

Abstract
High aldehyde dehydrogenase (ALDH) activity has been associated with stem and progenitor cells in various tissues. Human cord blood and bone marrow ALDH-bright (ALDHbr) cells have displayed angiogenic activity in preclinical studies and have been shown to be safe in clinical trials in patients with ischemic cardiovascular disease. The presence of ALDHbr cells in the heart has not been evaluated so far. We have characterized ALDHbr cells isolated from mouse hearts. One percent of nonmyocytic cells from neonatal and adult hearts were ALDHbr. ALDHvery-br cells were more frequent in neonatal hearts than adult. ALDHbr cells were more frequent in atria than ventricles. Expression of ALDH1A1 isozyme transcripts was highest in ALDHvery-br cells, intermediate in ALDHbr cells, and lowest in ALDHdim cells. ALDH1A2 expression was highest in ALDHvery-br cells, intermediate in ALDHdim cells, and lowest in ALDHbr cells. ALDH1A3 and ALDH2 expression was detectable in ALDHvery-br and ALDHbr cells, unlike ALDHdim cells, albeit at lower levels compared with ALDH1A1 and ALDH1A2. Freshly isolated ALDHbr cells were enriched for cells expressing stem cell antigen-1, CD34, CD90, CD44, and CD106. ALDHbr cells, unlike ALDHdim cells, could be grown in culture for more than 40 passages. They expressed sarcomeric α-actinin and could be differentiated along multiple mesenchymal lineages. However, the proportion of ALDHbr cells declined with cell passage. In conclusion, the cardiac-derived ALDHbr population is enriched for progenitor cells that exhibit mesenchymal progenitor-like characteristics and can be expanded in culture. The regenerative potential of cardiac-derived ALDHbr cells remains to be evaluated.
1. Introduction
Growing evidence suggests the adult heart may harbor resident stem and progenitor cells that participate in adaptive responses to myocardial injury, and possibly in cellular homeostasis under normal conditions. In most studies, the prospective isolation of cardiac stem cells has relied upon the use of specific antibodies that recognize cell-surface antigens expressed by stem cells in other tissues, particularly by hematopoietic stem cells (HSCs). Cardiac stem cells expressing stem cell antigen-1 (Sca-1) or the stem cell factor receptor, c-kit (CD117), have been described [1, 2]. A distinct population of cardioblasts expressing the transcription factor islet-1 (Isl-1) and entering fully differentiated cardiomyocyte lineages has also been identified [3, 4]. However, the expression of cell-surface epitopes can vary with the metabolic state of the cell and the experimental conditions used. Therefore, no individual cell-surface antigen marker definitely identifies a single entity of cardiac stem cell. Functional properties of tissue-resident stem cells that could be used for their prospective isolation, regardless of cell-surface marker expression, have been intensely searched for. It has been proposed that, irrespective of their lineal origin, stem cells may share common mechanisms to regulate self-renewal and differentiation, and that candidate “stemness” genes may serve as universal stem cell markers.
Cai et al. [5] proposed that high aldehyde dehydrogenase (ALDH) activity is one of a small set of common characteristics shared by stem cells among tissues (recently reviewed by Balber [6]). Few ALDH-bright (ALDHbr) pluripotential cells, measured by Aldefluor stain, gave rise to all somatic and reproductive cell lineages in tunicates [7, 8]. ALDH is a cytosolic enzyme responsible for the oxidation of intracellular aldehydes. It plays important roles in oxidation of alcohol and vitamin A, and in chemoresistance to cyclophosphamide. ALDH superfamily is highly conserved across species [9]. Nineteen ALDH isoforms are known in human. ALDHbr, side scatter-low (SSClo) populations from murine and human bone marrow (BM) [10–13], umbilical cord blood (UCB) [14–19], cytokine-mobilized peripheral blood [20], and circulating blood cells [21, 22] are enriched in stem and progenitor cells. The ALDHbrSSClo population includes virtually all human CD34+ and CD133+ cells that establish long-term, multilineage hematopoietic colonies in culture or long-term, multilineage xenografts in immunodeficient mice [16]. More recently, ALDHbr stem cells have been identified in nonhematopoietic systems, such as circulating endothelial progenitor cells (EPCs) [21, 22], neural stem cells [23, 24], human muscle precursor cells with high myogenic activity [25, 26], colonic stem cells [27], and mammary stem cells [28]. Moreover, ALDHbr cancer stem cells have been described in multiple types of cancer and shown to predict poor clinical outcomes in various contexts [27–29].
We therefore asked whether the postnatal heart harbors ALDHbr cells, as measured by Aldefluor stain. We found that the neonatal as well as the adult mouse heart harbors a relatively small population of ALDHbr cells (≈1% of all nonmyocytic cells present in the heart), which is enriched for cells expressing Sca-1 and other progenitor cell markers. ALDHbr cells, but not ALDHdim cells, adhered to plastic and grew in culture. The expanded population expressed sarcomeric α-actinin, a cardiac marker, along with mesenchymal stem cell (MSC) markers. It could be induced to differentiate along multiple mesenchymal cell lineages. These results suggest that ALDH may mark a population of cardiac-resident, MSC-like progenitor cells that possess superior ex vivo growth characteristics.
2. Methods
2.1. Mice and Cell Isolation
Neonatal (postnatal day 1), young adult (8 week-old), and aging (24 month-old) C57Bl/6 mice were purchased from Charles River Laboratories (France). Immediately after the sacrifice of the mice, the chest was opened, a canula was introduced into the left ventricular cavity, an incision was made in the right atrial wall, and the heart was perfused with heparinized PBS. The explanted heart was washed in PBS and cut into small pieces that were then placed in a Falcon tube (50 mL) containing 5 mL RPMI 1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 12.5 μL Liberase Blendzyme 4 and 25 μL DNAse I (both from Roche, Basle, Switzerland), and incubated for 45 min at 37°C under gentle shaking. The cell suspension was filtered through a 70 μm-filter using a syringe plunger, washed in PBS, and centrifuged. The pellet was resuspended in 100 μL Dead Cell Removal Kit microbeads solution (Miltenyi, Bergisch Gladbach, Germany), incubated for 15 min at RT, changed to Dead Cell Removal Buffer, and passaged through a Miltenyi LS column. The flow-through was centrifuged and the pellet was resuspended in FACS buffer. In a subset of experiments, atria and ventricles were processed separately to measure ALDHbr cells in the different cardiac chambers of origin.
2.2. Flow Cytometric Analyses
Cells were washed in PBS, resuspended in Aldefluor buffer, and then reacted with Aldefluor substrate (STEMCELL Technologies, Vancouver, BC, Canada) according to the manufacturer's instructions. After the ALDH enzyme reaction, cells were washed, resuspended in cold Aldefluor buffer, and maintained on ice during all subsequent manipulations. A forward scatter (FSC) versus side scatter (SSC) cytogram was used to gate signals from cells, and an Aldefluor fluorescence versus SSC cytogram was constructed. A unique ALDHbr population was present in Aldefluor-reacted samples and absent in cells in the samples treated with the specific inhibitor of ALDH, diethylaminobenzaldehyde (DEAB). Cells incubated with Aldefluor substrate and DEAB were used to establish baseline fluorescence of these cells and to define the ALDHbr and the ALDHvery-br region as less than 0.1% and 0% of total events, respectively. Cell incubation with Aldefluor substrate in the absence of inhibitor induced a shift in FL1 fluorescence defining the ALDHdim, the ALDHbr, and the ALDHvery-br population. To assess cell-surface antigen expression, cells were incubated with antibody for 20 min, washed, and resuspended in cold Aldefluor buffer. Flow cytometric analyses were performed on a FACSCalibur instrument (Becton Dickinson, San Jose, CA, USA) operating at 488 nm excitation with standard emission filters. Aldefluor fluorescence was measured in FL1 and APC in FL4. Gates used to resolve antigen-expressing cells were set using appropriate isotype-matched control Abs. Data files containing at least 2,000 ALDHbr cells were acquired for analysis using the CellQuest software (Becton Dickinson).
2.3. Immunostaining
Culture-expanded ALDHbr cells grown in Lab-Tek chambers were fixed with 1% paraformaldehyde (PFA) for 10 min, followed by blocking solution (1x PBS, 1% BSA, 0,3% Triton) for 1 h, and incubated with rat anti-mouse c-Kit/CD117 mAb (clone 3C1; Miltenyi) coupled to APC for 1 h at RT, followed by goat anti-rat IgG coupled to Alexa 488 (1 : 400 dilution; Molecular Probes, Life Technologies, Grand Island, NY, USA). To detect type II collagen, cells were fixed in PFA, incubated in blocking solution for 1 h, and then with mouse monoclonal anti-type II collagen 4 Abs coktail (1 : 100 dilution; Chondrex, Redmond, WA, USA), followed by goat anti-mouse IgG coupled to Alexa 488 (1 : 400). To detect sarcomeric α-actinin, cells were fixed with 4% PFA for 1 h, incubated in blocking solution for 1 h, and then with mouse antisarcomeric α-actinin mAb (1 : 750 dilution; clone EA-53, Sigma-Aldrich, St. Louis, MO, USA) at 4°C overnight, followed by goat anti-mouse IgG coupled to Alexa 488 (1 : 400). To detect α-smooth muscle actin (α-SMA), cells were stained with anti-α-SMA rabbit polyclonal Ab (abcam5694; Abcam). Control sections were incubated with secondary Ab only. Nuclei were stained with Hoechst 33342 and slides were mounted with Mowiol. Pictures were taken with a Zeiss Axioplant 2 microscope (100x objective).
2.4. Antibodies
The following mAbs were used to detect the cell-surface antigen distribution of cardiac cells by flow cytometry: CD11b-APC (clone M1; Miltenyi), CD14-APC (clone Sa2-8; eBioscience; San Diego, CA, USA), CD29-PE (clone RTK2758; BioLegend), CD31-APC (clone 390; eBioscience), CD34-APC (clone RAM-34; eBioscience), CD38-APC (clone 90; eBioscience), CD40-APC (clone 1C10; eBioscience), CD44-APC (clone IM7; eBioscience), CD45-APC (clone 30-F11; eBioscience), CD90.2-APC (clone HIS51; eBioscience), CD105-PE (clone MJ7/18; eBioscience), CD106-PE (clone 429; eBioscience), c-Kit/CD117-APC (clone 3C1; Miltenyi), CD133-APC (clone 13A4; eBioscience), CD140b-APC (eBioscience), CD146-FITC (Miltenyi), Flk-1-APC (clone Avas12a1; eBioscience), Lineage Cell Detection Cocktail-Biotin mouse (Miltenyi; cat. no. 130-092-613), MHC class II-APC (clone M5; eBioscience), NG2 chondroitin sulfate proteoglycan (Chemicon/Millipore; cat. no. AB5320), Sca-1-APC (clone D7; eBioscience), rat IgG2a isotype control-APC (clone eBR2a; eBioscience), rat IgG2a isotype control-FITC (clone eBR2a; eBioscience), rat IgG2a isotype control-PE (clone eBR2a; eBioscience), rat mouse IgG1, κ isotype control (clone P3; eBioscience), and Arminian hamster IgG isotype control (clone eBio299Arm; eBioscience).
2.5. Real Time RT-PCR
Freshly isolated ALDHdim, ALDHbr, and ALDHvery-br cells from PBS-perfused hearts from 8-week-old mice (n = 4) were sorted using a Beckman Coulter MoFlo Astrios FACS system. Total mRNA from each cell subset was extracted using the RNeasy Micro kit (Qiagen). The different c-DNAs were generated using the Quantitect reverse transcription kit (Qiagen) from total mRNA obtained with a genomic DNA digestion step according to the manufacturer's instructions. cDNA (1 : 10 dilution) was used for quantification using the RT2 SYBR Green qPCR Kit (Qiagen) and the Rotor-Gene 2000 system (Qiagen) according to the manufacturer's instructions. Real-time PCR reactions (in triplicates) were set up in 10 μL reaction volume with 5 μL of RT2 SYBR Green mix, 0.4 μL of 10 μM RT2 qPCR Primer Assay, 3 μL cDNA, and 1.6 μL water. The polymerase was heat-activated for 10 min at 95°C, and the reactions were then cycled 50 times (95°C, 15 sec; 55°C, 40 sec; 72°C, 30 sec), followed by a melting step. Primers were obtained from sAbiosciences (Qiagen). Relative expression was calculated with the comparative ΔCt method using GUSB as a reference gene.
2.6. Ex Vivo Cultures of ALDHbr Cells
In two preliminary experiments, cells from enzymatically dissociated atria and ventricles were reacted separately with Aldefluor, and ALDHbr cells were sorted by FACS and placed in Corning Costar 6-well plates (Sigma) with no extracellular matrix protein coating. Ventricular ALDHbr cells grew poorly in culture, possibly as a result of a very long sorting procedure due to the scarcity of ALDHbr cells in the ventricular population. Therefore, atrial cells from 8-week-old mice were used in subsequent experiments (n = 3 per experiment), allowing for a marked abbreviation of the sorting procedure. ALDHbr cells were cultured in MesenCult medium (MesenCult MSC Basal Medium supplemented with serum-containing MesenCult MSC Stimulatory Supplements-Mouse; STEMCELL Technologies). ALDHdim cells were studied for comparison. To assess the impact of the culture medium on cell phenotype, ALDHbr sorted cells were also cultured in RPMI/FCS medium (RPMI 1640; Gibco, supplemented with 10% fetal calf serum). The AlamarBlue assay (Promega, Madison, WI, USA) was used to assess cell viability and growth. In a separate experiment, cells were cultured in the presence of imatinib (methanesulfonate salt, 0.1–10 μM; LC Laboratories, Woburn, MA, USA), a inhibitor of receptor tyrosine kinases encoded by c-kit and platelet-derived growth factor receptor B (PDGFRB). Cells were incubated with imatinib for 42 h (in triplicates) before AlamarBlue was added. Absorbance was measured at 570 nm and 595 nm every hour during 6 hours, and AlamarBlue reduction was calculated.
2.7. Differentiation Potential of Expanded ALDHbr Cells
Cells sorted on the basis of high ALDH activity were cultured for 8 months in MesenCult, which was then replaced by NH ChondroDiff Medium (for 21 days), NH OsteoDiff Medium (7 days), or AdipoDiff Medium (14 days; all from Miltenyi). Chondrogenic differentiation was assessed by immunostaining for type II collagen. To assess osteogenic and adipogenic differentiation, cells were incubated with 1% PFA for 10 min, followed by 2% Alizarin Red solution for 5 min and Oil Red-O solution for 15 min, respectively, and by 3 PBS-washes.
3. Results
3.1. ALDHbr Cells in Neonatal and Young Adult Hearts
Freshly isolated nonmyocytic cells from enzymatically dissociated, neonatal or 8-week-old hearts were analyzed by flow cytometry using the Aldefluor reagent. ALDHbr cells amounted to 1.00 ± 0.59% of all nonmyocytic cells in the neonatal heart (n = 3) and to 0.99 ± 0.55% of these cells in the young adult heart (n = 7, NS; Figure 1). Within the ALDHbr population, the percentage of ALDHvery-br cells in neonatal hearts was higher than in young adults (52.71 ± 6.87% versus 19.39 ± 2.42%; P < 0.05).
Figure 1.
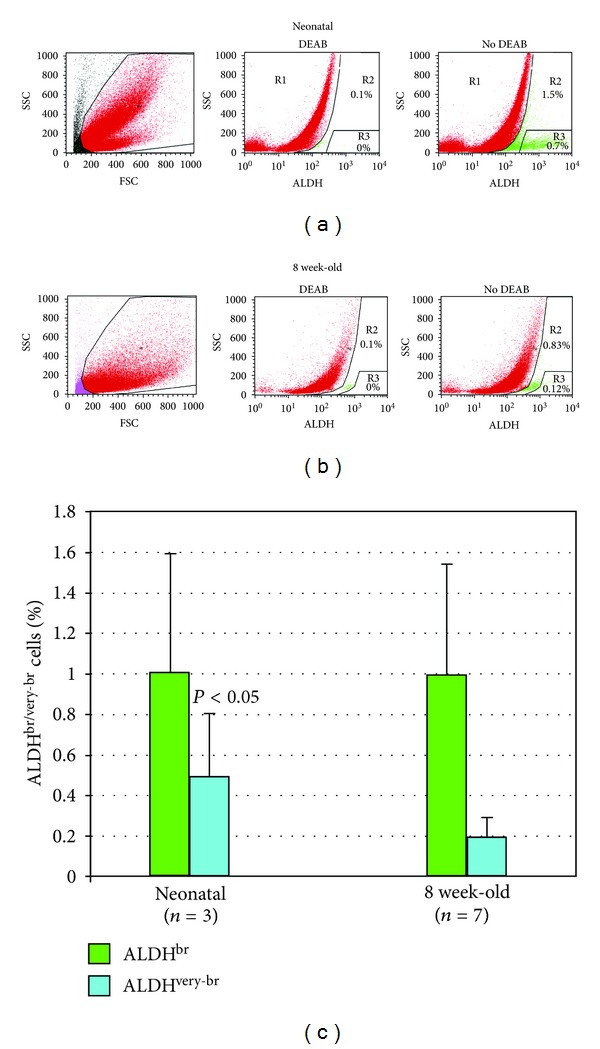
Identification of ALDHbr cells in populations isolated from whole hearts from neonatal and young adult mice. Flow cytometric analysis of ALDH activity (a). Neonatal mice. Left panel: cells were selected according to forward scatter (FSC) and side scatter (SSC) properties using the gated region. Middle panel: cells incubated with Aldefluor substrate and the specific inhibitor of ALDH, DEAB were used to establish baseline fluorescence of these cells and to define the ALDHbr (R2) and the ALDHvery-br (R3) region as less than 0.1% and 0% of total events, respectively. Right panel: cell incubation with Aldefluor substrate in the absence of inhibitor induced a shift in FL1 fluorescence defining the ALDHdim (R1), the ALDHbr (R2), and the ALDHvery-br (R3) populations (b). Eight-week-old mice (c). Bar histogram showing ALDHbr and ALDHvery-br cell percentages (mean, SD) in neonatal and young adult mice. P value refers to the difference between neonatal and 8 week-old mice.
3.2. ALDHbr Cells in Atria and Ventricles
In young adult mice, ALDHbr cells in the atrial population were more frequent than in the ventricular (10.29 ± 8.44% versus 1.05 ± 0.78%; n = 4; P < 0.05; Figure 2). Atrial ALDHbr cells predominantly exhibited SSClo properties but a subset showed intermediate-to-high SSC properties. Ventricular ALDHbr cells exhibited more homogeneous SSClo properties.
Figure 2.
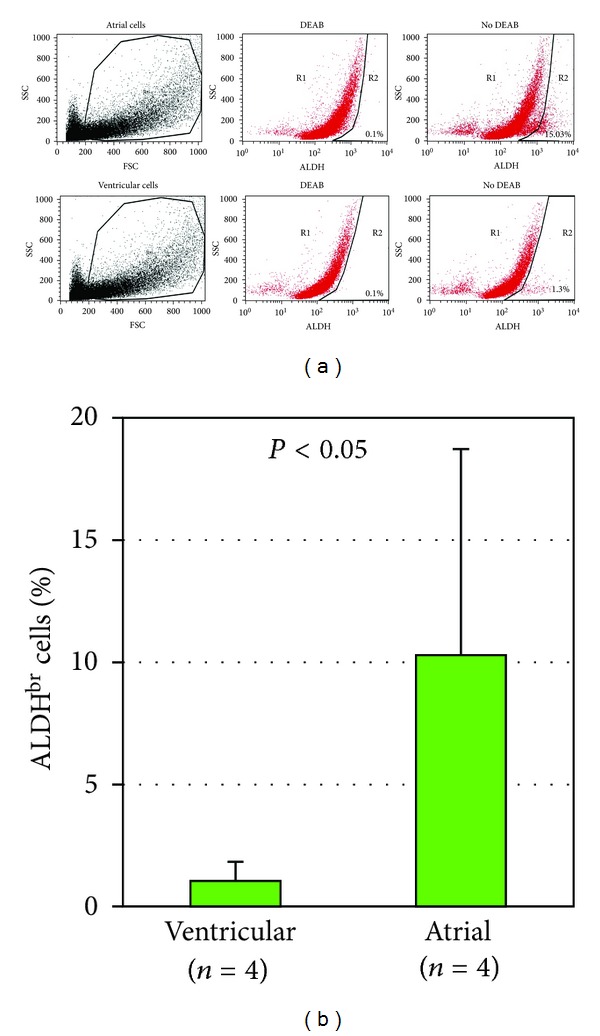
Identification of ALDHbr cells in the atrial and ventricular population from 8 week-old mice. Flow cytometric analysis of ALDH activity (a). Left panels: atrial and ventricular cells were selected according to forward scatter (FSC) and side scatter (SSC) properties using the gated region. Middle panels: cells incubated with Aldefluor substrate and the specific inhibitor of ALDH, DEAB, were used to establish baseline fluorescence of these cells and to define the ALDHbr region (R2) as less than 0.1% of total events. Right panels: cell incubation with Aldefluor in the absence of inhibitor induced a shift in FL1 fluorescence defining the ALDHdim (R1) and the ALDHbr (R2) population. In the example shown, not all of the atrial cells exhibiting a shift in FL1 fluorescence in the absence of inhibitor were found in the gated region R2, suggesting that atrial ALDHbr cells may have been underestimated (b). Bar histogram showing percent ALDHbr cells (mean, SD) in the ventricular and atrial population.
3.3. Immunophenotype of Freshly Isolated ALDHbr Cells
We used a panel of mAbs to determine the cell-surface marker profile of ALDHbr, ALDHvery-br, and ALDHdim cells in the freshly isolated, nonmyocytic cardiac-derived population in 8 weeks old as well as in 24 months old mice (Figure 3). Aging hearts contained 1.36% ALDHbr cells, of which 36.8% were ALDHvery-br (pooled sample from 5 hearts). A majority of ALDHbr and ALDHvery-br cells from young adult hearts stained positive for Sca-1 (≈60% and ≈70%, resp.). Substantial subsets (≈25–50%) of ALDHbr cells stained positive for CD34, CD106, CD44, CD90, and CD105 (Figure 4). The ALDHbr population was significantly enriched for cells expressing Sca-1, CD90, CD34, CD106, and CD44 (fold-changes: 4.2, 4.6, 6.2, 24.1, and 34.3, resp.,) whereas it was significantly depleted for cells expressing CD45 (leukocyte common antigen), CD31 (an endothelial marker), and CD38 (fold-changes: 0.5, 0.36, and 0.22 fold). The immunophenotype of ALDHbr cells derived from aging hearts was similar to that of ALDHbr cells from young adult hearts.
Figure 3.
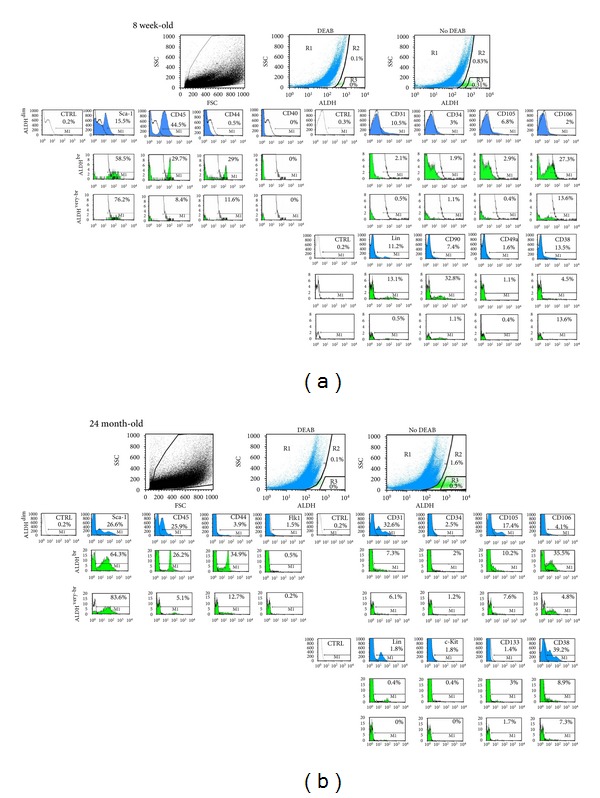
Freshly isolated ALDHbr cells from young adult hearts (upper panel; (a)) and from aging hearts (lower panel; (b)) were analyzed by flow cytometry for ALDH expression in combination with the expression of cell-surface markers. Cells were incubated with Aldefluor substrate and an inhibitor of ALDH (DEAB), or with Aldefluor alone, to establish R1, R2, and R3 gates for ALDHdim (blue), ALDHbr, and ALDHvery-br cells (both green), respectively. These populations were subsequently analyzed for expression of markers. Cytograms for each surface marker in each cell population are shown (the surface markers indicated for the cytograms of ALDHdim cells also refer to the cytograms of the respective ALDHbr and ALDHvery-br populations; corresponding cytograms are aligned vertically. Percentages of positive cells are indicated (CTRL; isotype-matched irrelevant IgG).
Figure 4.
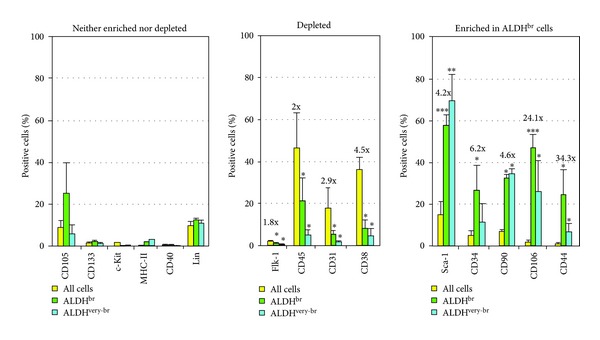
Flow cytometric analysis of the expression of cell-surface antigen markers in freshly isolated nonmyocytic cardiac cells from 8-week-old mice. Data are percentages (mean, SD) of positive cells for the indicated markers, subdivided into three groups: those significantly enriched, those significantly depleted, and those neither enriched nor depleted in the ALDHbr population. Yellow bars indicate bulk populations of cardiac-derived cells, green bars indicate ALDHbr cells, and blue bars indicate ALDHvery-br cells. Numbers are enrichment/depletion factors for the indicated surface markers in the ALDHbr population (***P < 0.001, *P < 0.05 versus ALDHdim cells).
3.4. mRNA Expression of Selected ALDH Isoforms
mRNA expression of ALDH1A1, ALDH1A2, ALDH1A3, and ALDH2 isoforms was measured by real-time RT-PCR in purified ALDHdim, ALDHbr, and ALDHvery-br cells (Figure 5). Gene transcripts of all of the four ALDH isoforms could be detected in ALDHbr and ALDHvery-br cells, whereas only ALDH1A1 and ALDH1A2 were detectable in ALDHdim cells. In ALDHbr and ALDHvery-br cells, ALDH1A1 was expressed at higher levels than ALDH1A3 (2−ΔCt = 4.0 and 4.7, resp.,) and ALDH2 (2−ΔCt = 4.8 and 8.8, resp.,) while ALDH1A2 was expressed at higher levels than ALDH1A3 (2−ΔCt = 1.7 and 12.3, resp.,) and ALDH2 (2−ΔCt = 2.1 and 23.2, resp.). In addition, ALDH1A1 was expressed at higher levels in ALDHbr and ALDHvery-br cells compared to ALDHdim cells (2−ΔΔCt = 2.0 and 3.5, resp.). ALDH1A2 expression was higher in ALDHvery-br cells compared to ALDHdim cells (2−ΔΔCt = 4.9) and was lowest in ALDHbr cells (2−ΔΔCt = 0.5 versus ALDHdim). mRNA expression of three genes implicated in angiogenesis was also measured. 2−ΔCt values for ALDHdim, ALDHbr, and ALDHvery-br cells were as follows: endoglin (CD105): 11.70, 10.41, and 14.86; ephrin B4: 2.98, 0.90, and 2.60; angiopoietin 1: undetectable, 0.01, and 0.008, respectively.
Figure 5.
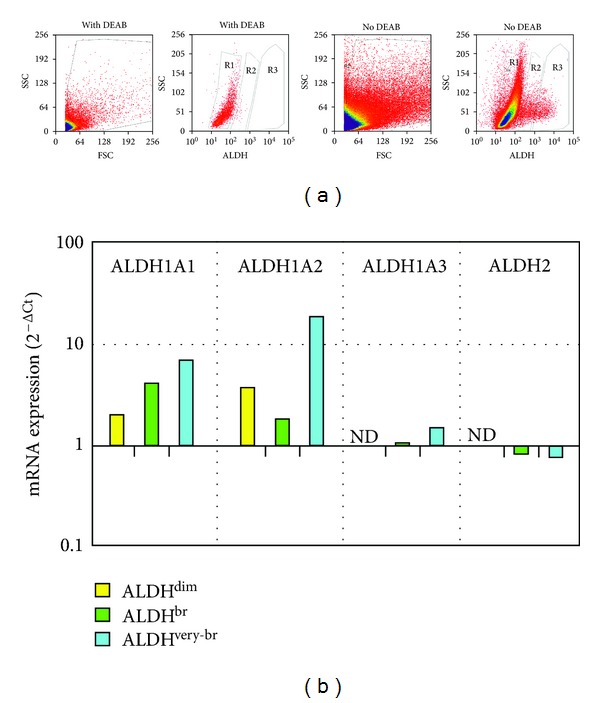
Real time RT-PCR analysis of gene transcripts of selected ALDH isoforms (a). FAC sorting of freshly isolated ALDHdim (R1), ALDHbr (R2), and ALDHvery-br (R3) atrial cells from 8-week-old mice (b). RT-PCR analysis of ALDH1A1, ALDH1A2, ALDH1A3, and ALDH2 gene transcripts (data are 2−ΔCt values using GUSB as a reference gene). ND: not detectable.
3.5. Ex Vivo Culture-Expansion of ALDHbr Cells
ALDHbr cells were purified from the atrial population, which was enriched for these cells compared to ventricles, in order to abbreviate the FACS procedure, thereby limiting cell damage. Purified atrial ALDHbr cells grew in culture, whereas ALDHdim cells did not, even when plated at a 10-fold higher density compared to their ALDHbr counterparts (Figure 6(a)). However, >95% ALDHdim cells appeared to be viable, as assessed by DAPI, immediately after the FACS procedure. Atrially derived bulk populations of nonmyocytic cells grew poorly. Purified ALDHbr cells gave rise to small numbers of cell colonies, each one apparently originating from a single cell, which formed a monolayer of plastic-adherent cells, which could be expanded for more than 40 passages (later passages were not tested). In one cell culture, the percentage of ALDHbr cells at passage 7 was 11%; however, this parameter was not systematically measured at various time points. Growth rates in MesenCult medium were higher than in RPMI/FCS medium (Figure 6(b)). This was confirmed using the AlamarBlue assay (Figure 6(c)). Imatinib inhibited cell growth in a dose-dependent manner (Figure 6(d)).
Figure 6.
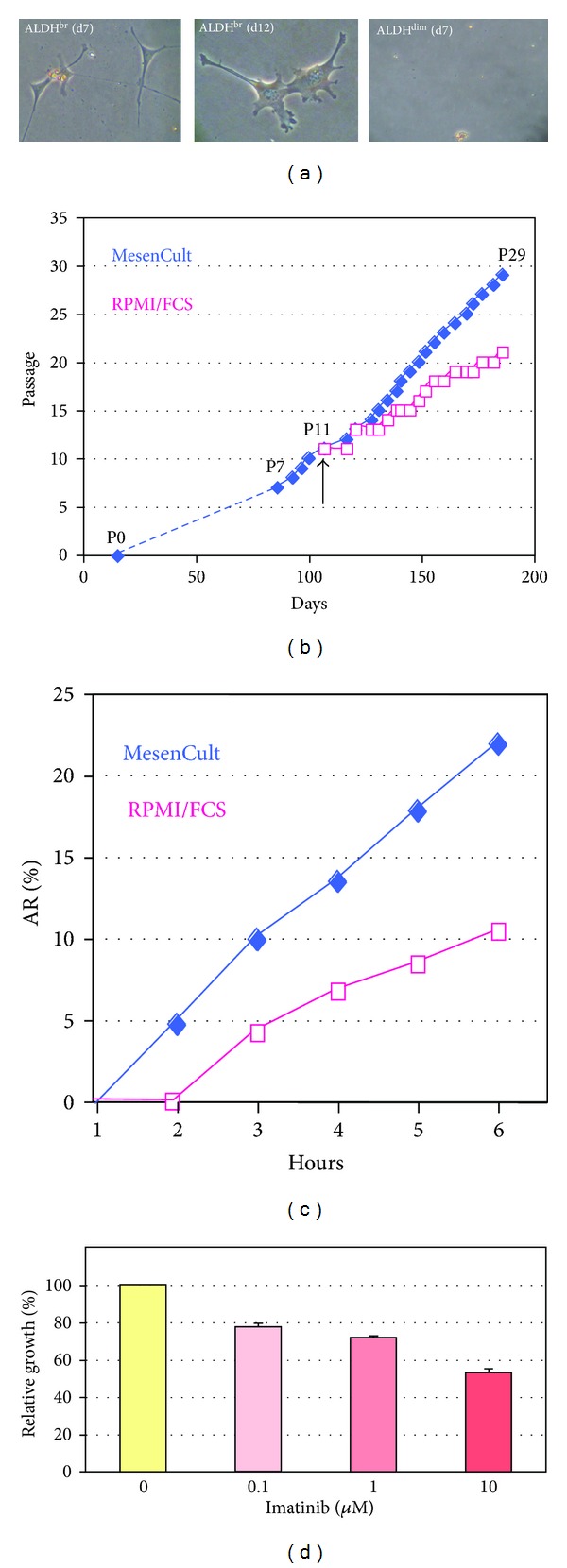
Ex vivo culture-expansion of ALDHbr atrial cells (a). Photomicrographs of ALDHbr cells cultured for 7 and 12 days in MesenCult medium. ALDHdim cells were dead by day 7 (b). Growth curves of cells in different growth media. Cells were cultured in MesenCult medium up to P10 (arrow). From P11 on, MesenCult was either maintained or replaced by RPMI/FCS. Symbols (squares and diamonds) indicate cell passage (no data available from P1 to P7; dotted line) (c). AlamarBlue assay on cells (P40) cultured in either MesenCult or RPMI/FCS (AR%: percent AlamarBlue reduction) (d). Dose-response study of Imatinib with respect to growth inhibition of expanded ALDHbr cells (P25).
3.6. Immunophenotype of Culture-Expanded ALDHbr Cells
The marker profile of expanded ALDHbr cells (P11-13) was analyzed by flow cytometry (Figure 7). Cells expanded in MesenCult medium stained positive for Sca-1, CD29, CD44, CD105, CD106, and, in part, CD146 and CD14. They stained negative for CD45, CD11b, CD31, and CD133. To assess whether the culture medium affected marker expression, cells were also grown in RPMI/FCS. The immunophenotype of these cells was similar to those grown in MesenCult, although larger cell subsets stained positive for c-kit, CD140b (PDGFRB) and NG2 chondroitin sulfate proteoglycan (Figures 7(b) and 7(c)). NG2 essentially colocalized with CD140b and CD146 (Figure 7(d)). c-kit expression was demonstrated by immunocytochemistry (Figure 7(e)).
Figure 7.
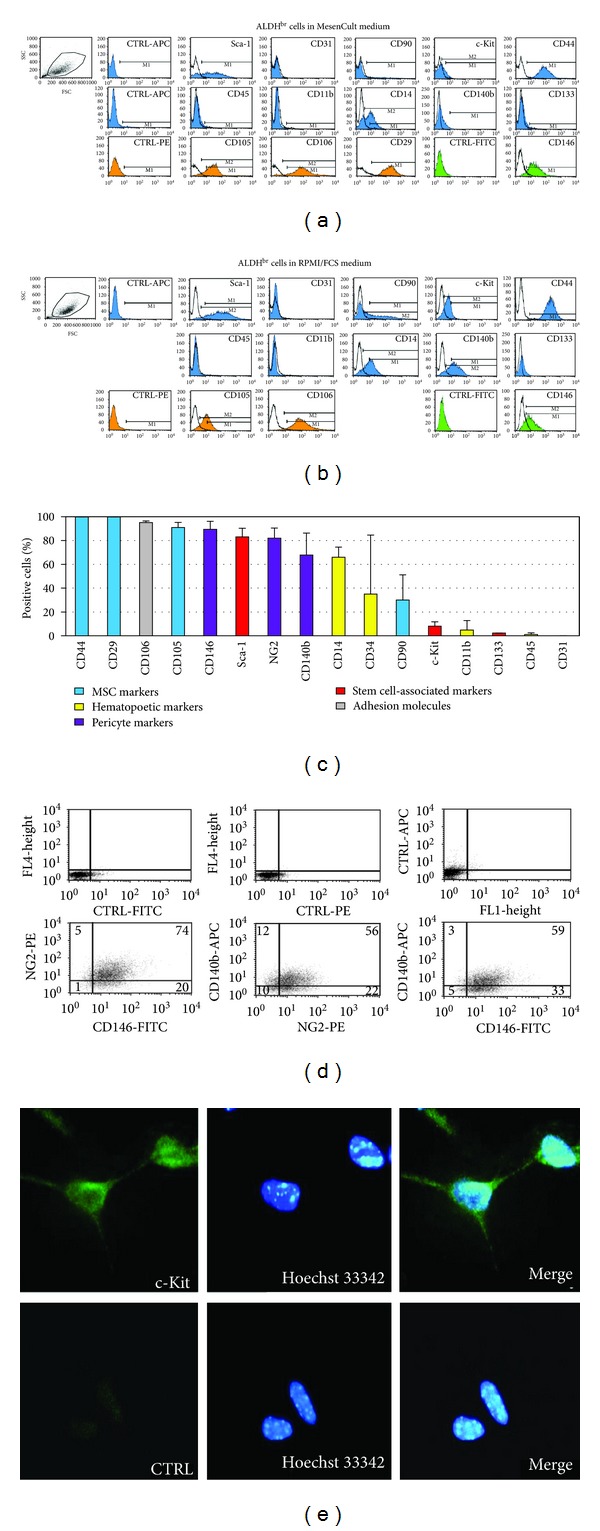
(a)/(b) Flow cytometric analysis of ALDHbr atrial cells expanded in MesenCult (P11) or RPMI/FCS medium (P13). (c) Percentages of cells (mean, SD) expressing the indicated markers (data from 3 separate experiments in MesenCult). (d) Two-color analysis of pericyte/perivascular markers (NG2, CD146, and CD140b; numbers are cell percentages). (e) c-kit immunostaining of expanded ALDHbr cells. Nuclear staining with Hoechst 33342 (CTRL, secondary Ab only as a control).
3.7. Expanded Cells Express Sarcomeric α-Actinin and Differentiate along Mesenchymal Lineages
Culture-expanded ALDHbr cells could be induced to differentiate along adipogenic, osteogenic, and chondrogenic lineages in appropriate culture media, as evidence by staining with Oil red-O, Alizarin red, Alcian blue (not shown), and type II collagen immunostaining, respectively (Figures 8(a), 8(c) and 8(g)). Cells cultured in MesenCult stained positive for sarcomeric α-actinin (Figure 8(e)) but negative for α-SMA and von Willebrand factor (not shown).
Figure 8.
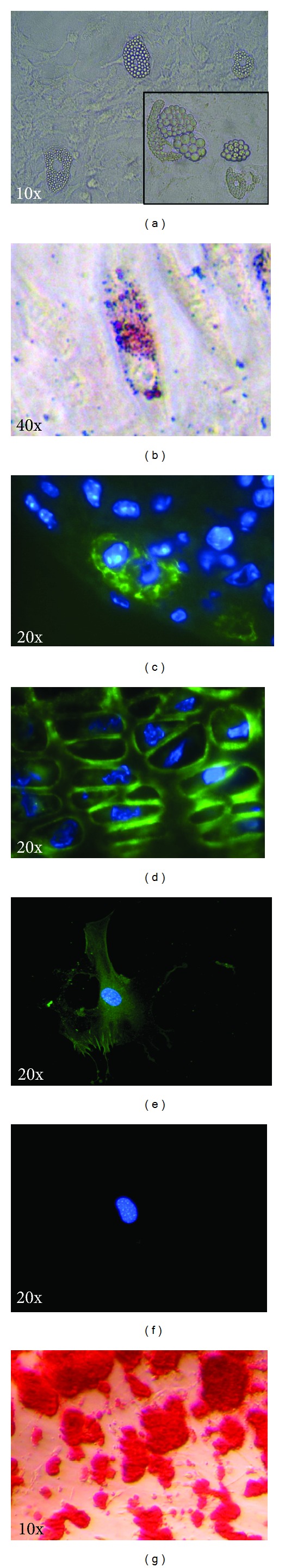
Differentiation potential of expanded ALDHbr cells. (a) Photomicrograph showing intracellular fat droplets in cells cultured in adipogenic medium. (b) Positive Oil Red-O staining of intracellular droplets. (c) Positive type-II collagen immunostaining (green) in cells cultured in chondrogenic medium. (d) Type-II collagen immunostaining of leg cartilage, as a positive control. (e) Positive sarcomeric α-actinin immunostaining (green) of an ALDHbr cell (the protein does not show clear sarcomeric distribution). (f) Corresponding negative control (secondary Ab only). (g) Positive Alizarin Red staining of expanded ALDHbr cells cultured in osteogenic medium.
4. Discussion
Growing evidence suggests high ALDH activity may be a common feature shared by stem and progenitor cells across normal tissues, as well as in cancer. Human UCB and BM cells possessing high ALDH activity have shown angiogenic activity in preclinical studies [30–32] and have been used safely in phase I clinical trials in patients with ischemic cardiovascular disease [33–35].
We characterized, for the first time, ALDHbr cells isolated from the heart. Approximately one percent of all nonmyocytic cells present in the young adult mouse heart were ALDHbr, as measured by Aldefluor stain (Figure 1). The neonatal heart contained a similar number of ALDHbr cells, although with a higher proportion of cells exhibiting very high ALDH activity (ALDHvery-br) compared to the young adult heart. The frequency of ALDHbr cells in the population isolated from the atria was approximately 10-fold higher than in cells derived from ventricles (Figure 2). This observation has analogies with previous reports on increased numbers of putative stem/progenitor cells, such as DNA-label retaining cells in rodents [36, 37] and c-kit+ cells in human [38], in atria relative to ventricles. In the present study, ALDHbr cells isolated from atria and ventricles exhibited slightly different side scatter (SSC) properties. Most atrial ALDHbr cells showed SSClo properties but a small subset possessed intermediate SSC properties. In contrast, ventricular ALDHbr cells uniformly exhibited SSClo properties. The original characterizations of ALDHbr cells from human UCB revealed that the ALDHbrSSClo population was enriched 50- to 100-fold for primitive hematopoietic progenitors [11, 15]. In several studies, SSClo properties therefore were associated with the traditional definition of ALDHbr hematopoietic cells. However, other studies focused on ALDHbrLin− (lineage-depleted) cells [13, 14, 16]. We defined ALDHbr cells based solely on ALDH activity, regardless of SSC and lineage properties. This methodological approach affected ALDHbr cell counts only marginally, given the predominant ALDHbrSSClo phenotype. Nevertheless, the slightly different SSC properties of atrial and ventricular cells may suggest differences in the cellular composition of the ALDHbr population depending on the cardiac chamber of origin. This issue remains to be addressed in future studies.
Freshly isolated ALDHbr cells exhibited a heterogeneous phenotype, reflecting a mixture of different cell types (Figures 3 and 4). This population was significantly enriched for cells expressing Sca-1, the early hematopoietic and EPC marker CD34, the MSC marker CD90, the hyaluronic acid receptor CD44, and vascular cell adhesion molecule-1 (CD106). CD44 and CD106 were the most highly enriched markers in the ALDHbr versus ALDHdim population (34.3 and 24.1 fold-increases, resp.). Previous studies associated CD44 and CD106 expression with MSCs [39]. However, a recent study showed that freshly isolated murine and human BM stromal cells did not express CD44 and CD106, but they could acquire expression of these antigens in vitro [40]. CD44+CD24− cells have been proposed to represent cancer stem cells in breast cancer [41], as well as in head and neck squamous cell carcinoma (the latter includes an ALDH1+CD44+CD24− stem cell subset) [42]. In the present study, ALDHvery-br cells were highly enriched for Sca-1 and CD90, but only moderately so for CD34, possibly reflecting a predominant MSC-like subpopulation with fewer hematopoietic/vascular progenitors compared to ALDHbr cells with lower levels of ALDH activity. The ALDHbr population was significantly depleted for common leukocyte antigen (CD45), the endothelial marker CD31, and CD38 (an antigen expressed by the differentiating progeny of CD34+CD38− hematopoietic/vascular progenitors). ALDHvery-br cells were further depleted for lineage differentiation markers compared to ALDHbr cells. Thus, the ALDHbr population was enriched for MSC-like progenitors and depleted for differentiating hematopoietic and endothelial cells.
Purified ALDHbr atrial cells could be expanded ex vivo, whereas their ALDHdim counterparts could not. Because the atrial population contained more ALDHbr cells than the ventricular population, atrial cells were used for the purification of ALDHbr cells in order to shorten the sorting procedure, and therefore to attenuate cell damage. Purified ALDHbr cells formed a few plastic-adherent colonies in each culture well. Each colony apparently originated from a single cell, providing observational evidence of clonal expansion, even though this was not firmly established using a clonogenic assay. The mechanism responsible for the selective growth of ALDHbr cells in culture remains unclear. To initially address this question, we determined the respective contributions of selected ALDH isozymes to ALDH activity, measured by Aldefluor stain. Fifteen ALDH isozymes have been described in mice [43] and 19 in human [9, 44]. They play different biological roles that vary among cell types and species. ALDH1A1, the ALDH isozyme most highly overexpressed in human BM ALDHbr versus ALDHdim cells [44], was believed to be responsible for ALDH activity, measured by Aldefluor stain, in BM and other tissues. However, recent findings in genetically modified mice demonstrated that ALDH1A1 was dispensable for stem cell function in the mouse hematopoietic and nervous systems [45]. ALDH1 was identified as a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome in breast cancer [28]. In a distinct study, shRNA knock-down data indicated that ALDH1A3, not ALDH1A1, correlated best with ALDH activity in breast cancer stem cells [46]. ALDH1A1 and ALDH3A1 were highly expressed in nonsmall cell lung cancer, and knock-down of these ALDH isoforms were associated with in vitro functional changes in the proliferation and motility of these cells [47]. Human ALDH1A1, ALDH1A2, ALDH1A3, and ALDH8A1 function in retinoic acid cell signaling via retinoic acid production by oxidation of all-trans-retinal and 9-cis-retinal [6, 46]. ALDH2, a mitochondrial isozyme that mediates both the detoxification of reactive aldehydes and the bioactivation of nitroglycerin to nitric oxide, mediates cytoprotection in the heart [48]. Recent data indicated that the aldehyde-oxidizing activity of mouse HSCs, measured by Aldefluor stain, was due to the ALDH2 isozyme and correlated with protection against acetaldehyde toxicity [49]. Mice deficient in both Fanconi anemia pathway-mediated DNA repair and acetaldehyde detoxification showed more than a 600-fold reduction in the HSC pool.
Based on these reports, we measured the expression of ALDH1A1, ALDH1A2, ALDH1A3, and ALDH2 gene transcripts in purified ALDHdim, ALDHbr, and ALDHvery-br cardiac-derived cells (Figure 5). Expression of ALDH1A1 was lowest in ALDHdim cells, intermediate in ALDHbr cells, and highest in ALDHvery-br cells (2−ΔΔCt = 3.5 using ALDHdim as a reference population). By contrast, expression of ALDH1A2 was lowest in ALDHbr cells, intermediate in ALDHdim cells, and highest in ALDHvery-br cells (2−ΔΔCt = 4.9 versus ALDHdim). Expression of ALDH1A3 and ALDH2 was detectable in ALDHbr and ALDHvery-br cells, albeit at lower levels compared to ALDH1A1 and ALDH1A2, but not in ALDHdim cells. Thus, ALDH1A1 correlated best with ALDH activity; however, contributory roles of ALDH1A2, ALDH1A3, and ALDH2 to ALDH activity are possible based on our data. While the present analysis included the four ALDH isoforms most often associated with ALDH activity in previous reports [44–49], it should be completed to include the remaining ALDH isoforms. In addition, shRNA knock-down experiments are needed for identifying the ALDH isozyme(s) responsible for ALDH activity in these cells.
Recent data suggested BM and UCB ALDHbr cells might repair ischemic tissues in vivo by releasing angiogenic factors [30–32]. A comparison of gene expression profiles by ALDHbr and ALDHdim human BM cells identified three angiogenic factors (endoglin/CD105, ephrin B4, and angiopoietin-1) as the most highly overexpressed factors in ALDHbr versus ALDHdim cells (fold-changes: 66.9, 64.6, and 7.0, resp.) [44]. We therefore measured mRNA expression of these angiogenic genes in cardiac-derived cells but found no correlation with ALDH activity (besides the preliminary observation that angiopoietin-1 transcripts were detectable in ALDHbr and ALDHvery-br cells but not in ALDHdim cells). The angiogenic potential of cardiac-derived ALDHbr cells remains to be addressed in future studies. Previous reports showed that ALDHbr MSCs from human UCB were more responsive to hypoxia than their ALDHdim counterparts, with upregulation of Flt-1, CXCR4, and angiopoietin-2 [50]. The same group reported that ALDHdim EPCs, but not ALDHbr EPCs, from human UCB upregulated hypoxia-inducible factor proteins as well as VEGF, CXCR4, and GLUT-1 mRNAs under hypoxic conditions [22]. The introduction of ALDHdim EPCs significantly reduced ischemic tissue damage in a mouse flap model, whereas ALDHbr EPCs were ineffective. These findings suggested varying angiogenic activities of ALDHbr cells depending on the cell type studied.
In the present study, plastic-adherent cell clusters grew rapidly and could be passaged more than 40 times (later passages were not tested). Growth rates in complete MesenCult medium were higher than in RPMI/FCS (Figure 6). Imatinib, which selectively inhibits PDGFRB (CD140b) and c-kit tyrosine kinases with a similar potency [51], reduced cell proliferation in a dose-dependent manner. Expression of PDGFRB and c-kit was detected in subsets of expanded cells. The effect of Imatinib might reflect a role for PDGFRB and/or c-kit in the proliferation of these cells. Previous studies showed that ALDHbr MSCs from UCB proliferated more than their ALDHdim counterparts [50], whereas ALDHdim EPCs proliferated more than ALDHbr EPCs [22]. Proliferative properties correlated with angiogenic activities in these populations. Again, the association of ALDH and proliferation may vary among cell types. The immunophenotype of expanded ALDHbr cardiac cells resembled that of freshly isolated ALDHbr cells (Figure 7). Positive stains for pericytic markers (NG2, CD146, and CD140b) were consistent with a MSC-like phenotype. It has been shown that perivascular cells of arterioles and capillaries from many tissues express pericytic markers and show MSC-like and angiogenic features [52–54]. In the present study, expanded cells stained positive for sarcomeric α-actinin, a cardiac marker, and exhibited differentiation potential along multiple mesenchymal lineages. Previous data showed that aldehyde dehydrogenase activity did not increase the chondrogenic potential of human adipose-derived adult stem cells [55]. Recent reports showed that human BM contained ALDHbr multipotential mesenchymal progenitor cells (MPCs), besides ALDHbr HSCs and EPCs [12, 56]. These MPCs strongly expressed ALDH, expressed embryonic markers not present in mesenchymal cells, and could be differentiated to microvascular endothelial cells and mesenchymal cells. The latter lacked strong ALDH expression and could not be induced to back differentiate into MPCs.
We observed a loss of ALDHbr cells with cell passage (e.g., only 11% of cells at passage 7 were still ALDHbr). Whether this loss of ALDHbr cells with time reflected cellular adaptation to ex vivo conditions or the replacement of early progenitors by a progeny of late progenitor and precursor cells remains unclear. The observation that growth rates and the immunophenotype remained stable for more than 30 and 10 passages, respectively, seemed not to support cell differentiation. ALDH as a stem/progenitor cell marker has chiefly been used in freshly isolated cells and tissues, as opposed to expanded cell populations. The significance of ALDH in culture-expanded cells is unclear, as is its contribution to stem cell fate. The aforementioned observation that the mesenchymal cell progeny of ALDHbr human BM cells lacked strong ALDH expression and could not be induced to back differentiate into MPCs suggested a parallel loss of ALDH expression and cell potency [56].
In conclusion, cardiac-derived ALDHbr cells are progenitor cells with MSC-like phenotype and function and with superior ex vivo growth characteristics compared to their ALDHdim counterparts. Further studies are needed for assessing the regenerative potential of these cells in animal models of heart disease.
Acknowledgments
This work is supported by the Swiss Heart Foundation (Berne, Switzerland), the Fondation Vaudoise de Cardiologie (Lausanne, Switzerland), the Cecilia Augusta Foundation (Lugano, Switzerland), the METIS Foundation Sergio Mantegazza (Lugano, Switzerland), and the “Fondazione per la ricerca sulla trasfusione e sui trapianti” (Lugano, Switzerland).
References
- 1.Oh H, Bradfute SB, Gallardo TD, et al. Cardiac progenitor cells from adult myocardium: homing, differentiation, and fusion after infarction. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(21):12313–12318. doi: 10.1073/pnas.2132126100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Beltrami AP, Barlucchi L, Torella D, et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell. 2003;114(6):763–776. doi: 10.1016/s0092-8674(03)00687-1. [DOI] [PubMed] [Google Scholar]
- 3.Laugwitz KL, Moretti A, Lam J, et al. Postnatal isl1+ cardioblasts enter fully differentiated cardiomyocyte lineages. Nature. 2005;433(7026):647–653. doi: 10.1038/nature03215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Laugwitz KL, Moretti A, Lam J, et al. Erratum: Postnatal isl1+ cardioblasts enter fully differentiated cardiomyocyte lineages. Nature. 2007;446(7138):p. 934. doi: 10.1038/nature03215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cai J, Weiss ML, Rao MS. In search of “stemness”. Experimental Hematology. 2004;32(7):585–598. doi: 10.1016/j.exphem.2004.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Balber AE. Concise review: aldehyde dehydrogenase bright stem and progenitor cell populations from normal tissues: characteristics, activities, and emerging uses in regenerative medicine. Stem Cells. 2011;29(4):570–575. doi: 10.1002/stem.613. [DOI] [PubMed] [Google Scholar]
- 7.Laird DJ, De Tomaso AW, Weissman IL. Stem cells are units of natural selection in a colonial ascidian. Cell. 2005;123(7):1351–1360. doi: 10.1016/j.cell.2005.10.026. [DOI] [PubMed] [Google Scholar]
- 8.Laird DJ, De Tomaso AW, Weissman IL. Erratum: Stem cells are units of natural selection in a colonial ascidian. Cell. 2006;124(3):647–648. doi: 10.1016/j.cell.2005.10.026. [DOI] [PubMed] [Google Scholar]
- 9.Marchitti SA, Brocker C, Stagos D, Vasiliou V. Non-P450 aldehyde oxidizing enzymes: the aldehyde dehydrogenase superfamily. Expert Opinion on Drug Metabolism and Toxicology. 2008;4(6):697–720. doi: 10.1517/17425250802102627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kastan MB, Schlaffer E, Russo JE, Colvin OM, Civin CI, Hilton J. Direct demonstration of elevated aldehyde dehydrogenase in human hematopoietic progenitor cells. Blood. 1990;75(10):1947–1950. [PubMed] [Google Scholar]
- 11.Storms RW, Trujillo AP, Springer JB, et al. Isolation of primitive human hematopoietic progenitors on the basis of aldehyde dehydrogenase activity. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(16):9118–9123. doi: 10.1073/pnas.96.16.9118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gentry T, Foster S, Winstead L, Deibert E, Fiordalisi M, Balber A. Simultaneous isolation of human BM hematopoietic, endothelial and mesenchymal progenitor cells by flow sorting based on aldehyde dehydrogenase activity: implications for cell therapy. Cytotherapy. 2007;9(3):259–274. doi: 10.1080/14653240701218516. [DOI] [PubMed] [Google Scholar]
- 13.Chute JP, Muramoto GG, Whitesides J, et al. Inhibition of aldehyde dehydrogenase and retinoid signaling induces the expansion of human hematopoietic stem cells. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(31):11707–11712. doi: 10.1073/pnas.0603806103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hess DA, Meyerrose TE, Wirthlin L, et al. Functional characterization of highly purified human hematopoietic repopulating cells isolated according to aldehyde dehydrogenase activity. Blood. 2004;104(6):1648–1655. doi: 10.1182/blood-2004-02-0448. [DOI] [PubMed] [Google Scholar]
- 15.Storms RW, Green PD, Safford KM, et al. Distinct hematopoietic progenitor compartments are delineated by the expression of aldehyde dehydrogenase and CD34. Blood. 2005;106(1):95–102. doi: 10.1182/blood-2004-09-3652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hess DA, Wirthlin L, Craft TP, et al. Selection based on CD133 and high aldehyde dehydrogenase activity isolates long-term reconstituting human hematopoietic stem cells. Blood. 2006;107(5):2162–2169. doi: 10.1182/blood-2005-06-2284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pearce DJ, Bonnet D. The combined use of Hoechst efflux ability and aldehyde dehydrogenase activity to identify murine and human hematopoietic stem cells. Experimental Hematology. 2007;35(9):1437–1446. doi: 10.1016/j.exphem.2007.06.002. [DOI] [PubMed] [Google Scholar]
- 18.Christ O, Lucke K, Imren S, et al. Improved purification of hematopoietic stem cells based on their elevated aldehyde dehydrogenase activity. Haematologica. 2007;92(9):1165–1172. doi: 10.3324/haematol.11366. [DOI] [PubMed] [Google Scholar]
- 19.Gentry T, Deibert E, Foster SJ, Haley R, Kurtzberg J, Balber AE. Isolation of early hematopoietic cells, including megakaryocyte progenitors, in the ALDH-bright cell population of cryopreserved, banked UC blood. Cytotherapy. 2007;9(6):569–576. doi: 10.1080/14653240701466347. [DOI] [PubMed] [Google Scholar]
- 20.Fallon P, Gentry T, Balber AE, et al. Mobilized peripheral blood SSCloALDHbr cells have the phenotypic and functional properties of primitive haematopoietic cells and their number correlates with engraftment following autologous transplantation. British Journal of Haematology. 2003;122(1):99–108. doi: 10.1046/j.1365-2141.2003.04357.x. [DOI] [PubMed] [Google Scholar]
- 21.Povsic TJ, Zavodni KL, Kelly FL, et al. Circulating progenitor cells can be reliably identified on the basis of aldehyde dehydrogenase activity. Journal of the American College of Cardiology. 2007;50(23):2243–2248. doi: 10.1016/j.jacc.2007.08.033. [DOI] [PubMed] [Google Scholar]
- 22.Nagano M, Yamashita T, Hamada H, et al. Identification of functional endothelial progenitor cells suitable for the treatment of ischemic tissue using human umbilical cord blood. Blood. 2007;110(1):151–160. doi: 10.1182/blood-2006-10-047092. [DOI] [PubMed] [Google Scholar]
- 23.Corti S, Locatelli F, Papadimitriou D, et al. Transplanted ALDHhiSSClo neural stem cells generate motor neurons and delay disease progression of nmd mice, an animal model of SMARD1. Human Molecular Genetics. 2006;15(2):167–187. doi: 10.1093/hmg/ddi446. [DOI] [PubMed] [Google Scholar]
- 24.Corti S, Nizzardo M, Nardini M, et al. Neural stem cell transplantation can ameliorate the phenotype of a mouse model of spinal muscular atrophy. Journal of Clinical Investigation. 2008;118(10):3316–3330. doi: 10.1172/JCI35432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Vauchez K, Marolleau JP, Schmid M, et al. Aldehyde dehydrogenase activity identifies a population of human skeletal muscle cells with high myogenic capacities. Molecular Therapy. 2009;17(11):1948–1958. doi: 10.1038/mt.2009.204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jean E, Laoudj-Chenivesse D, Notarnicola C, et al. Aldehyde dehydrogenase activity promotes survival of human muscle precursor cells. Journal of Cellular and Molecular Medicine. 2011;15(1):119–133. doi: 10.1111/j.1582-4934.2009.00942.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Huang EH, Hynes MJ, Zhang T, et al. Aldehyde dehydrogenase 1 is a marker for normal and malignant human colonic stem cells (SC) and tracks SC overpopulation during colon tumorigenesis. Cancer Research. 2009;69(8):3382–3389. doi: 10.1158/0008-5472.CAN-08-4418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ginestier C, Hur MH, Charafe-Jauffret E, et al. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell. 2007;1(5):555–567. doi: 10.1016/j.stem.2007.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Marcato P, Dean CA, Pan D, et al. Aldehyde dehydrogenase activity of breast cancer stem cells is primarily due to isoform ALDH1A3 and its expression is predictive of metastasis. Stem Cells. 2011;29(1):32–45. doi: 10.1002/stem.563. [DOI] [PubMed] [Google Scholar]
- 30.Sondergaard CS, Hess DA, Maxwell DJ, et al. Human cord blood progenitors with high aldehyde dehydrogenase activity improve vascular density in a model of acute myocardial infarction. Journal of Translational Medicine. 2010;8, article 24 doi: 10.1186/1479-5876-8-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Capoccia BJ, Robson DL, Levac KD, et al. Revascularization of ischemic limbs after transplantation of human bone marrow cells with high aldehyde dehydrogenase activity. Blood. 2009;113(21):5340–5351. doi: 10.1182/blood-2008-04-154567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bell GI, Broughton HC, Levac KD, Allan DA, Xenocostas A, Hess DA. Transplanted human bone marrow progenitor subtypes stimulate endogenous islet regeneration and revascularization. Stem Cells and Development. 2012;21(1):97–109. doi: 10.1089/scd.2010.0583. [DOI] [PubMed] [Google Scholar]
- 33.Perin EC, Silva G, Gahremanpour A, et al. A randomized, controlled study of autologous therapy with bone marrow-derived aldehyde dehydrogenase bright cells in patients with critical limb ischemia. Catheterization and Cardiovascular Interventions. 2011;78(7):1060–1067. doi: 10.1002/ccd.23066. [DOI] [PubMed] [Google Scholar]
- 34.Perin EC, Silva GV, Zheng Y, et al. First randomized, controlled double-blind trial of transendocardial injection of autologous aldehyde dehydrogenase-bright stem cells. American Heart Journal. 2011;161(6):1078–11087. doi: 10.1016/j.ahj.2011.11.020. [DOI] [PubMed] [Google Scholar]
- 35.Perin EC, Silva GV, Zheng Y, et al. Randomized, double-blind pilot study of transendocardial injection of autologous aldehyde dehydrogenase-bright stem cells in patients with ischemic heart failure. American Heart Journal. 2012;163(3):415–421. doi: 10.1016/j.ahj.2011.11.020. [DOI] [PubMed] [Google Scholar]
- 36.Urbanek K, Cesselli D, Rota M, et al. Stem cell niches in the adult mouse heart. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(24):9226–9231. doi: 10.1073/pnas.0600635103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Meinhardt A, Spicher A, Roehrich ME, Glauche I, Vogt P, Vassalli G. Immunohistochemical and flow cytometric analysis of long-term label-retaining cells in the adult heart. Stem Cells and Development. 2011;20(2):211–222. doi: 10.1089/scd.2009.0203. [DOI] [PubMed] [Google Scholar]
- 38.Castaldo C, Di Meglio F, Nurzynska D, et al. CD117-positive cells in adult human heart are localized in the subepicardium, and their activation is associated with laminin-1 and α6 integrin expression. Stem Cells. 2008;26(7):1723–1731. doi: 10.1634/stemcells.2007-0732. [DOI] [PubMed] [Google Scholar]
- 39.Baddoo M, Hill K, Wilkinson R, et al. Characterization of mesenchymal stem cells isolated from murine bone marrow by negative selection. Journal of Cellular Biochemistry. 2003;89(6):1235–1249. doi: 10.1002/jcb.10594. [DOI] [PubMed] [Google Scholar]
- 40.Qian H, Le Blanc K, Sigvardsson M. Primary mesenchymal stem and progenitor cells from bone marrow lack expression of CD44 protein. The Journal of Biological Chemistry. 2012;287(31):25795–25807. doi: 10.1074/jbc.M112.339622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(7):3983–3988. doi: 10.1073/pnas.0530291100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chen C, Wei Y, Hummel M, et al. Evidence for epithelial-mesenchymal transition in cancer stem cells of head and neck squamous cell carcinoma. PLoS ONE. 2011;6(1) doi: 10.1371/journal.pone.0016466.e16466 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Alnouti Y, Klaassen CD. Tissue distribution, ontogeny, and regulation of aldehyde dehydrogenase (Aldh) enzymes mRNA by prototypical microsomal enzyme inducers in mice. Toxicological Sciences. 2008;101(1):51–64. doi: 10.1093/toxsci/kfm280. [DOI] [PubMed] [Google Scholar]
- 44.Storrie White H, Smith L, Gentry T, Balber AE. Mechanisms of action of human aldehyde dehydrogenase bright cells in therapy of cardiovascular diseases: expression analysis of angiogenic factors and aldehyde dehydrogenase isozymes. Journal of Stem Cell Research. In press, http://dx.doi.org/10.4172/2157-7633.S1-001. [Google Scholar]
- 45.Levi BP, Yilmaz ÖH, Duester G, Morrison SJ. Aldehyde dehydrogenase 1a1 is dispensable for stem cell function in the mouse hematopoietic and nervous systems. Blood. 2009;113(8):1670–1680. doi: 10.1182/blood-2008-05-156752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Marcato P, Dean CA, Giacomantonio CA, Lee PWK. Aldehyde dehydrogenase its role as a cancer stem cell marker comes down to the specific isoform. Cell Cycle. 2011;10(9):1378–1384. doi: 10.4161/cc.10.9.15486. [DOI] [PubMed] [Google Scholar]
- 47.Moreb JS, Baker HV, Chang LJ, et al. ALDH isozymes downregulation affects cell growth, cell motility and gene expression in lung cancer cells. Molecular Cancer. 2008;7, article 87 doi: 10.1186/1476-4598-7-87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Budas GR, Disatnik MH, Mochly-Rosen D. Aldehyde dehydrogenase 2 in cardiac protection: a new therapeutic target? Trends in Cardiovascular Medicine. 2009;19(5):158–164. doi: 10.1016/j.tcm.2009.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Garaycoechea JI, Crossan GP, Langevin F, Daly M, Arends MJ, Patel KJ. Genotoxic consequences of endogenous aldehydes on mouse haematopoietic stem cell function. Nature. 2012;489(7417):571–575. doi: 10.1038/nature11368. [DOI] [PubMed] [Google Scholar]
- 50.Nagano M, Kimura K, Yamashita T, et al. Hypoxia responsive mesenchymal stem cells derived from human umbilical cord blood are effective for bone repair. Stem Cells and Development. 2010;19(8):1195–1210. doi: 10.1089/scd.2009.0447. [DOI] [PubMed] [Google Scholar]
- 51.Buchdunger E, Cioffi CL, Law N, et al. Abl protein-tyrosine kinase inhibitor STI571 inhibits in vitro signal transduction mediated by c-Kit and platelet-derived growth factor receptors. Journal of Pharmacology and Experimental Therapeutics. 2000;295(1):139–145. [PubMed] [Google Scholar]
- 52.Crisan M, Yap S, Casteilla L, et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell. 2008;3(3):301–313. doi: 10.1016/j.stem.2008.07.003. [DOI] [PubMed] [Google Scholar]
- 53.Campagnolo P, Cesselli D, Al Haj Zen A, et al. Human adult vena saphena contains perivascular progenitor cells endowed with clonogenic and proangiogenic potential. Circulation. 2010;121(15):1735–1745. doi: 10.1161/CIRCULATIONAHA.109.899252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Katare R, Riu F, Mitchell K, et al. Transplantation of human pericyte progenitor cells improves the repair of infarcted heart through activation of an angiogenic program involving micro-RNA-132. Circulation Research. 2011;109(8):894–906. doi: 10.1161/CIRCRESAHA.111.251546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Estes BT, Wu AW, Storms RW, Guilak F. Extended passaging, but not aldehyde dehydrogenase activity, increases the chondrogenic potential of human adipose-derived adult stem cells. Journal of Cellular Physiology. 2006;209(3):987–995. doi: 10.1002/jcp.20808. [DOI] [PubMed] [Google Scholar]
- 56.Petrini M, Pacini S, Trombi L, et al. Identification and purification of mesodermal progenitor cells from human adult bone marrow. Stem Cells and Development. 2009;18(6):857–866. doi: 10.1089/scd.2008.0291. [DOI] [PMC free article] [PubMed] [Google Scholar]