

Abstract
Muskmelons, both cantaloupe (Cucumis melo Reticulatus Group) and orange-fleshed honey dew (C. melo Inodorus Group), a cross between orange-fleshed cantaloupe and green-fleshed honey dew, are excellent sources of β-carotene. Although β-carotene from melon is an important dietary antioxidant and precursor of vitamin A, its bioaccessibility/bioavailability is unknown. We compared β-carotene concentrations from previously frozen orange-fleshed honey dew and cantaloupe melons grown under the same glasshouse conditions, and from freshly harvested field-grown, orange-fleshed honey dew melon to determine β-carotene bioaccessibility/bioavailability, concentrations of novel β-apocarotenals, and chromoplast structure of orange-fleshed honey dew melon. β-Carotene and β-apocarotenal concentrations were determined by HPLC and/or HPLC-MS, β-carotene bioaccessibility/bioavailability was determined by in vitro digestion and Caco-2 cell uptake, and chromoplast structure was determined by electron microscopy. The average β-carotene concentrations (μg/g dry weight) for the orange-fleshed honey dew and cantaloupe were 242.8 and 176.3 respectively. The average dry weights per gram of wet weight of orange-fleshed honey dew and cantaloupe were 0.094g and 0.071g, respectively. The bioaccessibility of field-grown orange-fleshed honey dew melons was determined to be 3.2±0.3 percent, bioavailability in Caco-2 cells was about 11%, and chromoplast structure from orange-fleshed honey dew melons was globular (as opposed to crystalline) in nature. We detected β-apo-8’-, β-apo-10’, β-apo-12’-, β-apo-14’-carotenals and β-apo-13-carotenone in orange-fleshed melons (at a level of 1-2% of total β-carotene). Orange-fleshed honey dew melon fruit had higher amounts of β-carotene than cantaloupe. The bioaccessibility/bioavailability of β-carotene from orange-fleshed melons was comparable to that from carrot (Dacus carota).
Keywords: carotenoids, β-Carotene, β-apocarotenoids, bioaccessibility, bioavailability, in vitro digestion, caco-2 cells, chromoplasts
INTRODUCTION
Vitamin A deficiency unfortunately affects over 100 million people throughout the world (1). Fruits and vegetables are often utilized as the only treatment for vitamin A deficiencies in human diets as they contain provitamin A carotenoids e.g. β-carotene (2). β-Carotene is the most potent precursor of vitamin A (3). An excellent source of β-carotene is orange-fleshed muskmelons: cantaloupe (Cucmis melo Reticulatus Group) and the novel, little studied orange-fleshed honey dew (Cucumis melo, Inodorus group), a cross between cantaloupe and green-fleshed honey dew (Cucumis melo Inodorus Group) (4). However nothing is known of the bioaccessibility/bioavailability of β-carotene from orange-fleshed melons or of the fruit's chromoplast structure.
β-carotene is lipid soluble and must be incorporated into micelles to be absorbed and bioaccessibility reflects the efficiency of micellerization. Bioavailability represents the amount of the nutrient that is absorbed by the intestinal epithelium and made available for use by the body. β-Carotene bioavailability is affected by chromoplast structure in mature plant tissues, and naturally occurring chromoplasts are globular, tubular, reticulo-tubular, membranous and crystalline types, with globular types providing the best and crystalline types providing the poorest structure for bioavailability of β-carotene (5). Many previous studies have used in vitro digestion and Caco-2 cell uptake to assess the bioaccessibility/bioavailability of β-carotene (6-9). β-Carotene can also undergo asymmetric cleavage to yield β-apocarotenoids that have potential biological roles such as vitamin A activity and transcriptional regulation.
The purpose of this study was to 1) determine β-carotene concentrations in previously frozen orange-fleshed cantaloupe and honey dew melons grown under the same glasshouse conditions, 2) determine the bioaccessibility/bioavailability of β-carotene from aforementioned frozen melon tissues and from fresh, field-grown orange-fleshed honey dew melon tissue; 3) determine the morphological features of chromoplasts in the orange-fleshed melon; and 4) report, newly determined, β-apo-13-carotenone, β-apo-14’-carotenal, β-apo-12’-carotenal, β-apo-10’-carotenal, β-apo-8’-carotenal levels in orange-fleshed melon tissues.
MATERIALS AND METHODS
Chemicals and Supplies
Unless otherwise stated, all chemicals and supplies were purchased from Sigma-Aldrich (St. Louis, MO, USA) or Fisher Scientific (Pittsburgh, PA, USA).
Plant Material
Glasshouse grown orange-fleshed honey dew ‘Orange Dew’ and cantaloupe ‘Cruiser’ plants, 20 each were grown in Weslaco, Texas in well fertilized potting soil as previously described (10). Upon harvest, following natural abscission (indicating fruit maturity), melons were washed in tap water, cut and homogenized according to the protocol of Lester (4). Briefly, the two polar ends of the melons were removed and discarded. The outer epidermis and adjacent hypodermal mesocarp tissues were removed. The remaining edible mesocarp tissue devoid of seed cavity inner meoscarp tissue was homogenized in a Waring® blender. The glasshouse grown melon tissue was stored at -80 °C until shipped on dry ice to Columbus, OH for the study, and used to asses dry weights, β-carotene concentration, and bioaccessibility/bioavailability. Five, fresh, field-grown, orange-fleshed honey dew ‘Uncle Sam’ melons, which is the same cultivar as the ‘Orange Dew’ melon, were received from The Turlock Fruit Company in Turlock, California for identification of the chromoplast structure and to assess bioaccessibility/bioavailability. Upon arrival from California melon tissues were prepared as described above and stored at 4°C until further analysis. We also obtained a locally purchased cantaloupe for the detection of β-apocarotenoids.
Compositional analyses
β-carotene
Melon tissue samples (0.5g) were extracted with 5mL HPLC grade hexane (0.1% BHT), and 200μL of a saturated NaCl solution was added to facilitate phase separation. Samples were vortexed for 60s and centrifuged at 5000 × g for 5min to separate phases. The upper layer was collected and the extraction was repeated two times. The combined hexane layer was dried under a stream of nitrogen and resuspended in 1mL of 2:1 isopropanol/dichloromethane and filtered thru a 0.22 micron syringe filter into an HPLC vial. β-Carotene standard was dissolved in hexane, the absorbance measured at 450nm, and a dilution series was used to generate a standard curve. The β-carotene was detected using a Waters 600 HPLC Pump with a Waters 996 Photodiode Array Detector and a Waters 717plus Autosampler set at 10°C. The separation and quantification was achieved using a Waters YMC C30 reversed phase column (250mm × 4.6 mm) and a two solvent gradient with methanol and methyl t-butyl ether (MTBE) as described in Thakkar et al. (11). Dry weights were determined by adding 1g of melon tissue in triplicate to pre-weighed crucibles, drying in an oven overnight and weighing the air temperature crucible plus contents.
β-Apocarotenoids
Melon samples were analyzed by liquid chromatography mass spectrometry (LC/MS) for β-carotene, β-apo-8’-carotenal, β-apo-10’-carotenal, β-apo-12’-carotenal, β-apo-14’-carotenal, and β-apo-13-carotenone levels. Three 1g samples of fresh, locally purchased, cantaloupe and three 1g samples from a randomly chosen previously frozen ‘OrangeDew’ melon tissue were extracted using a modification of the method of Kane et al. (12). The first fraction containing neutral lipids was collected and analyzed. Briefly, 1 mL of 0.025 M KOH was added to the 1g samples and compounds of interest were extracted three times into 10 mL of hexane. The hexane extracts were combined and dried under N2. The dried extracts were subsequently redissolved in a 1:1 (v/v) mixture of MTBE and methanol and 20 μL was injected onto the HPLC for separation of β-carotene and β-apocarotenoids. Separations were accomplished by reversed phase HPLC using a 4.6 × 150 mm 5 μm YMC C30 column (Waters Corp, Milford, MA). A ternary solvent system consisting of: solvent A, 0.1% formic acid; solvent B, 100% methanol; and solvent C, 100% MTBE was employed. The initial HPLC solvent consisted of 20% A/80% B/0% C and from the time of sample injection through 12 min the solvent was linearly changed to 0% A/30% B/70% C. This was followed by a 3 min re-equilibration period. The flow rate of the solvent was maintained constant at 1.8 mL/min and the column temperature at 35°C. The UV–VIS absorbance of the eluent was monitored using a Waters 996 photodiode array detector.
The HPLC eluate was interfaced with a quadrupole/time-of-flight mass spectrometer (Q Tof Premier, Micromass, UK) via an atmospheric pressure chemical ionization (APcI) probe. The HPLC/MS method used to detect apocarotenoids was used as previously described in Shmarakov et al. (13) with one modification. β-apo-13-Carotenone was measured in APcI positive mode which required a second 20μL injection of the same sample. β-Carotene, β-apo-8’-carotenal, β-apo-10’-carotenal, β-apo-12’-carotenal, and β-apo-14’-carotenal ionized in APcI negative mode and β-apo-13-carotenone ionized in APcI positive mode and were detected as their respective radical ions, at m/z of 536.438, 416.31, 376.28, 350.26, 310.23, and 259.2. The QTof system allowed for quantitative detection with the confidence of accurate mass (typically 1ppm). Mass spectra were acquired in V-mode (~8000 resolution) from 100-1000 m/z with a scan time of 0.5s, peak centroiding, and enhanced duty cycle enabled for the parent m/z. At intervals of 30s, a 0.1s lockspray scan was acquired with leucine enkephalin as the lockspray compound (554.2615 m/z) to correct for minor deviations in calibration due to temperature fluctuations. Prior to analysis, the QTof was fully calibrated from 114 to 1473 m/z using a solution of sodium formate. The resultant MS spectra were acquired and integrated with MassLynx software, V4.1 (Micromass UK, Manchester, UK). Source parameters included: 30 μA corona current, 500 °C probe, 110 °C source block, 35 V cone, 50 L/hr cone gas (N2), 400 L/hr desolvation gas (N2), collision energy 8 eV (non-fragmenting) with argon CID gas (4.2×10-3 mBar). Standard curves were generated using standards of β-carotene (CAS# 7235-40-7), β-apo-8’-carotenal (CAS # 1107-26-2) (Fluka), β-apo-12’-carotenal (CAS # 1638-05-7) (CaroteNature, Lupsingen, Switzerland), β-apo-14’-carotenal(CAS # 6985-27-9) (synthesized for the study), and β-apo-13-carotenone (CAS # 17974-57-1) (synthesized for the study). The purity of β-carotene, β-apo-8’-carotenal, β-apo-12’-carotenal, β-apo-14’-carotenal, and β-apo-13-carotenone by HPLC is 95%, 96%, 99%, 94%, and 98% respectively. β-apo-10’-Carotenal was quantified using the standard curve for β-apo-12’-carotenal. Recoveries for all compounds were determined to be greater than 98%.
Chromoplast Isolation
The protocol for cantaloupe chromoplast enrichment and subsequent preparation for microscopy was provided through personal communication with Wayne W. Fish, South Central Agriculture Research Laboratory, Agricultural Research Service, U.S. Department of Agriculture (14). Fresh orange-fleshed honeydew mesocarp tissue (1000g) and 1000 mL water were mixed in a Waring® blender for 4 min and filtered through parachute cloth. After two centrifugations (30000 × g, 1hr, 4°C) the top layer of the pellet containing the chromoplasts was scraped and suspended in 15.4 mL water with added sodium ascorbate 10:1 (w/w) and 0.02% sodium azide. Two mL of chromoplasts solution were centrifuged to remove buffer and 5 mL 0.05M sodium phosphate, pH 7.2 + 0.3% SDS was added and mixed. Solubilized chromoplasts were fixed in gluteraldehyde. Acetonitrile was added to precipitate the chromoplasts (15) which were washed with 5 mL phosphate buffer. After the third wash, 4 mL of 0.05M sodium phosphate buffer pH 7.2 + 0.3% SDS + 0.02% sodium azide were added and mixed. Chromoplasts were prepared for microscopy and identified according to Vaäsquez-Caicedo et al. (16) by transmission electron microscopy at The Ohio State University Microscopy and Imaging Facility.
In vitro Digestion
In vitro digestion was performed on five fresh and randomly chosen greenhouse grown melons samples, in triplicate to determine the percent digestive stability and the percent micellerization according to the protocol of Thakkar et al. (11), without the “oral phase”. Briefly, this involved a “gastric phase” where the pH of the melon homogenate is adjusted to 2.5±0.1, pepsin is added at 40mg/mL and the mixture is incubated in a shaking water bath at 37°C for 1hr. In the subsequent “intestinal phase” the pH is adjusted to 6.5±0.1, porcine pancreatic lipase, pancreatin, and bile extract is added and the mixture is incubated in a shaking water bath at 37°C for 2hr. The micelle fraction is then isolated from the digesta by centrifugation at 5000 × g for 45 min at 4°C and filtration (0.22 mm pore size) of the collected aqueous (micelle) fraction (11). The aqueous fractions were then applied to Caco-2 cells as described by Chitchumroonchokchai et al. (17).
Caco-2 Cells
Stock cultures of Caco-2 (HTB-39) cells were obtained from American Type Culture Collection and were maintained as previously described (17). The Caco-2 human cell line exhibits characteristics of mature enterocytes (18). T75 flasks of Caco-2 cells were grown 10-14 days post confluency. Following in vitro digestion, the aqueous fractions containing the micelles were collected and each diluted 1:4 with Dulbecco`s minimum essential medium (DMEM) and 12.5 mL of the medium was added to each flask. At the end of 4 hours the medium was collected and cells were washed with ice cold phosphate-buffered saline (PBS) with albumin, which was also collected and combined with the medium. The cells were washed twice with ice cold PBS and the wash was discarded. 10mL of ice cold PBS was then added to each flask, the cells were scraped, collected, and the process repeated. The collected cells in PBS were centrifuged at 2000 × g at 4°C for 5 min and the supernatant was discarded. The cell pellet was resuspended in 2mL PBS and extracted for HPLC analysis. Resuspended cell pellets (100 μL) were used for a protein assay according to Bradford (19). Aliquots of the whole digestion, aqueous (micelle) fraction, fresh 1:4 media, spent media, and the cells were extracted with tetrahydrofuran (THF) and hexane. Briefly, 2mL of THF was added to 2mL of sample and vortexed, then 3mL of hexane was added and vortexed, and centrifuged at 5000 × g for 5min to separate phases. The upper layer was collected and the extraction was repeated two times. Extracts were dried under a stream of nitrogen and redisolved in 2:1 isopropanol/dichloromethane and filtered thru a 0.22 micron syringe filter and injected into the HPLC. HPLC analysis was performed with a Waters 1525μ Binary HPLC Pump with a Waters 996 Photodiode Array Detector and a Waters 717plus Autosampler set at 10°C. A YMC Carotenoid 5μm particle (4.6 × 150mm) Column with a YMC Carotenoid 5μm particle (4.0 × 20mm) Guard Cartridge was used. Separation was achieved by gradient elution with a binary mobile phase of methanol-0.1%(v/v) formic acid (FA) as Solvent A (80:20) and MTBE-methanol-0.1% FA as Solvent B (78:20:2) at a flow rate of 1.8mL/min. Initial conditions were held at 100% A for 1 min then a linear gradient to 40:60 A:B over 5 min, followed by a linear gradient to 100% B over 9 min, a linear gradient back to 100% A for 1 min, and held at 100% A for 4 min for a final chromatographic run time of 20 min. Identification and quantification of the compounds of interest was accomplished by comparison with synthetic standards run in a dilution series before and after the samples.
Statistical Analysis
All data is presented as averages and standard deviations. In Table 1 where comparisons of glasshouse grown orange dew and cantaloupe for dry weight and β-carotene content were made we used a Student's t-test for these pair-wise comparisons.
Table 1.
Dry weight percent and β-carotene concentration of glasshouse grown frozen and field-grown fresh orange-fleshed honey dew and cantaloupe melon edible mesocarp tissues. N=20 for orange dew and cantaloupe and N=5 for orange-fleshed honey dew.
| Orange-fleshed melon type | Tissue preparation | Dry weight % | β-carotene |
|
|---|---|---|---|---|
| μg/g fresh wt. | μg/g dry wt. | |||
| Orange dew | Frozen | 9.4±0.9a* | 22.8±2.4* | 242.3±25.7* |
| Cantaloupe | Frozen | 7.1±0.8 | 12.5±3.8 | 176.3±54.0 |
| Orange-fleshed honey dew | Fresh | 9.8±0.9 | 21.0±2.6 | 213.9±33.5 |
means ± standard deviations are given
Dry weight and β-carotene content of the frozen melons were compared using Student's t-test.
For these parameters orange dew were significantly higher than cantaloupe (P≤0.001).
RESULTS AND DISCUSSION
Orange-fleshed Honey Dew Melon and Cantaloupe: Dry Weights and β-carotene Content
Orange-fleshed honey dew ‘Orange Dew’ had significantly higher dry weight (P ≤0.001) than hybrid orange-fleshed cantaloupe ‘Cruiser’ (Table 1). The difference in dry weight may suggest orange-fleshed honey dew ‘Orange Dew’ melons have a different matrix than the cantaloupe ‘Cruiser’. β-Carotene was the only carotenoid detected at 450 nm by HPLC in either orange-fleshed honey dew or cantaloupe, and greater than 98% was the all-trans isomer (20) (Figure 1.).
Figure 1.
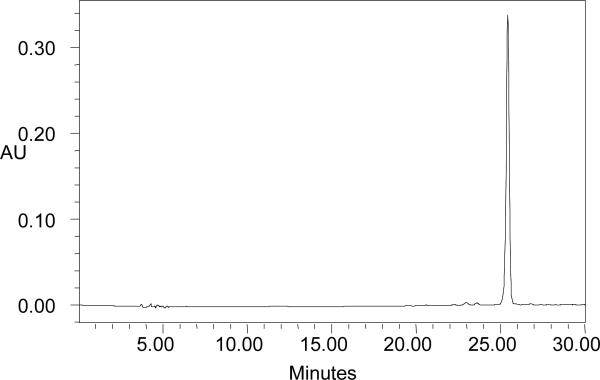
HPLC chromatogram (at 450nm) of lipid extracts of orange-fleshed honey dew and cantaloupe melon tissues. Peak at retention time of 26 min was identified as all-trans-β-carotene by comparison with an authentic standard.
Since dry weights were determined to be significantly different for orange-fleshed honey dew and cantaloupe, β-carotene concentrations per gram of fresh (wet) weight were divided by the dry weight for each sample to correct for fresh weight differences. The mean β-carotene content for orange-fleshed honey dew was significantly greater (P ≤0.001) than cantaloupe (Table 1).
The average dry weight of fresh orange-fleshed melons ‘Uncle Sam’ was similar to dry weights determined for frozen orange-fleshed melon samples (Table 1). The average concentration of all-trans β-carotene of fresh orange-fleshed honey dew melon was similar to that of frozen orange-fleshed honey dew samples (Table 1).
Chromoplast Structure
The chromoplast is a plant compartment in which most of the carotenoids accumulate (21). Chromoplast structure, globular vs. crystalline, has been reported to affect β-carotene bioaccessibility, with a globular structure providing the superior matrix for β-carotene bioaccessibility (5). Chromoplasts were isolated from fresh melon tissue and imaged by TEM as described above. The TEM image in Figure 2 shows that melon chromoplasts were globular in structure as opposed to the crystalline matrix found in carrots (22) (5).
Figure 2.
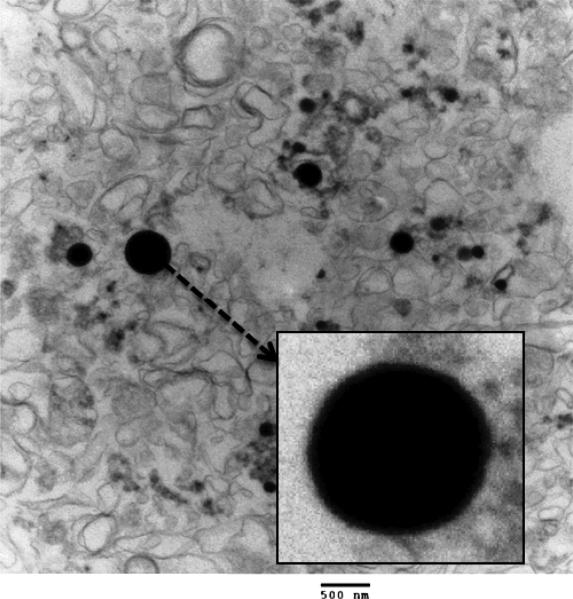
The dark spheres shown in the image are the chromoplasts. Magnification of 18500x. The box shows an enlargement of the large chromoplast.
Bioaccessibility and Bioavailability of β-carotene from Orange-fleshed Melons
Fresh orange-fleshed honey dew melon homogenates were subjected to simulated gastric and intestinal phases of in vitro digestion (Table 2). The percent digestive stability is the percent of β-carotene present in the digesta after simulated digestion. The digesta was separated into the aqueous fraction which contained micelles. The percent micellerization (bioaccessibility) was the percent of β-carotene that was incorporated into the micelles. In vitro digestion was performed in triplicate for each of five fresh melon fruit. The average percent digestive stability was 63.8±3.6% and the average percent micellerization (bioaccessibility) of β-carotene from orange-fleshed edible melon tissue was 3.2±0.3%. Percent micellerization was corrected for loss during digestion, i.e., the percent of digesta that was micellerized (Table 2). These percentages are consistent with results from previous in vitro digestions with glasshouse grown melons where the digestive stability and the percent micellerization (β-carotene bioaccessibility) did not differ between orange-fleshed honey dew and cantaloupe melons (results not shown). The values for digestive stability are consistent with previously published results in maize (Zea mays), cassava (Manihot esculenta), drumstick (Moringa oleiflora) leaves, carrots (Daucus carota), green leafy vegetables, mangoes (Mangifera indica), sweet potatoes (Ipomoea batatas), squash (Cucurbita moschata), and pumpkins (Cucurbita maxima) (11, 23-28). There are many factors that affect the intestinal absorption of carotenoids such as the type and quantity of dietary fat, competition among co-consumed carotenoids, the plant or food matrix (29), and cooking. We tested the effects of adding 2.5-3% vegetable oil to the in vitro digestion of the melons but there was no significant difference in the extent of micellerization. Since melons are typically eaten raw we did not test the effects of cooking. The bioaccessibilities of β-carotene reported for fruits and vegetables are mostly for cooked foods and, thus, the ‘lower’ bioaccessibility of β-carotene in melons may be only relative. Although the bioaccessibility of β-carotene in melons observed in this study is lower than for other fruits and vegetables the reasons appear to be “physiological” in the sense that melon is eaten raw and not cooked and melons have very high water content. In comparison with carrot, the bioaccessibility of β-carotene of 3.2% found in our melon samples are similar to bioaccessibilities found in raw carrots (30) which was confirmed in a separate in vitro digestion assay using locally purchased carrots (results not shown). Carrots in general have 82 μg/g fresh weight β-carotene (31) but are on average 21 % dry wt. (32). Whereas our orange-fleshed honey dew melons samples averaged 22 μg/g fresh weight β-carotene, nearly four-fold less β-carotene than carrot, and averaged 9% dry wt. Bioaccessibility comparison of raw carrots and melon at 3.2% on a dry weight basis yields a total bioaccessibility of 12 μg β-carotene/g dry wt. and 8 μg β-carotene/g dry wt. for carrot and orange-fleshed melons respectively, which are nearly the same. This suggests that the globular chromoplasts of melons appear to provide a more efficient matrix which leads to a higher bioaccessibility of β-carotene than the crystalline chromoplast matrix of carrot (27).
Table 2.
β -carotene concentration from orange-fleshed melon in whole edible tissue, digesta (digestive stability) and aqueous fractions (micellerization or bioaccessibility) following in vitro digestion. (N=5)
| β-carotene |
||||
|---|---|---|---|---|
| Fraction | Stage of Digestion | μg/g fresh wt. | μg/g dry wt. | Percent of whole tissue |
| Whole tissue | Whole food | 20.98±2.67a | 213.9±33.5 | 100 |
| Digesta | Digestive stability | 13.35±1.51 | 135.7±15.5 | 63.8±3.6 |
| Aqueous fraction | Bioaccessibility | 0.43±0.03 | 4.36±0.43 | 3.1±0.3 |
Values are shown as means ± standard deviation
In a separate experiment, Caco-2 cells were used to assess the bioavailability of melon β-carotene. Fresh orange-fleshed honey dew melon underwent in vitro digestion and the diluted micelle fractions were applied to Caco-2 cells. The average uptake of β-carotene by the Caco-2 cells after 4h of incubation (i.e., bioavailability) was 11.6% of the β-carotene applied to the cells (results not shown). This is consistent with the percent uptake of β-carotene in other studies with other fruits and vegetables (11, 33). Results obtained with fresh orange-fleshed melon were consistent with values obtained from frozen, glasshouse grown melons where there was no significant difference between β-carotene of orange-fleshed honey dew and cantaloupe melons for percent digestive stability, micellerization, and Caco-2 cell bioavailability (results not shown).
β-apocarotenoids in Melons
We measured the levels of β-apo-13-carotenone, β-apo-14’-carotenal, β-apo-12’-carotenal, β-apo-10’-carotenal, β-apo-8’-carotenal, and β-carotene of a locally purchased cantaloupe and a greenhouse grown OrangeDew (Figure 3). The β-apocarotenoids in total are present at approximately 1.5% of the level of β-carotene. Figure 4 shows the relative concentration of the β-apocarotenoids. The β-apocarotenoids profiles of the two different types of melons are approximately the same. β-apocarotenoids are thus present in foods at low levels and may also be formed enzymatically in mammals (17) where they may have biological activities mediated in part by their interaction with retinoid receptors (34). We are currently assessing the β-apocarotenoid levels of other β-carotene rich fruits and vegetables with an improved LC-MS method, in addition to assessing the bioaccessibility/bioavailability. β-apocarotenoids may be more bioavailable than β-carotene due to chain length and the addition of oxygen.
Figure 3.
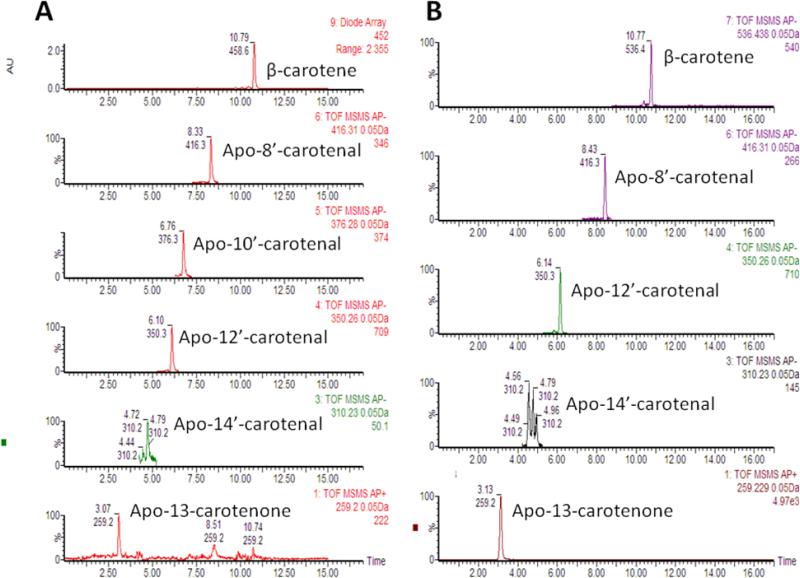
β-Apocarotenoids and β-carotene in the ‘OrangeDew’ melons A and standards B. In panel A the top chromatogram shows the absorbance at 452nm and the other chromatograms show the apocarotenoids at their respective masses. β-apo-10’-carotenal standard was not available and was quantified using the standard curve for β-apo-12’-carotenal.
Figure 4.
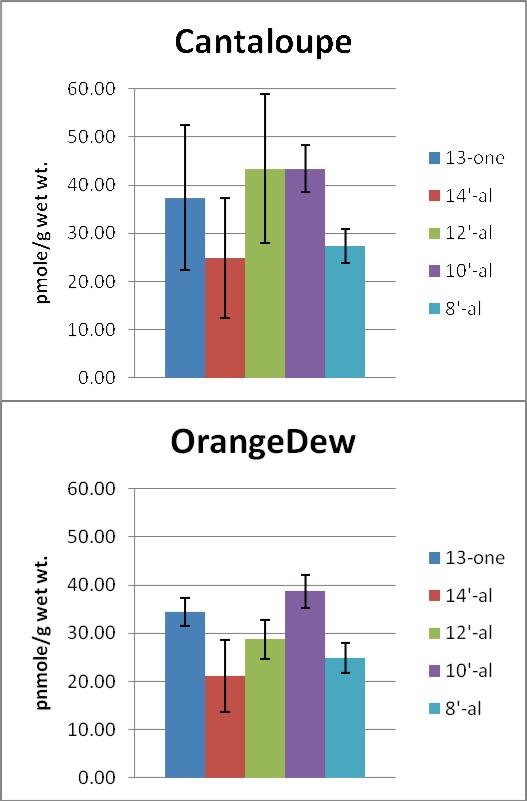
Average β-apocarotenoid levels in two different orange-fleshed melons. Values are from triplicate 1g extractions of each melon with error bars showing SD.
The orange-fleshed honeydew melons are known to be safer for the consumer because they lack rough outer netting, unlike cantaloupe fruit which harbors enteric bacteria, and they are sweeter and store longer than the typical cantaloupe melon (35). In our study orange-fleshed honey dew had significantly higher dry weight and β-carotene concentrations than cantaloupe but each cultivar had similar β-carotene bioaccessibilities. Thus, both appear to be comparable sources of dietary provitamin A for humans and on par with carrots, another major source of provitamin A.
Acknowledgements
We thank Ms. Vanessa Reed for expert technical assistance, Dr. Wayne Fish for sharing the details of the isolation procedure for chromoplasts, and Dr. Mark Failla for assistance with in vitro digestions and Caco-2 cells. This work was supported by grants from the National Institutes of Health to EHH (R01-DK044498 and R01-HL049879) and by the USDA.
Abbreviations Used
- MTBE
methyl-t-butyl-ether
- TEM
transmission electron microscopy
- HPLC
high performance liquid chromatography
- PBS
phosphate-buffered saline
- SDS
sodium dodecyl sulfate
- FA
fatty acid
- THF
tetrahydrofuran
- DMEM
Dulbecco's modified Eagle's medium
- BHT
butylated hydroxytoluene
- LC/MS
liquid chromatography/mass spectrometry
- APcI
atmospheric pressure chemical ionization
Literature Cited
- 1.During A, Harrison EH. Mechanisms of provitamin A (carotenoid) and vitamin A (retinol) transport into and out of intestinal Caco-2 cells. J. Lipid Res. 2007;48:2283–2294. doi: 10.1194/jlr.M700263-JLR200. [DOI] [PubMed] [Google Scholar]
- 2.Harrison EH. Mechanisms of digestion and absorption of dietary vitamin A. Annu. Rev. Nutr. 2005;25:87–103. doi: 10.1146/annurev.nutr.25.050304.092614. [DOI] [PubMed] [Google Scholar]
- 3.Botella-Pavia P, Rodriguez-Conception M. Carotenoid biotechnology in plants for nutritionally improved foods. Physiol. Plant. 2006;126:369–381. [Google Scholar]
- 4.Lester GE. Antioxidant, sugar, mineral, and phytonutrient concentrations across edible fruit tissues of orange-fleshed honeydew melon (Cucumis melo L.). J. Agric. Food Chem. 2008;56:3694–3698. doi: 10.1021/jf8001735. [DOI] [PubMed] [Google Scholar]
- 5.Sitte P, Falk H, Liedvogel B. Chromoplast. In: Czygan FC, editor. Pigments in plants. 2nd ed Gustav Fisher Verlag; Stuttgart, Germany: 1980. pp. 1117–1148. [Google Scholar]
- 6.Failla ML, Huo T, Thakkar SK. In vitro screening of relative bioaccessibility of carotenoids from foods. Asia Pac. J. Clin. Nutr. 2008;17(Suppl 1):200–203. [PubMed] [Google Scholar]
- 7.Garrett DA, Failla ML, Sarama RJ. Development of an in vitro digestion method to assess carotenoid bioavailability from meals. J. Agric. Food Chem. 1999;47:4301–4309. doi: 10.1021/jf9903298. [DOI] [PubMed] [Google Scholar]
- 8.Garrett DA, Failla ML, Sarama RJ, Craft N. Accumulation and retention of micellar beta-carotene and lutein by Caco-2 human intestinal cells. J. Nutr. Biochem. 1999;10:573–581. doi: 10.1016/s0955-2863(99)00044-3. [DOI] [PubMed] [Google Scholar]
- 9.Failla ML, Thakkar SK, Kim JY. In vitro bioaccessibility of beta-carotene in orange fleshed sweet potato (Ipomoea batatas, Lam.). J. Agric. Food Chem. 2009;57:10922–10927. doi: 10.1021/jf900415g. [DOI] [PubMed] [Google Scholar]
- 10.Lester GE. Supplemental Foliar Potassium Applications durin Muskmelon Fruit Development Can Improve Fruit Quality, Ascorbic Acid, and Beta-carotene Contents. J. AMER. Soc. Hort. Sci. 2005;130:649–653. [Google Scholar]
- 11.Thakkar SK, Maziya-Dixon B, Dixon AG, Failla ML. Beta-carotene micellarization during in vitro digestion and uptake by Caco-2 cells is directly proportional to beta-carotene content in different genotypes of cassava. J. Nutr. 2007;137:2229–2233. doi: 10.1093/jn/137.10.2229. [DOI] [PubMed] [Google Scholar]
- 12.Kane MA, Chen N, Sparks S, Napoli JL. Quantification of endogenous retinoic acid in limited biological samples by LC/MS/MS. Biochem. J. 2005;388:363–369. doi: 10.1042/BJ20041867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Shmarakov I, Fleshman MK, D'Ambrosio DN, Piantedosi R, Riedl KM, Schwartz SJ, Curley RW, Jr, von Lintig J, Rubin LP, Harrison EH, Blaner WS. Hepatic stellate cells are an important cellular site for beta-carotene conversion to retinoid. Arch. Biochem. Biophys. 2010 doi: 10.1016/j.abb.2010.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fish WW. Interaction of sodium dodecyl sulfate with watermelon chromoplasts and examination of the organization of lycopene within the chromoplasts. J. Agric. Food Chem. 2006;54:8294–8300. doi: 10.1021/jf061468+. [DOI] [PubMed] [Google Scholar]
- 15.Fish WW. Novel procedure for the extraction and concentration of carotenoid-containing chromoplasts from selected plant systems. J. Agric. Food Chem. 2007;55:1486–1490. doi: 10.1021/jf0626213. [DOI] [PubMed] [Google Scholar]
- 16.Vasquez-Caicedo AL, Heller A, Neidhart S, Carle R. Chromoplast morphology and beta-carotene accumulation during postharvest ripening of Mango Cv. ‘Tommy Atkins’. J. Agric. Food Chem. 2006;54:5769–5776. doi: 10.1021/jf060747u. [DOI] [PubMed] [Google Scholar]
- 17.Chitchumroonchokchai C, Schwartz SJ, Failla ML. Assessment of lutein bioavailability from meals and a supplement using simulated digestion and caco-2 human intestinal cells. J. Nutr. 2004;134:2280–2286. doi: 10.1093/jn/134.9.2280. [DOI] [PubMed] [Google Scholar]
- 18.Ellwood KC, Chatzidakis C, Failla ML. Fructose utilization by the human intestinal epithelial cell line, Caco-2. Proc. Soc. Exp. Biol. Med. 1993;202:440–446. doi: 10.3181/00379727-202-43556. [DOI] [PubMed] [Google Scholar]
- 19.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 20.Bohm V, Puspitasari-Nienaber NL, Ferruzzi MG, Schwartz SJ. Trolox equivalent antioxidant capacity of different geometrical isomers of alpha-carotene, beta-carotene, lycopene, and zeaxanthin. J. Agric. Food Chem. 2002;50:221–226. doi: 10.1021/jf010888q. [DOI] [PubMed] [Google Scholar]
- 21.Vishnevetsky M, Ovadis M, Itzhaki H, Levy M, Libal-Weksler Y, Adam Z, Vainstein A. Molecular cloning of a carotenoid-associated protein from Cucumis sativus corollas: homologous genes involved in carotenoid sequestration in chromoplasts. Plant J. 1996;10:1111–1118. doi: 10.1046/j.1365-313x.1996.10061111.x. [DOI] [PubMed] [Google Scholar]
- 22.Zhou JR, Gugger ET, Erdman JW., Jr The crystalline form of carotenes and the food matrix in carrot root decrease the relative bioavailability of beta- and alpha-carotene in the ferret model. J. Am. Coll. Nutr. 1996;15:84–91. doi: 10.1080/07315724.1996.10718569. [DOI] [PubMed] [Google Scholar]
- 23.Kean EG, Hamaker BR, Ferruzzi MG. Carotenoid bioaccessibility from whole grain and degermed maize meal products. J. Agric. Food Chem. 2008;56:9918–9926. doi: 10.1021/jf8018613. [DOI] [PubMed] [Google Scholar]
- 24.Thakkar SK, Failla ML. Bioaccessibility of Pro-Vitamin A Carotenoids Is Minimally Affected by Non Pro-Vitamin A Xanthophylls in Maize (Zea mays sp.). J. Agric. Food Chem. 2008 doi: 10.1021/jf802430u. [DOI] [PubMed] [Google Scholar]
- 25.Pullakhandam R, Failla ML. Micellarization and intestinal cell uptake of beta-carotene and lutein from drumstick (Moringa oleifera) leaves. J. Med. Food. 2007;10:252–257. doi: 10.1089/jmf.2006.250. [DOI] [PubMed] [Google Scholar]
- 26.Hedren E, Diaz V, Svanberg U. Estimation of carotenoid accessibility from carrots determined by an in vitro digestion method. Eur. J. Clin. Nutr. 2002;56:425–430. doi: 10.1038/sj.ejcn.1601329. [DOI] [PubMed] [Google Scholar]
- 27.Mulokozi G, Hedren E, Svanberg U. In vitro accessibility and intake of beta-carotene from cooked green leafy vegetables and their estimated contribution to vitamin A requirements. Plant Foods Hum. Nutr. 2004;59:1–9. doi: 10.1007/s11130-004-4305-7. [DOI] [PubMed] [Google Scholar]
- 28.Priyadarshani AM, Chandrika UG. Content and in-vitro accessibility of pro-vitamin A carotenoids from Sri Lankan cooked non-leafy vegetables and their estimated contribution to vitamin A requirement. Int. J. Food Sci. Nutr. 2007;58:659–667. doi: 10.1080/09637480701395580. [DOI] [PubMed] [Google Scholar]
- 29.Lemke SL, Dueker SR, Follett JR, Lin Y, Carkeet C, Buchholz BA, Vogel JS, Clifford AJ. Absorption and retinol equivalence of beta-carotene in humans is influenced by dietary vitamin A intake. J. Lipid Res. 2003;44:1591–1600. doi: 10.1194/jlr.M300116-JLR200. [DOI] [PubMed] [Google Scholar]
- 30.Hedren E, Mulokozi G, Svanberg U. In vitro accessibility of carotenes from green leafy vegetables cooked with sunflower oil or red palm oil. Int. J. Food Sci. Nutr. 2002;53:445–453. doi: 10.1080/09637480220164334. [DOI] [PubMed] [Google Scholar]
- 31.U.S. Department of Agriculture . Agricultural Research Service USDA National Nutrient Database for Standard Reference, Release 22. Nutrient Data Laboratory; 2010. [Google Scholar]
- 32.Lester GE, Baker LR, Kelly JF. Physiology of sugar accumulation in carrot breeding lines and cultivars. J. AMER. Soc. Hort. Sci. 1982;107:381–387. [Google Scholar]
- 33.De Jesus Ornelas-Paz J, Failla ML, Yahia EM, Gardea-Bejar A. Impact of the stage of ripening and dietary fat on in vitro bioaccessibility of beta-carotene in ‘Ataulfo’ mango. J. Agric. Food Chem. 2008;56:1511–1516. doi: 10.1021/jf072751r. [DOI] [PubMed] [Google Scholar]
- 34.Eroglu A, Hruszkewycz DP, Curley RW, Jr, Harrison EH. The eccentric cleavage product of beta-carotene, beta -apo-13-carotenone, functions as an antagonist of RXRalpha. Arch. Biochem. Biophys. 2010 doi: 10.1016/j.abb.2010.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Flores A. Orange-Fleshed, Organic Honeydews! Agricultural Research Magazine. Vol. 2007;55:17. [Google Scholar]