
Figure 5. MC migration requires both SphK1 and SphK2.
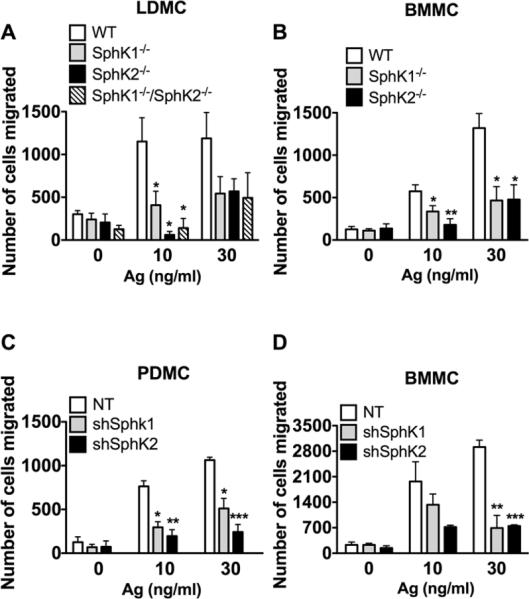
Fetal liver (A) and bone marrow (B) MC were derived from SphK1-, SphK2- mice (A, B) or SphK1/SphK2 double-deficient (A) mice. Alternatively, shRNA was used to silence SphK1 and SphK2 in peritoneal (C) and bone marrow-derived (D) MC. The ability of MC to migrate towards increasing concentrations of antigen was measured using transwell assays. Cells were placed in the upper inserts and the indicated concentrations of antigen were placed in the lower chamber. Cells were allowed to migrate for three hours. The number of cells migrated was quantified using Cyquant-GR dye. Data represent the mean ± S.E. of at least three individual cultures. Statistical significance compared to WT (A and B) or nontarget (NT) control (C and D), * P<0.05, **P<0.01, ***P<0.001. Despite the trends, some comparisons did not achieve statistical significance due to variability in the total of cells migrating between different cultures. However, in each experiment silencing or genetic deletion of SphK1 or SphK2 showed an average of at least a 50% reduction in mast cell migration regardless of the MC type. Deletion or silencing of SphK1 or SphK2 also resulted in impairment of MC migration towards SCF (30 ng/ml), a potent MC chemoattractant. In LDMC migration to SCF was as follows: WT: 3402±823 ; SphK1-/-:1450± 650 ; SphK2-/-: 719±167; SphK1-/- /SphK2-/-: 473 ± 148 (mean+ SEM).
(mean+ SEM).