

Abstract
Parasites may have large effects on host population dynamics, marine fisheries and conservation, but a clear elucidation of their impact is limited by a lack of ecosystem-scale experimental data. We conducted a meta-analysis of replicated manipulative field experiments concerning the influence of parasitism by crustaceans on the marine survival of Atlantic salmon (Salmo salar L.). The data include 24 trials in which tagged smolts (totalling 283 347 fish; 1996–2008) were released as paired control and parasiticide-treated groups into 10 areas of Ireland and Norway. All experimental fish were infection-free when released into freshwater, and a proportion of each group was recovered as adult recruits returning to coastal waters 1 or more years later. Treatment had a significant positive effect on survival to recruitment, with an overall effect size (odds ratio) of 1.29 that corresponds to an estimated loss of 39 per cent (95% CI: 18–55%) of adult salmon recruitment. The parasitic crustaceans were probably acquired during early marine migration in areas that host large aquaculture populations of domesticated salmon, which elevate local abundances of ectoparasitic copepods—particularly Lepeophtheirus salmonis. These results provide experimental evidence from a large marine ecosystem that parasites can have large impacts on fish recruitment, fisheries and conservation.
Keywords: recruitment, salmon, parasites, conservation, fisheries
1. Introduction
Infectious diseases may threaten biodiversity conservation and food security [1,2]. Conceptually, a number of disease-associated mechanisms may elevate extinction risk, which include small pre-epidemic population size and the presence of a reservoir host population [3]. Furthermore, evolutionary similarity to domesticated animals may be a key factor associated with parasite-mediated population declines of wildlife [4]. For food security, disease threats to domesticated populations are associated with increased host density or emergence of novel pathogen strains [1,5]. More broadly, parasites are increasingly being recognized for their direct and indirect effects on host population dynamics and community structure [6–10].
However, clear experimental demonstrations of parasites regulating or limiting host population dynamics in field conditions are rare [11,12]. For marine fishes, uncertainty may be particularly high because fish recruitment is highly stochastic and nonlinear [13,14], and analyses of recruitment are correlative with correlates often being ephemeral [15]. Nevertheless, there is a growing perception of the potential regulatory importance of parasites in marine fish populations [16–19]. Furthermore, parasites that are shared between wild marine fish populations and evolutionarily similar domesticated stocks in aquaculture may affect fish population dynamics, fisheries productivity, biodiversity conservation and aquaculture productivity [5,20,21].
In coastal seas, domesticated populations in aquaculture are rapidly growing and these experience both incidental and persistent disease outbreaks [22–24]. Although disease and its control are costly for aquaculture producers, concerns also have emerged on the potential wider consequences for marine ecosystems and wild fisheries [20,21,25,26]. Perhaps nowhere is this more contentious than with Pacific (Oncorhynchus spp.) and Atlantic salmon (Salmo salar L.) and their native, specialist ectoparasitic copepod, the salmon louse Lepeophtheirus salmonis (Krøyer) [27–29]. In the North Atlantic, stocks of wild S. salar have shown a marked and steady decline of approximately 45 per cent since the 1980s, with variability among stocks [30], and correlations to salmon aquaculture have been drawn [31,32].
Lepeophtheirus salmonis is a marine ectoparasitic copepod of salmonids (Salmo spp., Salvelinus spp., Oncorhynchus spp.) that feeds on host surface tissues causing morbidity and mortality [27,28]. There is no intermediate host and the parasitic gravid female releases free-living larvae to the water column where they can be dispersed among wild and farmed populations [5,33]. Lepeophtheirus salmonis is extremely unusual amongst macroparasites [34,35] in typically reaching 100 per cent prevalence on wild one sea-winter (1SW) and two sea-winter (2SW) Atlantic salmon [36,37]. Persistent re-infestation of wild salmon at sea is apparent from the significantly higher mean abundance of L. salmonis on wild 2SW fish compared with 1SW adults [37]. Thus, L. salmonis must be viewed either as being exceptionally well adapted in locating the host fish or, more likely, the behaviour of salmon is such that they inevitably encounter the infective copepodid stage of L. salmonis both in coastal waters and in the open Atlantic ocean.
Numerous studies have associated salmon farming and sea louse infestations of wild juvenile salmon, but the impact on salmon recruitment remains contentious [20,38–44]. A primary reason is that many studies are correlative and results depend on modelling assumptions [32,38,39]. By contrast, manipulative field experiments provide an opportunity for strong inferences based on differential survival between paired releases of control and parasiticide-treated groups of emigrant juvenile salmon smolts [40,42,45,46]. The parasiticides affect crustaceans, which include other native salmon parasites, particularly Caligus elongatus, though L. salmonis remains the focus owing to its prevalence and adverse lethal and sub-lethal effects [27,28]. Some of the field experiments have identified significant impacts on salmon survival [42,45], whereas others have claimed no significant effect at the population level [40,46].
We assembled and analysed all the published results of large-scale, pairwise field experiments of the marine survival of Atlantic salmon in the Northeast Atlantic Ocean. These studies were conducted in Ireland and Norway, and typically occurred in river systems discharging to near shore marine waters that accommodated salmon farms [40,42,45–47]. Figure 1 shows the location of coastal salmon farm sites adjacent to the named experimental release rivers in Norway and Ireland; however, not all farm sites were in active production in all years of experimental releases. The published studies all were substantial, involving the release of thousands of hatchery-raised juvenile salmon smolts into rivers. Prior to release, all fish were tagged (either coded wire or carlin tags) and approximately half the juveniles received a parasiticide treatment, while the others were untreated controls. Targeted tag recovery programmes and screening of commercial catches of return adult spawners allowed comparisons of marine survival between treatment and control groups. The large majority of the recovered fish spent 1 year at sea, and we focus on these so-called 1SW fish in the main analysis. Further details, including comparative analyses of return adults of all sea ages (1SW, 2SW and 3SW), and the data analysed, are presented in the electronic supplementary material.
Figure 1.

Map of the study regions showing rivers where salmon smolts were released and salmon farm locations (black circles). Interpretation of farming activities over the course of the study period should be made with caution, because the distributions of farms, stocking levels, parasite levels and management approaches have varied over the years.
2. Material and methods
(a). Data sources
The data spanned 24 trials between 1996 and 2008, and included a total of 283 347 smolts released into 10 rivers mostly in Ireland but also Norway (figure 1 and table 1; electronic supplementary material). Most trials involved a single release of paired control–treatment groups into a river (or estuary [47]) during the spring emigration (typically April–May). In some trials, there were multiple separate releases within an emigration season: for these, we summed the data across the multiple releases into a single observation for that annual trial. In 21 of 24 trials, the in-feed treatment emamectin benzoate (Slice; Schering-Plough) was applied; this is a widely used parasiticide in salmon aquaculture [48]. For three trials, a different proprietary compound, Substance Ex (Alpharma), was used [47] (table 1), which involves a topical bath treatment.
Table 1.
Summary of the data, giving the year, country and river of release as well as the numbers of smolts released (N) and of one sea-winter (1SW) adults recovered (R) for treatment groups receiving parasiticides (t) and controls (c). (Also shown are the total number of smolts released (Ntotal) and sources of the data (ref.), those studies with multiple within-season releases that were grouped (*), and the studies that used the proprietary Substance Ex as treatment (¶) rather than emamectin benzoate. Note that numbers of recaptured salmon for references [40] and [46] are their survival estimates that incorporated raising factors applied to the raw data (tags recovered), and may therefore be inflated relative to the other data.)
| year | country | river | Nt | Rt | Nc | Rc | Ntotal | ref. |
|---|---|---|---|---|---|---|---|---|
| 1996 | Norway | Agdenes | 3000 | 53 | 2985 | 43 | 5985 | [47]¶ |
| 1997 | Norway | Agdenes | 2935 | 16 | 2936 | 12 | 5871 | [47]¶ |
| 1998 | Norway | Agdenes | 2966 | 37 | 2977 | 14 | 5943 | [47]¶ |
| 2002 | Norway | Dale | 5086 | 56 | 4859 | 40 | 9945 | [45]* |
| 2004 | Ireland | Erriff | 4325 | 44 | 4229 | 34 | 8554 | [42] |
| 2005 | Ireland | Erriff | 4659 | 37 | 4689 | 2 | 9348 | [42] |
| 2003 | Ireland | Invermore | 4589 | 17 | 4594 | 9 | 9183 | [42] |
| 2004 | Ireland | Invermore | 4653 | 37 | 4671 | 26 | 9324 | [42] |
| 2005 | Ireland | Invermore | 4716 | 31 | 4750 | 17 | 9466 | [42] |
| 2003 | Ireland | Owengowla | 4955 | 35 | 4822 | 3 | 9777 | [42] |
| 2004 | Ireland | Owengowla | 4655 | 51 | 4699 | 22 | 9354 | [42] |
| 2005 | Ireland | Owengowla | 4583 | 54 | 4735 | 53 | 9318 | [42] |
| 2001 | Ireland | Burrishoole | 5496 | 565 | 10039 | 992 | 15535 | [40] |
| 2002 | Ireland | Burrishoole | 5960 | 544 | 5989 | 545 | 11949 | [40] |
| 2003 | Ireland | Burrishoole | 4755 | 472 | 4587 | 374 | 9342 | [40] |
| 2004 | Ireland | Burrishoole | 4437 | 402 | 4369 | 398 | 8806 | [40] |
| 2005 | Ireland | Burrishoole | 3793 | 253 | 3867 | 182 | 7660 | [40] |
| 2006 | Ireland | Burrishoole | 8716 | 508 | 12779 | 549 | 21495 | [40]* |
| 2007 | Ireland | Burrishoole | 6746 | 492 | 6795 | 435 | 13541 | [40] |
| 2008 | Ireland | Burrishoole | 10132 | 163 | 10224 | 125 | 20356 | [40]* |
| 2001 | Ireland | Bundorragha | 12787 | 2056 | 12753 | 1873 | 25540 | [46]* |
| 2006 | Ireland | Erne | 5752 | 70 | 10357 | 68 | 16109 | [46] |
| 2006 | Ireland | Lee | 5207 | 10 | 5131 | 10 | 10338 | [46] |
| 2006 | Ireland | Screebe | 10990 | 157 | 9618 | 121 | 20608 | [46] |
(b). Analytical approaches
We considered three independent but complementary analyses based on odds ratios, paired sample t-tests, and mixed-effects models, each of which synthesize the data across studies to quantitatively assess sea louse-induced mortality of Atlantic salmon. The first approach is a standard survival analysis that leads to estimates of the odds ratio in survival between control–treatment data pairs, as well as an overall meta-analytic mean odds ratio [49]. The second analysis begins with the standard assumptions of survival analysis, and leads to a simple paired sample t-test of survival estimates on a natural log scale. A key advantage of this approach is that it permits an estimation of the per cent of adult salmon recruitment that is lost to parasites. The third analytical approach also begins with the standard assumptions of survival analysis but, relative to the second approach, differs subtly in how residual variation is distributed between control and treatment groups. This third approach leads to a mixed-effects model of survival estimates, again on a log scale, and also permits estimation of the per cent of salmon recruitment lost to parasites.
Our initial standard meta-analysis is an approach that is often adopted when seeking to estimate the overall effect associated with a range of trials that report the same outcome variable. Because the outcomes were dichotomous, an appropriate measure of association was the odds ratio. Given the degree of heterogeneity evident among the studies, a random effects meta-analysis [49] was adopted. The meta.DSL function from the rmeta package of the R statistical software was used to carry out the analysis.
The second and third approaches begin with a standard survival analysis where the probability of survival from time t = 0 to time t = T is
| 2.1 |
and μ is known variously as the instantaneous mortality rate, hazard rate or force of mortality [50].
The second analytical approach involves a model of the data according to
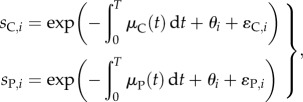 |
2.2 |
where sC,i and sP,i are the proportions of fish that were recovered from the control group (C) and parasiticide-treatment group (P) in trial i. The parameters μC and μP are the instantaneous mortality rates of Atlantic salmon from control (C) and parasiticide-treatment (P) groups. The mortality rate then is integrated over the duration of the study from the time of release (t = 0) to the average time of recapture (t = T). Variation in mortality rates among trials (but not between control and treatment groups within trials) is modelled as the normally distributed random variable, θi with a mean of zero; thus, the treatment and control groups in the same release share the same environmental variability in mortality rate. The parameter θi also encompasses random variation in T among trials, but we assume that T does not vary between control and treatment groups within trials. The parameter ɛC,i is a normally distributed random variable with a mean of zero that models random variation among trials in parasite exposure in control groups. The parameter ɛP,i is a normally distributed random variable with a mean of zero that models random variation among trials in the level of protection conferred by parasiticide treatment.
Equations (2.2) can be simplified to
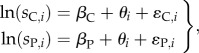 |
2.3 |
where
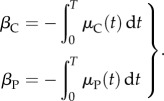 |
2.4 |
Equations (2.4) lead to an estimate of the average mortality owing to parasites
| 2.5 |
where φ = βP − βC. It follows that φ can therefore be estimated as
| 2.6 |
where ɛi = ɛP,i − ɛC,i is a random normal variable with mean of zero and variance to be estimated. Equation (2.6) indicates that statistical inference on φ, and therefore parasite-induced mortality of Atlantic salmon, can be conducted as a simple paired sample t-test between paired survival estimates of control and treatment groups among trials on a natural log scale.
The third model of the data we consider is
| 2.7 |
where j represents control (C) or parasiticide-treatment (P) groups. Equation (2.7) is similar to equations (2.2), except that ɛi,j is a normally distributed random variable that is distributed over control and treatment groups across the dataset. Here, it is assumed that mortality rates for each individual group—be it control or treatment—vary among each other according to a single random variable, whereas in equations (2.2) variation in mortality rate among trials within treatment or control groups was modelled separately for control and parasiticide-treatment groups.
This third approach leads to a model for the log-transformed survival data
| 2.8 |
which has the form of a mixed-effects model, where θi is a random effect for shared environmental variability for control and treatment groups within trials and ɛi,j is the residual variation. Here, statistical inference on whether parasiticide treatment has a significant effect on survival can be conducted by means of a likelihood ratio test between equation (2.8) and its null version where βj is constant (βj = β0). Estimation of φ, and therefore parasite-induced mortality (M) as defined above, can be conducted from the maximum-likelihood estimate of the difference (and associated s.e.) between the treatment and control fixed effects in the mixed model (equation (2.8)).
3. Results
Meta-analysis of differential survival between control and parasiticide-treatment groups indicated a consistent directionality of the effect of treatment among trials, albeit with variation among trials in effect size, and a significant protective effect of treatment (overall odds ratio 1.29 with 95% CI: 1.18–1.42; figure 2). The paired sample t-test revealed significant differences in survival between treatment and control groups (t23 = 3.37, p-value = 0.003). Similarly, for our third (mixed model) analytical approach, treatment had a significant effect on survival (likelihood ratio test; χ21 = 9.64, p = 0.002). The estimated difference in log survival between paired treatment and control groups was φ = 0.50 from both the latter analyses. Uncertainty in φ was given by 95% CIs of 0.19–0.80 in the paired sample t-test, and a range of two standard errors giving the interval 0.20–0.79 in the mixed model. These estimates of φ correspond to parasite-associated mortality of 1SW Atlantic salmon of M = 1 − exp[−φ] = 39.3 per cent (95% CI from paired sample t-test, 17.5–55.3%; range of two standard errors from mixed model, 18.3–54.8%). The complementary analysis (see the electronic supplementary material) that included all sea ages (1–3SW) of returning adults gave a final estimate of M = 39.9 per cent.
Figure 2.

Forest plot displaying a random effect meta-analysis of the effect of treatment on the likelihood of a one sea-winter (1SW) adult salmon returning. Horizontal lines represent the 95% CIs of the effect size in each trial, and the relative sizes of solid squares reflect the percentage weighting (based on s.e. of effect sizes) of each trial in the meta-analysis (range 0.42–8.12%). The open diamond shows the overall meta-analytic effect across all studies, with its width corresponding to the 95% CI. Results are given by trial, identified by location (country and river of release of smolts), year (year when smolts were released) and ref. (the reference where the data were published).
4. Discussion
These results provide manipulative experimental evidence at a large spatial scale from the marine environment that parasitism is a significant limiting factor for the recruitment of Atlantic salmon. Treatment of smolts with parasiticide significantly improved their survival to adulthood relative to control untreated groups. The most common treatment in the trials, emamectin benzoate, has a half-elimination period of 9–12 days in Atlantic salmon smolts [51] and is effective for up to the first 1 to 2 months at sea [48]. The differential survival between control and treatment groups is therefore attributable to near shore and coastal waters where the juvenile post-smolts transit from rivers to offshore waters. These near shore waters typically host large domesticated salmon populations in aquaculture operations (figure 1), which represent a large—but not exclusive—source of sea lice. Potential wild sources of sea lice include sea trout (Salmo trutta L.), Arctic char (Salvelinus alpinus L.) and spring-returning adult S. salar [27].
Although we focus attention on L. salmonis, in view of its prevalence and known adverse effects on host salmon, it is likely that C. elongatus, which also is native and occurs on both wild and farmed salmon [27], contributes to mortality. In addition, it is not implausible that there may be other unknown effects of the treatments that improve survival. Another caveat is that the data are based on hatchery-reared smolts, which, among other differences with wild smolts, are larger: hatchery smolts are typically 18–19 cm, whereas wild smolts are typically 11–14 cm in Ireland and Norway. The larger size of hatchery smolts can partially offset their typically low marine survivorship compared with wild smolts [52]. Nonetheless, owing to the size-related effects of sea-louse-induced stress on smolts [27,28], wild salmon would probably experience higher mortality than we have estimated here for hatchery smolts.
We considered the possibility of a dose-dependent effect—i.e. including sea louse abundance on farms as a covariate—but proceeded otherwise. It was not possible to formally structure the analysis in this way because we lack sufficient farm data for all areas and years. Furthermore, local hydrodynamic processes that disperse the infective sea louse larval stage will be highly variable among locations [33,53], making it difficult to construct a covariate that is geographically comparable. Rather, we regard the variance in the data of differential survival as representative of the spatio-temporal variation of ectoparasite exposure that wild salmon experience in salmon-farming regions of the Northeast Atlantic.
While it is apparent that parasites have potential to be a significant source of mortality in wild fish populations [10,16,54], it is difficult to measure marine mortality associated with disease, and furthermore to evaluate whether such mortality scales up to a limiting or regulating factor of recruitment. Disease-associated mortality of fishes in the marine environment often goes undetected or is observed sporadically, and parasite-induced mortality may be compensatory, for example owing to predation [55]. Some examples include parasite-associated mortality of juvenile coho salmon (Oncorhynchus kisutch) with infestations of the digenean trematode Apophallus sp. [56], mortality of European eels (Anguilla anguilla) infested with the nematode Anguillicoloides (=Anguillicola) crassus [57], and mortality of juvenile Pacific salmon (Oncorhynchus spp.) owing to the trematode Nonophyetus salmincola [58].
However, none of the above example studies of parasite-related mortality have been able to establish if parasitism is linked with declines in host fish recruitment, or is otherwise offset by compensatory mechanisms. From meta-analysis of taxonomically diverse field- and laboratory-based experimental studies, parasitized hosts tend to be approximately 2.6 times more likely to die compared with control groups, and that odds ratio is likely to be higher for fishes [10]. It has to be acknowledged, however, that variation in the odds ratio of host mortality is influenced by both taxon and latitude, and differs for (direct) parasite-associated effects and (indirect) predation-mediated influences of parasites on host survivorship [10]. Our analysis for one species of fish yielded a meta-analytic mean odds ratio of 1.29 (95% CI: 1.18–1.42), which is relatively low compared with reported values [10], and yet which still implies a high proportion (39%) of host mortality attributable to a specialist ectoparasite.
In contrast to the foregoing [56–58], our analysis of experimental manipulations of emigrant Atlantic salmon smolts permitted an explicit, though indirect, quantitative test of the significance of host mortality attributable to a specialist ectoparasite. All experimental fish were known to be free of L. salmonis infection at the start of the experiment, because the parasite is acquired only following seawater entry by the emigrant smolt. Fish in the control and treated groups all were identifiable by tags, and surviving adults that successfully completed the return marine migration in subsequent years could be recaptured because of the fidelity of salmon to their natal (or experimental release) river. Moreover, both the prophylactic experimental treatments applied have specific effects in controlling arthropod ectoparasites and emamectin benzoate is widely used in the aquaculture industry specifically to control L. salmonis infestations on cultured salmon.
The three analytical approaches we used reveal a key insight into interpreting the results of salmon and sea lice studies that formed the basis of our meta-analysis. While the assembled data indicate a clear consistency in direction of parasiticide effects on survival, the magnitude of the odds ratio is relatively small. This is owing to the very high marine mortality that naturally affects both control and treatment groups. This, perhaps, underlies the conclusions of some studies that the effect is biologically insignificant relative to the overall mortality within a salmon life cycle [40,46]. However, our analyses based on paired sample t-tests and mixed-effects models allowed us to account for the high natural mortality and isolate the estimated loss of recruitment owing to parasitism, revealing a large effect of parasites. Precisely because natural mortality rates are high, even a proportionally small additive mortality from parasites can amount to a large loss in salmon recruitment.
It is rare to identify and quantify the factors that affect population dynamics of marine fishes [15] as well as the role of infectious disease in conservation [59]. Our results indicate that parasite-associated mortality may cause the closure of some fisheries when conservation targets of return adult abundances are not being met. However, the implications of our results may be most acute for small populations in small river systems. Due largely to the fidelity to their natal rivers, populations of Atlantic salmon typically show substantial genetic structuring and variability that is considered adaptive [60]. Small river systems that support salmon populations of low effective population size [61] will be especially vulnerable. The concern therefore is not only for a 39 per cent loss in salmon abundance, but also the loss of genetic variability and its associated potential for adaptation to other environmental changes. More broadly, and in contrast to the conclusions from two of the original studies [40,46], our results supply manipulative field experimental evidence at a large spatial scale that parasitism may be a significant limiting factor for marine fishes, fisheries and conservation.
Acknowledgements
We are grateful to Brendan Connors for comments on the manuscript. This work was supported by funding from the University of Otago, the Norwegian Directorate for Nature Management, the Canada Research Chair Programme at UPEI, the Norwegian Institute of Marine Research and EU Framework 5 (Q5RS-2002-00730).
References
- 1.Daszak P, Cunningham AA, Hyatt AD. 2000. Emerging infectious diseases of wildlife: threats to biodiversity and human health. Science 287, 443–449 10.1126/science.287.5452.443 (doi:10.1126/science.287.5452.443) [DOI] [PubMed] [Google Scholar]
- 2.Ferguson NM, Donnelly CA, Anderson RM. 2001. Transmission intensity and impact of control policies on the foot and mouth epidemic in Great Britain. Nature 413, 542–548 10.1038/35097116 (doi:10.1038/35097116) [DOI] [PubMed] [Google Scholar]
- 3.de Castro F, Bolker B. 2005. Mechanisms of disease-induced extinction. Ecol. Lett. 8, 117–126 10.1111/j.1461-0248.2004.00693.x (doi:10.1111/j.1461-0248.2004.00693.x) [DOI] [Google Scholar]
- 4.Pedersen AB, Jones KE, Nunn CL, Altizer S. 2007. Infectious diseases and extinction risk in wild mammals. Conserv. Biol. 21, 1269–1279 10.1111/j.1523-1739.2007.00776.x (doi:10.1111/j.1523-1739.2007.00776.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jansen PA, Kristoffersen AB, Viljugrein H, Jimenez D, Aldrin M, Stien A. 2012. Sea lice as a density-dependent constraint to salmonid farming. Proc. R. Soc. B 279, 2330–2338 10.1098/rspb.2012.0084 (doi:10.1098/rspb.2012.0084) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hatcher MJ, Dick JTA, Dunn AM. 2012. Diverse effects of parasites in ecosystems: linking interdependent processes. Front. Ecol. Env. 10, 186–194 10.1890/110016 (doi:10.1890/110016) [DOI] [Google Scholar]
- 7.Fenton A, Brockhurst MA. 2008. The role of specialist parasites in structuring host communities. Ecol. Res. 23, 795–804 10.1007/s11284-007-0440-6 (doi:10.1007/s11284-007-0440-6) [DOI] [Google Scholar]
- 8.Hudson PJ, Rizzoli A, Grenfell BT, Heesterbeek H, Dobson AP. 2001. The ecology of wildlife diseases. Oxford, UK: Oxford University Press [Google Scholar]
- 9.Thomas F, Renaud F, Guegan JF. 2005. Parasitism and ecosystems. Oxford, UK: Oxford University Press [Google Scholar]
- 10.Robar N, Burness G, Murray DL. 2010. Tropics, trophics and taxonomy: the determinants of parasite-associated host mortality. Oikos 119, 1273–1280 10.1111/j.1600-0706.2009.18292.x (doi:10.1111/j.1600-0706.2009.18292.x) [DOI] [Google Scholar]
- 11.Hudson PJ, Dobson AP, Newborn D. 1998. Prevention of population cycles by parasite removal. Science 282, 2256–2258 10.1126/science.282.5397.2256 (doi:10.1126/science.282.5397.2256) [DOI] [PubMed] [Google Scholar]
- 12.Redpath SM, Mougeot F, Leckie FM, Elston DA, Hudson PJ. 2006. Testing the role of parasites in driving the cyclic population dynamics of a gamebird. Ecol. Lett. 9, 410–418 10.1111/j.1461-0248.2006.00895.x (doi:10.1111/j.1461-0248.2006.00895.x) [DOI] [PubMed] [Google Scholar]
- 13.Fogarty MJ, Sissenwine MP, Cohen EB. 1991. Recruitment variability and the dynamics of exploited marine populations. Trends Ecol. Evol. 6, 241–246 10.1016/0169-5347(91)90069-a (doi:10.1016/0169-5347(91)90069-a) [DOI] [PubMed] [Google Scholar]
- 14.Anderson CNK, Hsieh CH, Sandin SA, Hewitt R, Hollowed A, Beddington J, May RM, Sugihara G. 2008. Why fishing magnifies fluctuations in fish abundance. Nature 452, 835–839 10.1038/nature06851 (doi:10.1038/nature06851) [DOI] [PubMed] [Google Scholar]
- 15.Myers RA. 1998. When do environment–recruitment correlations work? Rev. Fish Biol. Fisher. 8, 285–305 10.1023/a:1008828730759 (doi:10.1023/a:1008828730759) [DOI] [Google Scholar]
- 16.Dobson AP, May RM. 1987. The effects of parasites on fish populations: theoretical aspects. Int. J. Parasitol. 17, 363–370 10.1016/0020-7519(87)90111-1 (doi:10.1016/0020-7519(87)90111-1) [DOI] [PubMed] [Google Scholar]
- 17.Lafferty KD, Porter JW, Ford SE. 2004. Are diseases increasing in the ocean? Annu. Rev. Ecol. Evol. Syst. 35, 31–54 10.1146/annurev.ecolsys.35.021103.105704 (doi:10.1146/annurev.ecolsys.35.021103.105704) [DOI] [Google Scholar]
- 18.McCallum HI, Kuris A, Harvell CD, Lafferty KD, Smith GW, Porter J. 2004. Does terrestrial epidemiology apply to marine systems? Trends Ecol. Evol. 19, 585–591 10.1016/j.tree.2004.08.009 (doi:10.1016/j.tree.2004.08.009) [DOI] [Google Scholar]
- 19.Wood CL, Lafferty KD, Micheli F. 2010. Fishing out marine parasites? Impacts of fishing on rates of parasitism in the ocean. Ecol. Lett. 13, 761–775 10.1111/j.1461-0248.2010.01467.x (doi:10.1111/j.1461-0248.2010.01467.x) [DOI] [PubMed] [Google Scholar]
- 20.Krkošek M, Ford JS, Morton A, Lele S, Myers RA, Lewis MA. 2007. Declining wild salmon populations in relation to parasites from farm salmon. Science 318, 1772–1775 10.1126/science.1148744 (doi:10.1126/science.1148744) [DOI] [PubMed] [Google Scholar]
- 21.Diana JS. 2009. Aquaculture production and biodiversity conservation. Bioscience 59, 27–38 10.1525/bio.2009.59.1.7 (doi:10.1525/bio.2009.59.1.7) [DOI] [Google Scholar]
- 22.Toranzo AE, Magarinos B, Romalde JL. 2005. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 246, 37–61 10.1016/j.aquaculture.2005.01.002 (doi:10.1016/j.aquaculture.2005.01.002) [DOI] [Google Scholar]
- 23.Johnson SC, Treasurer JW, Bravo S, Nagasawa K, Kabata Z. 2004. A review of the impact of parasitic copepods on marine aquaculture. Zool. Stud. 43, 229–243 [Google Scholar]
- 24.Asche F, Hansen H, Tveteras R, Tveteras S. 2009. The salmon disease crisis in Chile. Mar. Res. Econ. 24, 405–411 [Google Scholar]
- 25.Ward TM, Hoedt F, McLeay L, Dimmlich WF, Kinloch M, Jackson G, McGarvey R, Rogers PJ, Jones K. 2001. Effects of the 1995 and 1998 mass mortality events on the spawning biomass of sardine, Sardinops sagax, in South Australian waters. ICES J. Mar. Sci. 58, 865–875 10.1006/jmsc.2001.1077 (doi:10.1006/jmsc.2001.1077) [DOI] [Google Scholar]
- 26.Heuch PA, Bjørn PA, Finstad B, Holst JC, Asplin L, Nilsen F. 2005. A review of the Norwegian ‘National Action Plan Against Salmon Lice on Salmonids’: the effect on wild salmonids. Aquaculture 246, 79–92 10.1016/j.aquaculture.2004.12.027 (doi:10.1016/j.aquaculture.2004.12.027) [DOI] [Google Scholar]
- 27.Costello MJ. 2006. Ecology of sea lice parasitic on farmed and wild fish. Trends Parasitol. 22, 475–483 10.1016/j.pt.2006.08.006 (doi:10.1016/j.pt.2006.08.006) [DOI] [PubMed] [Google Scholar]
- 28.Pike AW, Wadsworth SL. 1999. Sealice on salmonids: their biology and control. Adv. Parasitol. 44, 233–337 10.1016/S0065-308X(08)60233-X (doi:10.1016/S0065-308X(08)60233-X) [DOI] [PubMed] [Google Scholar]
- 29.Jones SRM, Beamish RJ. 2011. Salmon lice: an integrated approach to understanding parasite abundance and distribution. Oxford, UK: Wiley-Blackwell [Google Scholar]
- 30.Hindar K, Hutchings JA, Diserud OH, Fiske P. 2010. Stock, recruitment, and exploitation. In Atlantic salmon ecology (eds Aas Ø, Einum S, Klemetsen A, Skurdal J.), pp. 299–311 Oxford, UK: Wiley-Blackwell [Google Scholar]
- 31.Ford JS, Myers RA. 2008. A global assessment of salmon aquaculture impacts on wild salmonids. PLoS Biol. 6, e33. 10.1371/journal.pbio.0060033 (doi:10.1371/journal.pbio.0060033) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Otero J, Jensen AJ, L'Abée-Lund JH, Stenseth NC, Storvik GO, Vøllestad LA. 2011. Quantifying the ocean, freshwater and human effects on year-to-year variability of one-sea-winter Atlantic salmon angled in multiple Norwegian rivers. PLoS ONE 6, e24005. 10.1371/journal.pone.0024005 (doi:10.1371/journal.pone.0024005) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Murray AG, Gillibrand PA. 2006. Modelling salmon lice dispersal in Loch Torridon, Scotland. Mar. Pollut. Bull. 53, 128–135 10.1016/j.marpolbul.2005.09.013 (doi:10.1016/j.marpolbul.2005.09.013) [DOI] [PubMed] [Google Scholar]
- 34.Poulin R. 2007. Are there general laws in parasite ecology? Parasitology 134, 763–776 10.1017/s0031182006002150 (doi:10.1017/s0031182006002150) [DOI] [PubMed] [Google Scholar]
- 35.Poulin R. 2011. The many roads to parasitism: a tale of convergence. Adv. Parasitol. 74, 1–40 10.1016/b978-0-12-385897-9.00001-x (doi:10.1016/b978-0-12-385897-9.00001-x) [DOI] [PubMed] [Google Scholar]
- 36.Todd CD, Whyte BDM, MacLean JC, Walker AM. 2006. Ectoparasitic sea lice (Lepeophtheirus salmonis and Caligus elongatus) infestations of wild, adult, one sea-winter Atlantic salmon Salmo salar returning to Scotland. Mar. Ecol. Prog. Ser. 328, 183–193 10.3354/meps328183 (doi:10.3354/meps328183) [DOI] [Google Scholar]
- 37.Todd CD, Walker AM, Hoyle JE, Northcott SJ, Walker AF, Ritchie MG. 2000. Infestations of wild adult Atlantic salmon (Salmo salar L.) by the ectoparasitic copepod sea louse Lepeophtheirus salmonis Krøyer: prevalence, intensity and the spatial distribution of males and females on the host fish. Hydrobiologia 429, 181–196 10.1023/A:1004031318505 (doi:10.1023/A:1004031318505) [DOI] [Google Scholar]
- 38.Marty G, Saksida S, Quinn TJ. 2010. Relationship of farm salmon, sea lice, and wild salmon populations. Proc. Natl Acad. Sci. USA 107, 22 599–22 604 10.1073/pnas.1009573108 (doi:10.1073/pnas.1009573108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Krkošek M, Connors BM, Morton A, Lewis MA, Dill LM, Hilborn R. 2011. Effects of parasites from salmon farms on productivity of wild salmon. Proc. Natl Acad. Sci. USA 108, 14 700–14 704 10.1073/pnas.1101845108 (doi:10.1073/pnas.1101845108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Jackson D, et al. 2011. An evaluation of the impact of early infestation with the salmon louse Lepeophtheirus salmonis on the subsequent survival of outwardly migrating Atlantic salmon, Salmo salar L., smolts. Aquaculture 320, 159–163 10.1016/j.aquaculture.2011.03.029 (doi:10.1016/j.aquaculture.2011.03.029) [DOI] [Google Scholar]
- 41.Beamish RJ, Jones S, Neville CE, Sweeting R, Karreman G, Saksida S, Gorden E. 2006. Exceptional marine survival of pink salmon that entered the marine environment in 2003 suggests that farmed Atlantic salmon and Pacific salmon can coexist successfully in a marine ecosystem on the Pacific coast of Canada. ICES J. Mar. Sci. 63, 1326–1337 10.1016/j.icesjms.2006.04.011 (doi:10.1016/j.icesjms.2006.04.011) [DOI] [Google Scholar]
- 42.Gargan P, Forde G, Hazon N, Russell DJF, Todd CD. 2012. Evidence for sea lice-induced marine mortality of Atlantic salmon (Salmo salar) in western Ireland from experimental releases of ranched smolts treated with emamectin benzoate. Can. J. Fish. Aquat. Sci. 69, 343–353 10.1139/f2011-155 (doi:10.1139/f2011-155) [DOI] [Google Scholar]
- 43.Gargan P, Tully O, Poole WR. 2002. The relationship between sea lice Infestation, sea lice production and sea trout survival in Ireland, 1992–2001. In Salmon on the edge (ed. Mills D.), pp. 119–135 Edinburgh, UK: Atlantic Salmon Trust/Atlantic Salmon Federation [Google Scholar]
- 44.Costello MJ. 2009. How sea lice from salmon farms may cause wild salmonid declines in Europe and North America and be a threat to fishes elsewhere. Proc. R. Soc. B 276, 3385–3394 10.1098/rspb.2009.0771 (doi:10.1098/rspb.2009.0771) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Skilbrei OT, Wennevik V. 2006. Survival and growth of sea-ranched Atlantic salmon, Salmo salar L., treated against sea lice before release. ICES J. Mar. Sci. 63, 1317–1325 10.1016/j.icesjms.2006.04.012 (doi:10.1016/j.icesjms.2006.04.012) [DOI] [Google Scholar]
- 46.Jackson D, et al. 2011. Impact of early infestation with the salmon louse Lepeophtheirus salmonis on the subsequent survival of outwardly migrating Atlantic salmon smolts from a number of rivers on Ireland's south and west coasts. Aquaculture 319, 37–40 10.1016/j.aquaculture.2011.06.042 (doi:10.1016/j.aquaculture.2011.06.042) [DOI] [Google Scholar]
- 47.Hvidsten NA, Finstad B, Kroglund F, Johnsen BO, Strand R, Arnekleiv JV, Bjørn PA. 2007. Does increased abundance of sea lice influence survival of wild Atlantic salmon post-smolt? J. Fish. Biol. 71, 1639–1648 10.1111/j.1095-8649.2007.01622.x (doi:10.1111/j.1095-8649.2007.01622.x) [DOI] [Google Scholar]
- 48.Stone J, Sutherland IH, Sommerville C, Richards RH, Varma KJ. 2000. Commercial trials using emamectin benzoate to control sea lice Lepeophtheirus salmonis infestations in Atlantic salmon Salmo salar. Dis. Aquat. Org. 41, 141–149 10.3354/dao041141 (doi:10.3354/dao041141) [DOI] [PubMed] [Google Scholar]
- 49.DerSimonian R, Laird N. 1986. Meta-analysis in clinical trials. Control. Clin. Trials 7, 177–188 [DOI] [PubMed] [Google Scholar]
- 50.Klein JP, Moeschberger ML. 2003. Survival analysis. New York, NY: Springer [Google Scholar]
- 51.Skilbrei OT, Glover KA, Samuelsen OB, Lunestad BT. 2008. A laboratory study to evaluate the use of emamectin benzoate in the control of sea lice in sea-ranched Atlantic salmon (Salmo salar L.). Aquaculture 285, 2–7 10.1016/j.aquaculture.2008.07.055 (doi:10.1016/j.aquaculture.2008.07.055) [DOI] [Google Scholar]
- 52.Jonsson N, Jonsson B, Hansen LP. 2003. The marine survival and growth of wild and hatchery-reared Atlantic salmon. J. Appl. Ecol. 40, 900–911 10.1046/j.1365-2664.2003.00851.x (doi:10.1046/j.1365-2664.2003.00851.x) [DOI] [Google Scholar]
- 53.Gillibrand PA, Kate JW. 2007. Dispersal of sea lice larvae from salmon farms: a model study of the influence of environmental conditions and larval behaviour. Aquat. Biol. 1, 63–75 10.3354/ab00006 (doi:10.3354/ab00006) [DOI] [Google Scholar]
- 54.Bakke TA, Harris PD. 1998. Diseases and parasites in wild Atlantic salmon (Salmo salar) populations. Can. J. Fish. Aquat. Sci. 55(S1), 247–266 10.1139/d98-021 (doi:10.1139/d98-021) [DOI] [Google Scholar]
- 55.Krkošek M, et al. 2011. Fish farms, parasites, and predators: Implications for salmon population dynamics. Ecol. Appl. 21, 897–914 10.1890/09-1861.1 (doi:10.1890/09-1861.1) [DOI] [PubMed] [Google Scholar]
- 56.Ferguson JA, Koketsu W, Ninomiya I, Rossignol PA, Jacobson KC, Kent ML. 2011. Mortality of coho salmon (Oncorhynchus kisutch) associated with burdens of multiple parasite species. Int. J. Parasitol. 41, 1197–1205 10.1016/j.ijpara.2011.07.005 (doi:10.1016/j.ijpara.2011.07.005) [DOI] [PubMed] [Google Scholar]
- 57.Kennedy CR. 2007. The pathogenic helminth parasites of eels. J. Fish Dis. 30, 319–334 10.1111/j.1365-2761.2007.00821.x (doi:10.1111/j.1365-2761.2007.00821.x) [DOI] [PubMed] [Google Scholar]
- 58.Jacobson KC, Teel D, Van Doornik DM, Casillas E. 2008. Parasite-associated mortality of juvenile Pacific salmon caused by the trematode Nanophyetus salmincola during early marine residence. Mar. Ecol. Prog. Ser. 354, 235–244 10.3354/meps07232 (doi:10.3354/meps07232) [DOI] [Google Scholar]
- 59.Smith KF, Sax DF, Lafferty KD. 2006. Evidence for the role of infectious disease in species extinction and endangerment. Conserv. Biol. 20, 1349–1357 10.1111/j.1523-1739.2006.00524.x (doi:10.1111/j.1523-1739.2006.00524.x) [DOI] [PubMed] [Google Scholar]
- 60.Garcia de Leaniz CG, et al. 2007. A critical review of adaptive genetic variation in Atlantic salmon: implications for conservation. Biol. Rev. 82, 173–211 10.1111/j.1469-185X.2006.00004.x (doi:10.1111/j.1469-185X.2006.00004.x) [DOI] [PubMed] [Google Scholar]
- 61.Nikolic N, Butler JRA, Baglinière J-L, Laughton R, McMyn IAG, Chevalet C. 2009. An examination of genetic diversity and effective population size in Atlantic salmon populations. Genet. Res. 91, 395–412 10.1017/s0016672309990346 (doi:10.1017/s0016672309990346) [DOI] [PubMed] [Google Scholar]