurance” hypothesis or effect [17], [18], [19], has received some strong [10] but not unequivocal support [9], [20], [21].
In phytoplankton communities, functional diversity can arise when species exhibit different traits along a few key axes, namely light or nutrient utilization and susceptibility to predators. Major taxonomic groups (e.g. diatoms, dinoflagellates, cyanobacteria) tend to differ in their mean functional trait values, despite physiological or morphological plasticity within groups [22]. It follows that communities with representative species from diverse taxonomic groups may be more productive or stable than communities with species from fewer groups. Indeed, experimental manipulations [23], as well as analyses of field-collected data [24], [25], support niche partitioning as a mechanism that underlies positive biodiversity-productivity relationships in phytoplankton communities. Yet, there has not been consistently strong support for positive biodiversity-productivity relationships in experimental phytoplankton communities [21], [26]. Furthermore, little attention has been devoted to understanding stability resulting from functional traits within phytoplankton communities.
Here we describe the relationships between biodiversity, productivity and stability within phytoplankton communities, and we test the insurance hypothesis in the context of grazing tolerances. Specifically, we test the hypothesis that grazing-tolerant cells will compensate for the predatory loss of poorly-defended cells, resulting in greater productivity and stability, defined temporally, of polycultures compared to monocultures. There are two novel aspects to this study. First, to study stability we impose a biological perturbation. Previous studies that investigated stability of phytoplankton communities used environmental perturbations [21], [27]. The outcomes of biological perturbations, likely more complex than environmental perturbations, have particular relevance to understanding natural populations that are also subject to changing pressures from predation, disease, competition, immigration, or emigration. Second, we test compensatory dynamics explicitly by creating phytoplankton polycultures with species that are likely to differ in their ability to resist predation. With this design, we show that algal polycultures are more productive and stable than algal monocultures, due to indirect predator-mediated effects. This finding highlights the importance of predator-prey dynamics in understanding the role of biodiversity in producer communities.
Methods
Study Organisms and Culture Conditions
Our study organisms included six phytoplankton species that were grouped into two functional groups according to growth rate and susceptibility to grazing (Table 1) by Brachionus plicatilis, a brachionid rotifer that suspension feeds by drawing particles into its corona with ciliated buccal fields. Relative phytoplankton growth rate was measured directly with laboratory cultures, and grazing tolerance was assessed based on morphology. We considered the spines of Chaetoceros, chain-formation of Chaetoceros and Melosira, size of Coscinodiscus, and silica frustules of all three larger species (Table 1) to be morphological defenses that would deter or prohibit grazing by the rotifer. Although B. plicatilis is capable of consuming particles up to 58 µm, siliceous frustules are thought to be effective defenses against predators for diatoms [28], [29], [30]. All species except Nannochloris oculata, which was already in culture, were obtained from the National Center for Marine Algae and Microbiota (formerly the Culture Center for Marine Phytoplankton, Bigelow, ME). Cultures were acclimated through consecutive batch transfers to one media type (f/2 with artificial seawater [31], [32], [33]), temperature (19°C) and light cycle (16 h light:8 h dark by cool white fluorescent lights that supplied 300 µmol photons m−2 s−1). Before starting the experiment, cultures were maintained in 250 mL flasks for approximately two months in a single incubator (I-36VL, Percival Inc., Perry IA). Although phytoplankton cultures were not axenic when the experiment was started, bacterial abundance in the non-axenic cultures was low (<1% of total biomass).
Table 1. Classification and key morphological characteristics of the study species.
| Species | ID # | Division | Class | Size | Key Characteristics |
| fast-growing, susceptible to grazing | |||||
| Nannochloris oculata | –– | Chlorophyta | Chlorophyceae | 2–3 µm | non-motile, green to yellow |
| Dunalliela tertiolecta | 1320 | Chlorophyta | Chlorophyceae | 6–11 µm | biflagellate, green pigments at posterior |
| Rhodomonas sp. | 768 | Cryptophyta | Cryptophyceae | 11–16 µm | biflagellate, red to light brown |
| slow-growing, potentially defended against grazers | |||||
| Chaetoceros decipiens | 173 | Heterokontophyta | Bacillariophyceae | 10–26 µm | straight chains, long setae, yellow to brown |
| Melosira octogona_cf | 483 | Heterokontophyta | Coscinodiscophyceae | 19–25 µm | cylindrical cells, chain-forming, yellow to brown |
| Coscinodiscus sp. | 1583 | Heterokontophyta | Coscinodiscophyceae | 75–90 µm | solitary, thick frustules, yellow to brown |
Species were divided into two functional groups, as indicated. Identification numbers from the National Center for Marine Algae and Microbiota (formerly the CCMP) are listed. Measurements of cell length and width or diameter and height were made on at least 25 cells per species. Size ranges encompass all dimensions.
Experimental Design
The experimental design included three richness treatments (two, four and six species), within which species combinations were nested (Table 2), plus monoculture controls. Species combinations were created by haphazardly selecting species from each of the two functional groups such that functional groups were equally represented in all treatments. Experimental cultures were inoculated using a replacement design, with the initial cell abundance constant across levels of richness and species combinations. This experiment was a follow-up experiment to one conducted in which starting biovolume was constant. As both experiments yielded similar results, we do not present our preliminary data here. Our design resulted in equal average starting biovolume (1.37 × 107 µm3/mL) across richness treatments because species occurred in the experimental design an equal number of times (Table 2). However, biovolume varied between species combinations, from 1.4 × 105 µm3/mL in the Chaetoceros and Nannochloris treatment to 4 × 107 µm3/mL in the Coscinodiscus and Dunaliella treatment. As such, initial biovolume was used as a covariate in statistical analyses (see “Data Analyses”). Inoculum was added to 20 mL glass tubes (16 mm outer diameter) with 15 mL f/2 media, resulting in an initial cell abundance of 170 cells/mL. All tubes were placed in an angled tissue culture roller drum (TC-7, New Brunswick Scientific, Edison NJ), which rotated at 15 rpm, in the same incubator in which stock cultures were maintained. Before sampling, cultures were grown for approximately two weeks to allow communities to assemble and populations to reach measurable abundance.
Table 2. Experimental design of richness and species combination treatments, with the number of replicates as indicated.
| Richness | Species Combinations |
| (a) Chaetoceros & Nannochloris [n = 3] | |
| 2 [n = 9] | (b) Melosira & Rhodomonas [n = 3] |
| (c) Coscinodiscus & Dunaliella [n = 3] | |
| (d) Chaetoceros, Melosira, Nannochloris & Rhodomonas [n = 3] | |
| 4 [n = 9] | (e) Coscinodiscus, Dunaliella, Melosira & Nannochloris [n = 3] |
| (f) Chaetoceros, Coscinodiscus, Dunaliella & Rhodomonas [n = 3] | |
| 6 [n = 9] | (g) all species combined [n = 9]Each monoculture control was grown in triplicate. |
After the two-week growth period, we measured net community production and sampled cultures to estimate total community biovolume every four days for 12 days (see “Response Variables”). Immediately after the third sampling point, we added 15 rotifers (L-type, B. plicatilis, Reed Mariculture, Campbell CA) to each culture tube. Before this predator addition, the rotifers were starved for a day and rinsed in sterile seawater to reduce the introduction of bacteria to the phytoplankton cultures. After rotifer addition, cultures were sampled for the aforementioned variables, as well as rotifer abundance, every four days for an additional 12 days.
Response Variables
Net community production was determined by direct measurement of O2 evolution in the headspace of culture vessels [34]. Prior to incubations, cultures were bubbled with the reference gas for two minutes to prevent carbon limitation during the incubation period and establish a baseline O2 concentration. After bubbling, cultures were incubated for approximately four hours and final concentrations of O2 in the headspace of the tubes were measured by flushing air from the headspace into an Oxzilla II Differential Oxygen Analyzer (Sable Systems, Henderson, NV) during a three-minute sampling period. After measuring O2 production, one milliliter (<7% of culture volume) of each culture was preserved with Lugol’s Iodine, and fresh media was used to replace the sampled volume. Preserved cells were enumerated under light and phase contrast microscopy (BH-2, Olympus, Central Valley PA). At least 400 cells and comparable numbers of each species per sample were counted (e.g. 200, 100, and 70 cells of each species in the two-, four-, and six-species combinations, respectively). Chaetoceros, Rhodomonas and Melosira were counted at 200X and Coscinodiscus at 100X using a gridded Sedgewick-Rafter chamber (1801–G20, Wildlife Supply Company, Yulee FL). Dunaliella and Nannochloris were counted in a Neubauer hemocytometer (Marienfeld GMBH & Co., Germany) at 200X and 400X, respectively. To estimate total biovolume, species abundance was multiplied by mean biovolume, calculated before the experiment using measurements of>25 cells per species and formulas of Hillebrand et al. [35]. Measurements made at the end of the experiment confirmed that the mean biovolume of each species had not changed. After rotifer addition, all rotifers in the samples were enumerated.
Data Analyses
To determine the relative yield (RY) of polycultures compared to monocultures before rotifer addition, deviation from total yield, DT sensu Loreau [36], was calculated for O2 evolution and biovolume. DT is a standardized metric of RY that allows an overall comparison of polyculture productivities across levels of richness. DT was calculated as 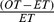 where OT =
where OT = 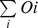 and ET =
and ET = 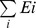 are the observed and expected yields, respectively, of polycultures with i species. Positive values of DT imply overyielding and negative values of DT imply underyielding, relative to the expected productivity of polycultures based on monoculture performance. Compared to other metrics, DT ignores shifts in the numerical dominance of individual species and is used as a rough tool to assess effects of biodiversity [36].
are the observed and expected yields, respectively, of polycultures with i species. Positive values of DT imply overyielding and negative values of DT imply underyielding, relative to the expected productivity of polycultures based on monoculture performance. Compared to other metrics, DT ignores shifts in the numerical dominance of individual species and is used as a rough tool to assess effects of biodiversity [36].
Temporal variability of O2 production and total biovolume was used to evaluate the stability of cultures. Stability here refers to constancy of phytoplankton biomass through time [8], [37] and is inversely related to temporal variability. We calculated temporal variability as the coefficient of variation (CV, σ/μ) of O2 production and biovolume through time (i.e., for each culture tube, a single CV was calculated from the six sampling points). With these data, we then calculated the mean CV for each of the four richness treatments and the mean CV for each of the 13 species combinations. Therefore, the number of replicates per treatment had no influence on the mean CV calculated. Temporal CV was calculated using transformed values of O2 production, as some values were negative towards the end of the experiment. In this paper, we restrict our analyses to experiment-long variability, as the temporal CVs for different periods (i.e. pre-rotifer addition, post-rotifer addition, experiment-long) were similar.
To test for effects of species richness and species combination on net oxygen production, total biovolume, RY and temporal CV we used mixed-model, nested ANOVA, in which species combination (a random factor) was nested within levels of species richness (a fixed factor). Separate ANOVA were conducted for each response variable, and initial biovolume was used as a covariate in the analyses. Because many of the raw data were non-normal, analyses were conducted on log- or rank-transformed data [38], [39]. O2 production was not standardized to biomass, as rotifer consumption of O2 confounded such standardization. Model diagnostics of residual plots, as suggested by Quinn & Keough [39], were performed for all analyses. To reduce the overall Type I error rate associated with multiple comparisons, the Dunn-Sidak method was used to adjust the α-value for post-hoc Tukey tests [40]. Analyses and model diagnostics were performed in Statistica 10.
To compare the relative importance of species richness and species combination on productivity and stability, we applied the approach suggested by Connolly et al. [41]. Briefly, the importance of species richness (the fixed effect) is defined as the slope of the richness gradient, scaled by the range of richness used in the experiment as 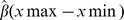 . The importance of species combination is defined as the absolute size of the difference between two randomly selected compositions, or 1.128(σsc) where σsc is the variance component for species combination. For this comparison, the parameters
. The importance of species combination is defined as the absolute size of the difference between two randomly selected compositions, or 1.128(σsc) where σsc is the variance component for species combination. For this comparison, the parameters 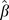 and σsc were taken directly from the results of the mixed-model analysis (i.e. prediction equation of model and variance component of species combination, respectively).
and σsc were taken directly from the results of the mixed-model analysis (i.e. prediction equation of model and variance component of species combination, respectively).
Finally, the additive partitioning methods of Loreau and Hector [42], using species biovolume, were used to separate the effects of complementarity and selection during the experiment. Positive complementarity, resulting from resource partitioning or positive species interactions, is measured as a positive change in polyculture RY compared to the weighted average of the component species’ monoculture yields. In contrast, positive selection, which occurs when a highly productive species dominates in a polyculture, is indicated by positive covariance between the monoculture yield of a species and its change in RY within the polyculture. Either effect can be positive or negative and the sum of the effects is the net biodiversity effect [42].
Results
Productivity and Relative Yield
There were significant effects of both species richness and species combination on O2 production and total biovolume (Table 3). In general, productivity increased with species richness and the six-species polycultures were the most productive (Fig. 1). However, species combination was approximately four to six times more important than species richness to productivity (Table 3). This strong species combination effect was due to a single, highly productive species: Dunaliella tertiolecta. Post-hoc comparisons showed that the biovolume of all polycultures containing Dunaliella was significantly higher than that of the other polycultures and monocultures except the Dunaliella monoculture (F(9) = 17.84, p<0.0006). Post-hoc comparisons for O2 production were similar to those of biovolume (data not shown). Relative Yield (RY) of both O2 production and total biovolume before rotifer addition was positive in all polycultures except in the two-species polyculture of Chaetoceros and Nannochloris (Fig. 2). In the six-species polycultures, RY was highest. There were significant effects of both richness and species combination on the RY of total biovolume, but not on RY of O2 production (Table 3, Fig. 2).
Table 3. Results of mixed model ANOVA comparing the effects of species richness and combination on oxygen production, total phytoplankton biovolume, relative yield (RY), and temporal variability (CV).
| response | effect | dominant | ratio | |
| variable | species richness | species combination | variable | |
| O2 production | F(3) = 50.39, p<0.001 | F(9) = 21.03, p<0.001 | combination | 5.8 |
| 1<2,4<6 | ||||
| biovolume | F(3) = 70.91, p<0.001 | F(9) = 17.84, p<0.001 | combination | 4.5 |
| 1<2, 4; 1,2<6 | ||||
| RY (O2 production) | F(2) = 2.58, p = 0.078 | F(4) = 1.27, p = 0.285 | ns | ns |
| RY (biovolume) | F(2) = 8.78, p<0.001 | F(4) = 4.39, p = 0.002 | combination | 2 |
| 2<6 | ||||
| CV (O2 production) | F(3) = 41.88, p<0.001 | F(9) = 4.60, p<0.001 | richness | 43 |
| 1>2,4,6; 2>6 | ||||
| CV (biovolume) | F(3) = 10.14, p<0.001 | F(9) = 4.43, p<0.001 | richness | 18 |
| 1>2,4,6 | ||||
Post-hoc Tukey results for effects of richness are expressed in the table as inequalities. See the text for post-hoc comparisons between species combinations. The dominant variable in each ANOVA derived following [41] and the ratio by which it is dominant over the other main factor are listed.
Figure 1. Net oxygen production (A) and total biovolume (B) of phytoplankton monocultures and polycultures.

Each box displays the experiment-long median (line within box), 25th and 75th percentiles (box boundaries), 10th and 90th percentiles (lower and upper error bars) and 5th and 95% percentiles (dots). Medians and percentiles were calculated using all data collected throughout the experiment (n = 54 for polycultures, 108 for monocultures). Net oxygen production in the monocultures was centered on zero due to the consumption of oxygen after rotifer addition (see Figure 3A).
Figure 2. Relative yield of net oxygen production (A) and total biovolume (B).

The main plots compare species combinations whereas the insets compare levels of species richness. Bars represent experiment-long means (± SE) calculated for each replicate and sampling day. Letters above each bar indicate the results of the post-hoc Tukey tests associated with ANOVA. Species combinations are listed with each plot, using the following abbreviations: D – Dunaliella, N – Nannochloris, R – Rhodomonas, Ch – Chaetoceros, Co – Coscinodiscus, M – Melosira and all – all species in the six-species combination.
Stability
Monocultures and polycultures exhibited extremely different responses to rotifer addition. There were short-term negative effects of rotifers on polyculture biomass, but persistent negative effects on monocultures (Fig. 3, S1). The post-rotifer decline in monoculture biomass magnified the pre-rotifer positive richness effects found on O2 production and total biovolume. For example, O2 production was more similar across treatments before rotifer addition than after rotifer addition. After rotifer addition, values of net O2 production diverged, resulting in O2 consumption in monocultures (Fig. 3A). Similar divergence was apparent in total biovolume. On the sampling date before rotifer addition, total biovolume varied less than three-fold between monocultures and polycultures (compare across Fig. 3B, C, D, E). In contrast, on the last sampling date, the differences in total biovolume between polycultures and monocultures were orders of magnitude.
Figure 3. Oxygen production, phytoplankton biovolume and rotifer abundance through time in the different richness treatments.

The top figure (A) shows the mean (± SE) net oxygen production of the different richness treatments. The bottom figures show total phytoplankton biovolume (symbols) and rotifer abundance (bars) of the one- (B), two- (C), four- (D) and six-species (e) treatments before rotifer addition (first three points) and after rotifer addition (last three points). Points and bars in b-e represent means ± standard errors (n = nine for polycultures, 18 for monocultures). The arrow indicates the point of rotifer addition at an initial abundance of one rotifer per mL. Rotifer abundance is shown on a log scale.
Rotifers grazed the monocultures of Dunaliella, Nannochloropsis, Rhodomonas and Melosira to or nearly to depletion (Fig. 4). Rotifer abundance was highest and the grazing effects most dramatic in Dunaliella monocultures. The monocultures of Chaetoceros and Coscinodiscus crashed after rotifer addition, but this decline in biomass was not coincident with an increase in rotifer abundance (Fig. 4). Each of the polycultures except the two-species combination of Chaetoceros and Nannochloris recovered from the negative effects of rotifers (Fig. 4). Rotifer abundance was lowest in the six-species polycultures.
Figure 4. Mean (± SE) phytoplankton biovolume (symbols) and rotifer abundance (bars) of species combinations through time.

The arrows indicate the point of rotifer addition at an initial abundance of one rotifer per mL. Species combinations are listed with each plot, using the following abbreviations: D – Dunaliella, N – Nannochloris, R – Rhodomonas, Ch – Chaetoceros, Co – Coscinodiscus, M – Melosira, all – all species in the six-species combination. All primary y-axes are identical with the exception of the y-axis in the row of plots second from the top. Rotifer abundance is shown on a log scale.
Consistent with patterns of species dynamics in polycultures and monocultures, species richness was the dominant variable affecting temporal variability of O2 production and total biovolume (Table 3). The effects of species richness were ∼20- to 40-fold greater than the effects of species combination; indeed, O2 production and biovolume in the one-species treatments were generally more variable than the two-, four- and six-species richness treatments (Fig. 5). Still, the effects of species combination were also apparent. For example, the temporal CV of Chaetoceros biovolume in monoculture was the highest of all coefficients, consistent with its population crash during the experiment (Fig. 4, 5).
Figure 5. Temporal CV of net oxygen production (A) and total biovolume (B).

The main plots compare species combinations whereas the insets compare levels of species richness. Bars represent experiment-long means (± SE) calculated for each replicate and sampling day. Note that the temporal CV was calculated for single culture tubes, such that the mean values for richness treatments and species combinations are not influenced by the number of replicates per treatment. Letters above each bar indicate the results of the post-hoc Tukey tests associated with ANOVA. Species combinations are listed with each plot, using the following abbreviations: D – Dunaliella, N – Nannochloris, R – Rhodomonas, Ch – Chaetoceros, Co – Coscinodiscus, M – Melosira and all – all species in the six-species combination.
Species Dynamics
Within the polycultures in which Dunaliella was present, it accounted for 65–90% of total biovolume and thus dominated patterns of species dynamics (Fig. S2B, C). In polycultures without Dunaliella, the smaller species were also consistently dominant: Rhodomonas in combination with Melosira (Fig. S2A); Nannochloris in combination with Chaetoceros (data not shown); and Rhodomonas in combination with Chaetoceros, Melosira and Nannochloris (data not shown). The biovolume of the larger taxa (i.e. Chaetoceros, Coscinodiscus and Melosira) was relatively constant before and after rotifer addition (Fig. S2A, B, C), in strong contrast to predictions of the insurance hypothesis based on differences in grazing tolerances. Of the larger cells, Melosira and Coscinodiscus were the only taxa with measurable biomass in the polycultures before rotifer addition; Chaetoceros did not survive in either monoculture or polyculture.
Complementarity and Selection
Complementarity tended to be positive in all cultures through time, with the exception of the Chaetoceros & Nannochloris culture, whereas selection was variable in sign, with no discernible pattern throughout the experiment (S3). Complementarity tended to increase with species richness; experiment-long medians of complementarity for total biovolume were 1.7, 2.5 and 2.7 × 105 µm3/µL for the two-, four- and six-species treatments, respectively. The Chaetoceros & Nannochloris treatment exhibited the lowest complementarity compared to other treatments. The median of experiment-long complementarity (1.6 x104 µm3/µL) was considerably lower than that of other treatments (1.6 to 3.0 × 105 µm3/µL).
Discussion
In this experiment, we used a diverse pool of taxa to quantify the effects of species combination and richness on productivity and stability of phytoplankton communities. There were strong effects of species combination and richness on productivity and stability, but the relative importance of species combination and richness varied for each of the response variables. Species combination was most important to productivity whereas species richness was most important to stability (Table 3). Before rotifer addition, the most productive cultures were those containing Dunaliella (Fig. 4). This finding supports the hypothesis that species-rich communities are productive because they are more likely to contain highly productive species. In contrast, the most species rich communities were the most stable, characterized by low temporal variability in biomass and a recovery after grazer addition. In fact, the polycultures persisted for weeks after rotifer addition (S1). However, the recovery of polycultures was not due to insurance related to grazing tolerances. After rotifer addition, the relative biovolume of the larger cells present in polyculture combinations (Melosira and Coscinodiscus) did not compensate for the loss of the smaller cells after rotifer addition (Fig. S2). Interestingly, monoculture dynamics indicated that Melosira (19–25 µm in diameter) was consumed by rotifers (Fig. 4). These findings are consistent with the ability of rotifers to ingest algae of vastly different size classes and shapes [43], [44], [45]. Although B. plicatilis preferentially consumes small particles with an optimal prey size of 8 µm, the approximate size of Dunaliella, it can consume particles up to 20–25 µm in diameter, the size of individual Melosira cells [46], [47]. Polyculture recovery without compensation by the larger cells therefore points to indirect mechanisms that promoted the temporal stability of polycultures.
We suggest that polyculture recovery was due to reduced rotifer grazing rates in polycultures, resulting in lower rotifer growth rates and a lower net effect of grazing. In particular, the presence of diverse prey in polycultures likely reduced the clearance rates of the rotifers, negatively affecting their growth and allowing polycultures to recover from short-term grazing effects. We define this potential mechanism as the “interference effect” (i.e., different sized algae interfere with the ability of rotifers to adjust their grazing mechanism to one particular prey item). Previous studies have shown that the clearance and growth rate of rotifers, including B. plicatilis, varies with prey size [46], [48], [49], morphological characteristics, such as spines [50] and texture [51], and cell abundance [52]. For B. plicatilis, relative clearance is highest (100%) on prey of about 8 µm in diameter (the size of Dunaliella) and lowest (20%) on prey of 2 µm and 21 µm in diameter (the sizes of Nannochloris and Melosira, respectively). There is support for selective feeding in B. plicatilis [47], [53], which would increase handling time and hence clearance rates in polycultures. In this experiment, rotifers exhibited the highest growth rates in the monocultures of Dunaliella, optimally sized prey, and lower growth rates in the monocultures of Nannochloris and Rhodomonas as well as monocultures of large species, and still lower (negative) growth rates in the six-species polycultures (Fig. 4).
From a predator perspective, it is interesting that resource diversity negatively influenced predator productivity. This finding has been previously documented by Narwani & Mazumder [54], who studied the effects of resource species diversity on the clearance rates of cladoceran zooplankton species. In that study, despite consumer-specific effects of changing resource diversity, resource diversity generally reduced consumer consumption rates. It is important to note that our experimental design employed the use of a single predator species, which has little ecological realism compared to manipulations containing multiple species among trophic levels [55]. For example, with multiple predator species, there may not have been negative effects of phytoplankton diversity on the growth and abundance of rotifers.
The positive complementarity found in this experiment, even before rotifer addition, suggests that either niche partitioning or facilitation may have been important. Although we did not attempt to quantify resource use of the phytoplankton communities, we constructed polycultures from a functionally diverse species pool such that partitioning of light and/or nutrients would be facilitated. Notably, each polyculture consisted of taxa with distinct pigment complements (i.e. chlorophylls a and b in the chlorophytes, fucoxanthins in the diatoms and phycoerythrins in the cryptophyte). In other studies, positive effects of increasing species richness on phytoplankton productivity have been found when species have varied along functional trait axes, including light requirements [23], [56], [57]. In contrast, when species richness has been manipulated using functionally similar algal taxa, no or weak biodiversity effects on productivity have been found [21], [26]. Our work, which demonstrated positive effects of biodiversity on productivity in diverse phytoplankton polycultures, together with these previous studies, highlights the importance of creating functionally diverse phytoplankton communities to study the mechanisms underlying and effects of biodiversity. Moreover, the effects of biodiversity on ecosystem functioning may depend directly on the range of species’ functional traits. Interestingly, biodiversity effects in this study, using evolutionarily distinct phytoplankton with a range of morphological and biochemical traits, were stronger than the effects found in studies of functionally different vascular plants [58], likely with a narrower trait range.
This work has important implications for understanding how biodiversity losses will affect productivity and stability, particularly in aquatic systems. With respect to phytoplankton communities specifically, potential dominance of harmful algal bloom taxa with climate change may create less productive and stable systems. In addition, it has previously been argued that paradigms of biodiversity differ inherently between terrestrial and aquatic systems, but our work refutes this suggestion. We show that phytoplankton polycultures are more productive and stable than monocultures when exposed to a biological perturbation. We further highlight the importance not only of considering functional groups in exploring the relationships between biodiversity, productivity and stability but also in exploring multi-trophic level interactions in biodiversity experiments.
Supporting Information
Photographs showing monocultures and polycultures 24 days after rotifer addition, well after the experiment was terminated. At this point, there was little measurable biomass in the monocultures and most polycultures consisted of communities dominated by rotifers and Dunaliella. These data were not presented in the manuscript because nutrient limitation of phytoplankton growth at this point was certain; however, it is interesting to note that nutrient recycling in the polycultures allowed for sustenance of the dominant organism Dunaliella.
(TIF)
Representative figures showing species dynamics in two- (A), four- (B) and six-species (C) polycultures.
(TIF)
Complementarity and selection data for each treatment (species combination/richness level) and sampling date.
(TXT)
Acknowledgments
We would like to acknowledge Dr. Wayne Van Voorhies for his support during this project and use of laboratory space and equipment. We also thank two anonymous reviewers for comments that improved earlier versions of this manuscript.
Funding Statement
This work was supported by the U.S. Department of Energy under contract DE-EE0003046 awarded to the National Alliance for Advanced Biofuels and Bioproducts and is also a New Mexico Agricultural Experiment Station publication, supported by state funds and the U.S. Hatch Act. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Pimm SL, Russell GJ, Gittleman JL, Brooks TM (1995) The future of biodiversity. Science 269: 347–350. [DOI] [PubMed] [Google Scholar]
- 2. Barnosky AD, Matzke N, Tomiya S, Wogan GOU, Swartz B, et al. (2011) Has the Earth’s sixth mass extinction already arrived? Nature 471: 51–57. [DOI] [PubMed] [Google Scholar]
- 3. Vitousek PM, Mooney HA, Lubchenco J, Melillo JM (1997) Human domination of Earth’s ecosystems. Science 277: 494–499. [Google Scholar]
- 4. Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime JP, et al. (2001) Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 294: 804–808. [DOI] [PubMed] [Google Scholar]
- 5. Chapin III FS, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, et al. (2000) Consequences of changing biodiversity. Nature 405: 234–242. [DOI] [PubMed] [Google Scholar]
- 6. Balvanera P, Pfisterer AB, Buchmann N, He JS, Nakashizuka T, et al. (2006) Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters 9: 1146–1156. [DOI] [PubMed] [Google Scholar]
- 7. Adler PB, Seabloom EW, Borer ET, Hillebrand H, Hautier Y, et al. (2011) Productivity is a poor predictor of plant species richness. Science 333: 1750–1753. [DOI] [PubMed] [Google Scholar]
- 8. Tilman D (1999) The ecological consequences of changes in biodiversity: a search for general principles. Ecology 80: 1455–1475. [Google Scholar]
- 9. Downing AL, Leibold MA (2010) Species richness facilitates ecosystem resilience in aquatic food webs. Freshwater Biology 55: 2123–2137. [Google Scholar]
- 10. Leary DJ, Petchey OL (2009) Testing a biological mechanism of the insurance hypothesis in experimental aquatic communities. Journal of Animal Ecology 78: 1143–1151. [DOI] [PubMed] [Google Scholar]
- 11. McCann KS (2000) The diversity-stability debate. Nature 405: 228–233. [DOI] [PubMed] [Google Scholar]
- 12. van Ruijven J, Berendse F (2005) Diversity-productivity relationships: initial effects, long-term patterns and underlying mechanisms. Proceedings of the National Academy of Sciences 102: 695–700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Griffin JN, Méndez V, Johnson AF, Jenkins SR, Foggo A (2009) Functional diversity predicts overyielding effect of species combination on primary productivity. Oikos 118: 37–44. [Google Scholar]
- 14. Hooper DU, Dukes JS (2004) Overyielding among plant functional groups in a long-term experiment. Ecology Letters 7: 95–105. [Google Scholar]
- 15. Tilman D, Knops J, Wedin D, Reich P, Ritchie M, et al. (1997) The influence of functional diversity and composition on ecosystem processes. Science 277: 1300–1302. [Google Scholar]
- 16. Hooper DU, Vitousek PM (1997) The effects of plant composition and diversity on ecosystem processes. Science 277: 1302–1305. [Google Scholar]
- 17. Naeem S, Li S (1997) Biodiversity enhances ecosystem reliability. Nature 390: 507–509. [Google Scholar]
- 18. Yachi S, Loreau M (1999) Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proceedings of the National Academy of Sciences 96: 1463–1468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Cottingham KL, Brown BL, Lennon JT (2001) Biodiversity may regulate the temporal variability of ecological systems. Ecology Letters 4: 72–85. [Google Scholar]
- 20. Valone TJ, Barber NA (2008) An empirical evaluation of the insurance hypothesis in diversity-stability models. Ecology 89: 522–531. [DOI] [PubMed] [Google Scholar]
- 21. Zhang Q-G, Zhang D-Y (2006) Resource availability and biodiversity effects on the productivity, temporal variability and resistance of experimental algal communities. Oikos 114: 385–396. [Google Scholar]
- 22. Litchman E, Klausmeier CA (2008) Trait-based community ecology of phytoplankton. Annual Review of Ecology and Systematics 39: 615–639. [Google Scholar]
- 23. Behl S, Donval A, Stibor H (2011) The relative importance of species diversity and functional group diversity on carbon uptake in phytoplankton communities. Limnology and Oceanography 56: 683–694. [Google Scholar]
- 24. Striebel M, Behl S, Diehl S, Stibor H (2009) Spectral niche complementarity and carbon dynamics in pelagic ecosystems. The American Naturalist 174: 141–147. [DOI] [PubMed] [Google Scholar]
- 25. Ptacnik R, Solimini AG, Andersen T, Tamminen T, Brettum P, et al. (2008) Diversity predicts stability and resource use efficiency in natural phytoplankton communities. Proceedings of the National Academy of Sciences 105: 5134–5138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Schmidtke A, Gaedke U, Weithoff G (2010) A mechanistic basis for underyielding in phytoplankton communities. Ecology 91: 212–221. [DOI] [PubMed] [Google Scholar]
- 27. Zhang Q-G, Zhang D-Y (2006) Species richness destabilizes ecosystem functioning in experimental aquatic microcosms. Oikos 112: 218–226. [Google Scholar]
- 28. Hamm C, Merkel R, Springer O, Jurkojc P, Maler C, et al. (2003) Architecture and material properties of diatom shells provide effective mechanical protection. Nature 421: 841–843. [DOI] [PubMed] [Google Scholar]
- 29.Hamm C, Smetacek V (2007) Armor: why, when and how. In: Falkowski P, Knoll AK, editors. The Evolution of Primary Producers in the Sea. San Diego: Elsevier.
- 30. Pondaven P, Gallinari M, Chollet S, Bucciarelli E, Sarthou G, et al. (2007) Grazing-induced changes in cell wall silicification in a marine diatom. Protist 158: 21–28. [DOI] [PubMed] [Google Scholar]
- 31.Andersen R, Berges JA, Harrison PJ, Watanabe MM (2005) Recipes for freshwater and seawater media. In: Andersen R, editor. Algal Culturing Techniques. London: Elsevier Academic Press.
- 32. McLachlan J (1964) Some considerations of the growth of marine algae in artificial media. Canadian Journal of Microbiology 10: 769–782. [DOI] [PubMed] [Google Scholar]
- 33. Berges JA, Franklin DJ, Harrison PJ (2001) Evolution of an artificial seawater medium: improvements in enriched seawater, artificial water over the last two decades. Journal of Phycology 37: 1138–1145. [Google Scholar]
- 34.Metcalf JS, Codd G, A. (2009) cyanobacteria, neurotoxins and water resources: Are there implications for human neurodegenerative disease? Amytrophic Lateral Sclerosis: 74–78. [DOI] [PubMed]
- 35. Hillebrand H, Durselen C-D, Kirschtel D, Pollingher U, Zohary T (1999) Biovolume calculation for pelagic and benthic microalgae. Journal of Phycology 35: 403–424. [Google Scholar]
- 36. Loreau M (1998) Separating sampling and other effects in biodiversity experiments. Oikos 82: 600–602. [Google Scholar]
- 37. Pimm SL (1984) The complexity and stability of ecosystems. Nature 307: 321–326. [Google Scholar]
- 38. Conover WJ, Iman RL (1981) Rank transformations between parametric and non-parametric statistics. American Statistician 35: 124–129. [Google Scholar]
- 39.Quinn GP, Keough MJ (2002) Experimental Design and Data Analysis for Biologists. Cambridge: Cambridge University Press. 537 p.
- 40.Sokal RR, Rohlf FJ (1995) Biometry: The Principles and Practices of Statistics in Biological Research New York: W.H. Freeman & Co.
- 41.Bidigare RR, Christensen SJ, Wilde SB, Banack SA (2009) Cyanobacteria and BMAA: Possible linkage with avian vacuolar myelinopathy (AVM) in the south-eastern United States. Amyotrophic Lateral Sclerosis: 71–73. [DOI] [PubMed]
- 42. Loreau M, Hector A (2001) Partitioning selection and complementarity in biodiversity experiments. Nature 412: 72–76. [DOI] [PubMed] [Google Scholar]
- 43. Arndt H (1993) Rotifers as predators on components of the microbial web (bacteria, heterotrophic flagellates, ciliates) - a review. Hydrobiologia 255/256: 231–246. [Google Scholar]
- 44. Starkweather PL (1980) Aspects of the feeding behavior and trophic ecology of susupension-feeding rotifers. Hydrobiologia 73: 63–72. [Google Scholar]
- 45. Gulati RD, Ejsmont-Karabin J, Postema G (1993) Feeding in Euchlanis dilatata lucksiana Hauer on filamentous cyanobacteria and a prochlorophyte. Hydrobiologia 255–256: 269–274. [Google Scholar]
- 46. Hansen B, WernbergMoller T, Wittrup L (1997) Particle grazing efficiency and specific growth efficiency of the rotifer Brachionus plicatilis (Muller). Journal of Experimental Marine Biology and Ecology 215: 217–233. [Google Scholar]
- 47. Pagano M, Saint-Jean L, Arfi R, Bouvy M, Guiral D (1999) Zooplankton food limitation and grazing impact in a eutrophic brackish-water tropical pond (Cote d’Ivoire, West Africa). Hydrobiologia 390: 83–98. [Google Scholar]
- 48. Vadstein O, Øie G, Olsen Y (1993) Particle size dependent feeding by the rotifer Brachionus plicatilis . Hydrobiologia 255–256: 261–267. [Google Scholar]
- 49. Yin XW, Zhao W (2008) Studies on life history characteristics of Brachionus plicatilis O. F. Muller (Rotifera) in relation to temperature, salinity and food algae. Aquatic Ecology 42: 165–176. [Google Scholar]
- 50. Condeporcuna JM, Sarma SSS (1995) Prey selection by Asplancha girodi (Rotifera) - the importance of prey defense mechanisms Freshwater Biology. 33: 341–348. [Google Scholar]
- 51. Mohr S, Adrian R (2000) Functional responses of the rotifers Brachionus calyciflorus and Brachionus rubens feeding on armored and unarmored ciliates. Limnology and Oceanography 45: 1175–1180. [Google Scholar]
- 52. Savas S, Guclu Z (2006) Filtration and ingestion rates of the rotifer Brachionus plicatilis fed five species of microalgae at different cell densities. Israeli Journal of Aquaculture-Bamidgeh 58: 39–45. [Google Scholar]
- 53. Hotos GN (2002) Selectivity of the rotifer Brachionus plicatilis fed mixtures of algal species with various cell volumes and cell densities. Aquaculture Research 33: 949–957. [Google Scholar]
- 54. Narwani A, Mazumder A (2010) Community composition and consumer identity determine the effect of resource species diversity on rates of consumption. Ecology 91: 3441–3447. [DOI] [PubMed] [Google Scholar]
- 55. Douglass JG, Duffy E, Bruno JF (2008) Herbivore and predator diversity interactively affect ecosystem properties in an experimental marine community. Ecology Letters 11: 598–608. [DOI] [PubMed] [Google Scholar]
- 56. Cardinale BJ (2011) Biodiversity improves water quality through niche partitioning. Nature 472: 86–U113. [DOI] [PubMed] [Google Scholar]
- 57. Power LD, Cardinale BJ (2009) Species richness enhances both algal biomass and rates of oxygen production in aquatic microcosms. Oikos 118: 1703–1711. [Google Scholar]
- 58. Schläpfer F, Schmid B (1999) Ecosystem effects of biodiversity: a classification of hypotheses and exploration of empirical results. Ecological Applications 9: 893–912. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Photographs showing monocultures and polycultures 24 days after rotifer addition, well after the experiment was terminated. At this point, there was little measurable biomass in the monocultures and most polycultures consisted of communities dominated by rotifers and Dunaliella. These data were not presented in the manuscript because nutrient limitation of phytoplankton growth at this point was certain; however, it is interesting to note that nutrient recycling in the polycultures allowed for sustenance of the dominant organism Dunaliella.
(TIF)
Representative figures showing species dynamics in two- (A), four- (B) and six-species (C) polycultures.
(TIF)
Complementarity and selection data for each treatment (species combination/richness level) and sampling date.
(TXT)