

Abstract
It is well-known that the onset of puberty is associated with changes in mood as well as cognition. Stress can have an impact on these outcomes, which in many cases, can be more influential in females, suggesting that gender differences exist. The adolescent period is a vulnerable time for the onset of certain psychopathologies, including anxiety disorders, depression and eating disorders, which are also more prevalent in females. One factor which may contribute to stress-triggered anxiety at puberty is the GABAA receptor (GABAR), which is known to play a pivotal role in anxiety. Expression of α4βδ GABARs increases on the dendrites of CA1 pyramidal cells at the onset of puberty in the hippocampus, part of the limbic circuitry which governs emotion. This receptor is a sensitive target for the stress steroid THP (3α-OH-5[α]β-pregnan-20-one), which paradoxically reduces inhibition and increases anxiety during the pubertal period (~PND 35–44) of female mice in contrast to its usual effect to enhance inhibition and reduce anxiety. Spatial learning and synaptic plasticity are also adversely impacted at puberty, likely a result of increased expression of α4βδ GABARs on the dendritic spines of CA1 hippocampal pyramidal cells, which are essential for consolidation of memory. This review will focus on the role of these receptors in mediating behavioral changes at puberty. Stress-mediated changes in mood and cognition in early adolescence may have relevance for the expression of psychopathologies in adulthood.
Keywords: Puberty, GABA-A receptor, alpha4, delta, anxiety, cognition
1.1 Introduction
It is well known that adolescence is a period frequently associated with behavioral and cognitive changes in humans, which can include mood swings (Buchanan et al., 1992), increased/altered response to stress (Modesti et al., 1994; Susman et al., 1988), risk-seeking behavior (Liang et al., 1995), altered social interaction, and, in some cases, decreases in CNS plasticity and cognition (Johnson and Newport, 1989; McGivern et al., 2002). It is also recognized as a particularly vulnerable time for the onset of certain psychiatric disorders, including generalized anxiety disorder (Reardon et al., 2009) and depression (Heim et al., 2004), which are more prevalent in females (Dorn and Chrousos, 1997), as well as a period when stress can influence the eventual development of some psychiatric disorders, such as schizophrenia, that occur later in adulthood (Corcoran et al., 2012; Dahl, 2004). Some studies have noted developmental milestones associated with chronological age which impact upon some of these cognitive and behavioral outcomes (Vetter-O'Hagen and Spear, 2012). However, the onset of puberty is associated with an array of hormonal events which converge to produce viable reproductive function. These same hormonal events may also contribute to some of the psychological and cognitive changes associated with puberty which are not directly linked to reproductive function.
Stress is defined as the body’s reaction to environmental demands which elicit physical, mental or emotional responses, in most cases correlated with changes in heart rate and/or blood pressure. Most studies typically use performance stress (oral presentations, mental arithmetic, mirror tracing, etc.) for human stress evaluations and restraint stress or CO2 inhalation for rodent studies. Stress steroids are steroids released as part of the reaction to stress. The most well-known stress hormones include epinephrine, norepinephrine and corticosterone; Animal studies have shown that corticosterone, in particular, produces profound effects on mood and cognition via acute and chronic effects on neuronal function and viability (Davidson and McEwen, 2012; McEwen, 2007; McEwen et al., 2012). However, this article will focus on the role of THP ([allo]pregnanolone, 3α-OH-5[α]β-pregnan-20-one) in mediating some of the changes in mood and cognition observed during the pubertal period. THP is also a steroid released by stress (Girdler et al., 2006; Higashi et al., 2005; Purdy et al., 1991), by the adrenal and CNS in both humans and rodents, which functions to modulate inhibition via direct effects on the GABAA receptor (GABAR), a receptor shown to play a pivotal role in anxiety (Rudolph et al., 1999; Trincavelli et al., 2012) in mice and humans.
1.2 Anxiety and adolescence
One of the most prevalent types of psychopathologies in human adolescents is anxiety (Costello et al., 2003). In fact, anxiety disorders with a lifetime course are most likely to begin at the time of puberty. In one study the median age of onset was 11 years (Kessler et al., 2005), consistent with pubertal onset, with more than 50% having an onset by age 14, during the pubertal period. There are also gender differences in psychopathology during adolescence, with girls more likely to develop an anxiety disorder (Hayward and Sanborn, 2002), for whom the most common subtypes reported are social anxiety and panic disorder (Costello et al., 2003; Zgourides and Warren, 1988). Both the prevalence and intensity of panic attacks have been correlated with pubertal status in girls (Hayward et al., 1992; Leen-Feldner et al., 2007).
1.3 Stress and adolescence
Response to a stressful experience is increased at the onset of puberty (Modesti et al., 2006; Sumter et al., 2010; Susman et al., 1988) in humans, where performance on a video game was shown to provoke increases in blood pressure, and the anticipatory stress of public speaking increased cortisol responses in adolescents to a greater degree than in younger children. Puberty is also widely regarded as the onset of gender differences in negative affect as a response to stress (Dorn and Chrousos, 1997; Ordaz and Luna, 2012). Many studies in humans have reported greater negative mood in females in early adolescence as a response to performance or psychosocial stressors (Garber et al., 2002; Ge et al., 2010), which is a trait that can extend into adulthood (Kelly et al., 2008). In studies investigating the autonomic changes triggered by stressors (mental arithmetic, mirror tracing, interpersonal stress) in early to mid-adolescence, females responded with a greater increase in heart rate compared to their male adolescent counterparts (Ewart and Kolodner, 1991; Matthews et al., 1990; Syme et al., 2009). Anxiety responses to acute stressful stimuli are also correlated with the more advanced pubertal Tanner stages in females (Huerta and Brizuela-Gamino, 2002; Leen-Feldner et al., 2007). In addition to the response to acute stress, the response to chronic stress is exacerbated in adolescence: in individuals experiencing post-traumatic stress disorder, puberty onset was associated with an increase in negative affect, re-experience and hyperarousal (Carrion et al., 2002). Stress during adolescence can lead to different coping mechanisms, which may either be palliative or result in dysfunctional behavior, including risk-taking behavior and substance abuse.
Regional activity is increased in an age and gender-specific fashion throughout the cortico-limbic circuit which is involved in the stress response during adolescence, which includes the amygdala, hippocampus, insula and hypothalamus. Functional magnetic resonance imaging (fMRI) has been used to assess such activity during a psychosocial stress paradigm (Guyer et al., 2009). In females, increased activity of hypothalamus, insula, nucleus accumbens and hippocampus was observed during the stressor, an effect correlated with age. In contrast, for males, the stressor was associated with decreased insula activity, but no change in any of the other structures. Surprisingly, there was no change in amygdala activity during the task.
There are also gender differences in the size of these structures which are correlated with the onset of puberty, when amygdala volumes are greater in males, while hippocampal volumes are greater in females (Blanton et al., 2012; Lenroot and Giedd, 2010; Neufang et al., 2009), effects tightly correlated to pubertal status as well as to circulating levels of testosterone and estradiol, respectively. Intriguingly, sex differences in the growth of hippocampal neurons has been linked to circulating levels of E2 in males (Bowers et al., 2010). Taken together, these findings suggest that adolescence is a period when stress produces dynamic changes in both the structural and functional aspects of associated CNS circuits.
1.4 Puberty and pubertal hormones
Pubertal development in the human is a process which takes years to complete and can be defined by various end-points which culminate in reproductive function. The onset of reproductive function (gonadarche) can be identified in the human by functional markers which occur around 11–13 years of age, onset of spermatogenesis in the male (spermarche) and onset of menstruation in the female (menarche), which precede the first ovulation that usually occurs up to 1–3 years later (Apter and Hermanson, 2002; Tinggaard et al., 2012). Puberty can also be defined by outward physical signs, the secondary sexual characteristics, which are divided into Tanner Stages. Tanner stage 1 is pre-pubertal, and pubertal development proceeds across stages 2 – 5 (Apter and Hermanson, 2002; Tinggaard et al., 2012). In rodents, puberty onset is defined by pre-putial separation in the male and vaginal opening in the female, which occur on average at ~5 weeks of age (Tena-Sempere, 2010). In both humans and rodents, these external markers of puberty are preceded during the peri-pubertal period by increases in circulating levels of testosterone (T) or 17β-estradiol (E2), respectively (Apter and Hermanson, 2002; Janfaza et al., 2006; Safranski et al., 1993; Tena-Sempere, 2010; Tinggaard et al., 2012). In the female mouse, circulating levels of E2 increase more than two-fold five days before vaginal opening (Ahima et al., 1997). However, ovarian hormonal fluctuations are not evidenced until 10 d – 2 weeks after puberty onset (Hodes and Shors, 2005). In both humans and rodents, the age of puberty onset is variable, and influenced by a variety of factors including seasonal changes, body fat, nutrition, pheromones, etc. (Apter, 2003).
The onset of puberty in the human is preceded by maturation of the adrenal gland (adrenarche), which releases adrenal steroids, including the adrenal androgens dehydroepiandrosterone (DHEA) and DHEA-sulfate (DHEAS), as well as progesterone (P), without releasing cortisol (Miller, 2009). Although adrenarche may not occur in the rodent, several studies have documented rising levels of P which precede puberty onset (Khan et al., 2008; Mannan and O'Shaughnessy, 1988). Thus, the circulating levels of multiple steroids change as puberty approaches. Many of these steroids have documented effects on mood and cognition in adult rodents (E2, T, P, DHEA, DHEAS) (Frye et al., 2007; Frye and Walf, 2009a; Frye and Walf, 2009b; Maninger et al., 2009; Reddy et al., 1998; Walf and Frye, 2008), suggesting that deciphering underlying mood changes at puberty may be a complex process. This review will focus on one steroid, THP ([allo]pregnanolone or 3α-OH-5[α]β-pregnan-20-one), a metabolite of P and a potent modulator of the GABAA receptor (GABAR), which has been shown to play a pivotal role in anxiety (Shen et al., 2007). For the purposes of this review, the onset of puberty in the female mouse will be defined by vaginal opening, and the prior 5 peri-pubertal days will be defined as “pre-pubertal”.
1.4.1 THP
THP is formed from P via two enzymatic conversions, with 5α-reductase and 3α-hydroxysteroid oxidoreductase (3α-HSD) (Compagnone and Mellon, 2000; Karavolas and Hodges, 1990; Mellon and Vaudry, 2001). This steroid can be produced in the adrenal gland, with release into the general circulation before puberty onset. It can also be converted from P produced by the granulosa cells of the ovary before puberty. In non-pregnant adult mice, P is also primarily produced by the granulosa cells on the afternoon of proestrus with lower levels produced by the corpus luteum on the diestrous1 stage of the estrous cycle (Bentley, 1998). (In contrast to humans, for whom the corpus luteum is the primary producer of this steroid.) In addition, recent studies suggest that THP can be produced de novo directly in the brain from cholesterol via side chain cleavage enzyme; thus, it is classified as a neurosteroid (Compagnone and Mellon, 2000). This can occur in a variety of CNS sites, in both glia and neurons, including the CA1 hippocampal pyramidal cell (Agis-Balboa et al., 2006). It is not clear, however, if these elevated pre-pubertal THP levels produce any effect on behavior or mood. Recent studies suggest that circulating and/or CNS levels of this steroid increase in the pre-pubertal period, but decline to low levels at the onset of puberty, when levels fluctuate on a circadian cycle (Fadalti et al., 1999; McCartney and et al., 2007). Unlike most steroids, THP has no known effect at classic nuclear steroid receptors, but instead is a modulator of the GABAR (Smith et al., 2007).
1.4.2 THP functions as a stress steroid
In addition to its fluctuations at puberty and across the estrous cycle, THP also functions as a stress steroid. In rodents, its levels in brain increase by up to 20-fold after 45 minutes of restraint stress or other forms of stress (CO2 inhalation) (Higashi et al., 2005; Mukai et al., 2008; Purdy et al., 1991) when decreases in anxiety are observed (Barbaccia et al., 2001). Circulating levels of this steroid are also evidenced in humans after sustained stress associated with performance (Droogleever Fortuyn et al., 2004; Girdler et al., 2006). One potential mechanism for this increase in THP is via CRH and ACTH which have been shown to increase circulating and brain levels of the steroid (Torres et al., 2001), most likely due to activation of adrenal steroidogenesis by ACTH.
2.1 GABAA receptors
GABAA receptors (GABARs) mediate most inhibition in the brain (Olsen and Sieghart, 2009). They are pentameric membrane proteins commonly of the form 2α, 2β, and 1 γ (Chang et al., 1990), but many subunit combinations exist from a pool of 6α, 3β, 3γ, δ, ε, θ, π and ρ (Olsen and Sieghart, 2009). These receptors mediate a Cl− conductance, which is inhibitory in most CNS sites, including hippocampus, after early development (Rivera et al., 1999) due to increases in the K+-Cl− co-transporter KCC2 which maintains a hyperpolarizing Cl− gradient. GABA is depolarizing in the neonatal rodent CA1 hippocampus (Rivera et al., 1999; Waddell et al., 2011), however, which extends for a longer period in the male due to expression of the N+-K+-Cl−co-transporter NKCC1 (Nunez and McCarthy, 2007), which maintains a depolarizing Cl− gradient. Although other cellular events may underlie this phenomenon (Dzhala et al., 2012), the evidence is as yet inconclusive.
2.1.1 GABAR modulators
The GABAR is recognized as playing a pivotal role in anxiety (Rudolph et al., 1999) and is the target for most anxiety-reducing, sedative drugs (Olsen and Sieghart, 2009), including benzodiazepine, barbiturate and anesthetic drugs as well as for endogenous steroids such as THP (Fig. 1). Early studies established the effect of both isomers, 3α-OH-5α-THP and 3α-OH-5β-THP, which were shown to enhance GABA-gated current in vitro (Callachan et al., 1987; Gee et al., 1987; Harrison et al., 1987; Majewska et al., 1986) and GABA responses in vivo (Smith et al., 1987a), an effect also seen with the parent compound P (Smith et al., 1987b). More recent studies have identified the selective binding site for the steroid for potentiation of current generated by α1β2γ2 (Hosie et al., 2006), which is located in a cavity formed by the α subunit transmembrane domains. A separate binding site was identified for the steroid to directly activate the receptor (Hosie et al., 2006). Behaviorally, THP is anxiolytic (Bitran et al., 1999), anti-convulsant (Frye, 1995) and sedative (Lancel et al., 1997) in adults, as would be predicted for a positive GABA modulator.
Figure 1. The GABAA receptor.
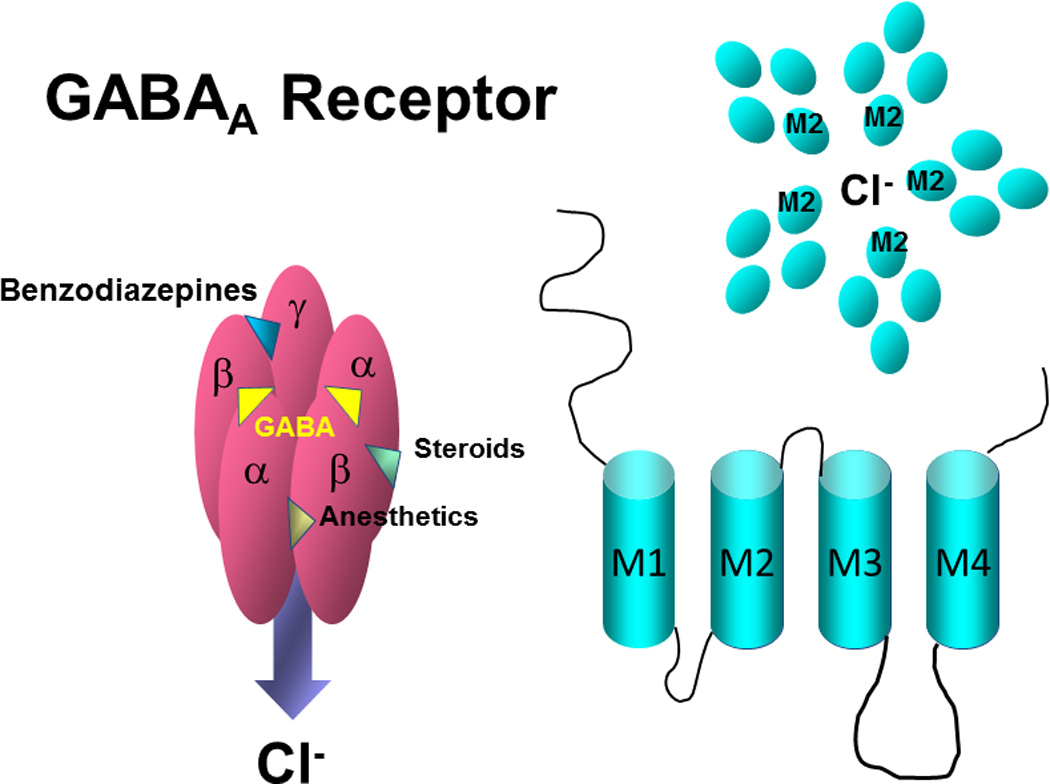
The GABAA receptor (GABAR) is a pentameric membrane protein typically composed of 2α, 2β and 1γ or δ subunit, but other possibilities exist from a pool of 6α, 3β, 3γ, δ, ε, θ, π and ρ. Each subunit is composed of 4 transmembrane spanning helices (M1–M4); M2 lines a central Cl− channel. The intracellular loop between M3 and M4 is 2-fold longer in α4 compared to α1, with very low homology. Binding of 2 GABA molecules (yellow) between α and β gates a Cl− current which can be increased by a variety of compounds including sedative drugs (benzodiazepines, anesthetics, alcohol) and the endogenous steroid THP. Benzodiazepines bind between α (α1–3 or 5) and γ, but do not bind to α4-containing GABARs.
2.1.2 GABAR sub-types
The most common GABAR subtype expressed in the brain is α1β2γ2, which expresses subsynaptically (Olsen and Sieghart, 2009), and encounters high concentrations of GABA (~1 mM) released as brief, episodic pulses (<1 ms) from local interneuron terminals (Maconochie et al., 1994). In addition to synaptic expression, however, select GABAR sub-types can also express at non-synaptic sites (extrasynaptically) on the neuron (soma and dendrites) where they underlie a tonic inhibitory current, that is estimated to contribute more to inhibitory tone than the phasic current from synaptic receptors (Bai et al., 2000). These include the α5β3γ2 GABAR, which predominates in the CA1 hippocampal pyramidal cell (Caraiscos et al., 2004; Wisden et al., 1992), and the α4βδ GABAR, which has highest levels of expression on dentate gyrus granule cells, thalamic relay nuclei and cortical pyramidal cells (Belelli et al., 2005; Chandra et al., 2006; Pirker et al., 2000; Stell et al., 2003; Stell and Mody, 2002; Wisden et al., 1992). The α4βδ GABAR is unique in that has a high sensitivity to GABA (EC50 = ~0.5 µM) (Brown et al., 2002; Sundstrom-Poromaa et al., 2002) and relatively little desensitization (Bianchi et al., 2002) under steady-state conditions, making it well-suited for its extrasynaptic location, where it would come into contact with ambient GABA (~1 µM), maintained by the GABA transporters (Wu et al., 2003). The receptor can desensitize in response to rapid exposure to agonist, however, suggesting that it may not be activated by transmitter spillover (Bright et al., 2011), while receptors distant from the synapse would maintain tonic inhibition supplied by the ambient GABA. This receptor is also unique in that its response to GABA modulatory drugs is distinct from most other GABAR subtypes; it is insensitive to benzodiazepines (Brown et al., 2002; Wieland et al., 1992) and highly sensitive to steroids such as THP and THDOC (Belelli et al., 2002; Bianchi and Macdonald, 2003; Brown et al., 2002; Wohlfarth et al., 2002), which are positive modulators of the receptor. Although conflicting reports exist, they are also sensitive to low concentrations of alcohol in the range which is relevant for social drinking (Sundstrom-Poromaa et al., 2002; Wallner M et al., 2003). In addition, GABA acts as a partial agonist at these receptors, where most modulators, such as THP and THDOC, act by increasing receptor efficacy (Bianchi and Macdonald, 2003; Zheleznova et al., 2008). In single channel studies, the steroid THDOC increased receptor efficacy by adding a third open state of longer duration to the two open states recorded from the receptor in the absence of steroid (Wohlfarth et al., 2002). Neurosteroids can increase the tonic current recorded from dentate gyrus granule cells (Stell et al., 2003), although some studies have not observed this effect, due to degradation of steroids such as THP and THDOC in this CNS region (Belelli and Herd, 2003). Recent studies suggest that α4 and δ co-express with β2 based on the lack of tonic current in dentate gyrus granule cells in β2 knock-out mice (Herd et al., 2008).
2.1.3 Chloride-dependent effects of THP at α4ββ GABARs
THP effects at α4βδ GABARs are unique in that they are dependent upon the direction of the Cl− current generated by the receptor. Most electrophysiology studies in recombinant receptors record current which is in the depolarizing direction (inward current) due to the ion gradients used. Under these conditions, THP increases current gated by α4βδ GABARs (Belelli et al., 2002; Bianchi and Macdonald, 2003; Wohlfarth et al., 2002). This effect would be relevant for its effect to increase tonic current in the dentate gyrus, which is depolarizing, but a shunting inhibition (Staley and Mody, 1992). However, THP decreases hyperpolarizing current generated by these receptors (Shen et al., 2007). These effects have been reported in recombinant receptors where either the ion gradient or holding potential were varied, and occurred in the absence of changes in the reversal potential, suggesting that other conductances are not involved (Shen et al., 2007). The effect of THP on hyperpolarizing current generated by α4βδ GABARs is due to very rapid (<10 ms) acceleration of desensitization (Shen et al., 2007), and is consistent with findings from Macdonald and colleagues who have shown that these receptors desensitize more rapidly when the current is in the hyperpolarizing direction (Bianchi et al., 2002; Haas and Macdonald, 1999), an effect which is accelerated by the steroid (Bianchi and Macdonald, 2003). This is in contrast to the α5β3γ2 GABAR, where desensitization is accelerated when the current is depolarizing (Burgard et al., 1996). This effect of the steroid is dependent upon the positively charged residue arginine 353 in the TM3-TM4 loop (Shen et al., 2007), which may serve as a modulatory site for Cl−. Modulatory effects of Cl− have been reported for other GABAR subtypes (Houston et al., 2009; Olsen and Snowman, 1982).
This chloride-dependent effect of the steroid has implications for its ultimate behavioral effect, where the typical effect of the steroid is to increase inhibition and reduce anxiety (Bitran et al., 1999). However, increased expression of α4βδ GABARs which generate a hyperpolarizing current would decrease inhibition and paradoxically increase anxiety, as has been shown at puberty (Shen et al., 2007).
2.2 Plasticity of the α4ββ GABARs at puberty
α4βδ GABARs display a high degree of plasticity even in CNS areas where their expression is normally low, including CA1 hippocampus and midbrain central grey (Pirker et al., 2000; Wisden et al., 1992). Their expression is increased by exposure to the steroid THP (Kuver et al., 2012; Maguire and Mody, 2007; Shen et al., 2005), for periods of 30 min to 48 h, as well as by fluctuating levels of THP produced by exogenous steroid administration or by endogenous cyclicity across the estrous cycle and pregnancy (Griffiths and Lovick, 2005a; Lovick et al., 2005b; Maguire and Mody, 2009; Maguire et al., 2005; Smith et al., 2006).
Our recent studies show that α4βδ GABAR expression also increases markedly at the onset of puberty in the female mouse (PND ~35–44, assessed by vaginal opening) from almost undetectable levels before puberty (Shen et al., 2007; Shen et al., 2010a). These receptors localize exclusively at non-synaptic (extrasynaptic) sites on the dendritic shaft as well as on the spine, adjacent to the excitatory synapse. Although in the cortex, GABAR expression is seen on the spine neck of pyramidal cells where it receives direct GABAergic input (i.e., is synaptic) (Kubota et al., 2007; Nusser et al., 1996), this is the first demonstration of a GABAR localized to the spine on a CA1 hippocampal pyramidal cell. Previous reports have shown that GABARs typically express on the soma (80%) and dendritic shaft (20%) of these CA1 neurons (Megias et al., 2001). Expression increased by more than 3 to 8-fold on the dendritic shaft as well as the dendritic spines of CA1 hippocampal pyramidal cells at puberty, which was quantified in the proximal stratum radiatum. The functional expression of this receptor was verified by responses of the pyramidal cells to the GABA agonist gaboxadol at a concentration (100 nM) selective for α4βδ GABARs (Brown et al., 2002; Meera et al., 2011). Negligible levels of α4 expression are seen on the interneurons in CA1 hippocampus at the onset of puberty, however (unpublished data), which is reflected by the fact that steroid effects on the pyramidal cell are identical whether or not interneuron activity is abolished with the voltage-gated Na+ channel blocker tetrodotoxin (TTX) (Shen et al., 2007).
2.2.1 The direction of GABAergic current in CA1 hippocampal pyramidal cell dendrites
The current generated by dendritic α4βδ GABAR populations on CA1 hippocampal pyramidal cells is hyperpolarizing at puberty. This was determined with direct application of a low concentration of the GABA agonist gaboxadol to the dendrites of a CA1 hippocampal pyramidal cell in recordings using cell-attached mode or gramicidin perforated patch recordings (Shen et al., 2007), which would not disrupt the chloride gradient. Early studies (Alger and Nicoll, 1982) suggesting that the dendritic GABAergic current is depolarizing on CA1 hippocampal pyramidal cells used high concentrations of GABA which would preferentially produce depolarizing responses in low volume systems like the dendrites, but not at the soma, where the larger volume would permit greater adaptive responses. This is likely due to the fact that excessive activation of dendritic GABARs collapses the chloride gradient and permits passage of other ions, such as HCO3−, which would elicit depolarizing responses (Staley and Proctor, 1999), as recently demonstrated (Isomura et al., 2003). In contrast, in the same study (Alger and Nicoll, 1982), dendritic application of the GABA agonist gaboxadol, which would produce less activation of GABARs, generates hyperpolarizing responses on CA1 hippocampal pyramidal cells, as do lower concentrations of GABA itself (Lambert et al., 1991).
2.2.2 THP effects on CA1 hippocampal pyramidal cells at puberty
Because THP exerts chloride-dependent effects at α4βδ GABARs, the effect of this steroid at puberty would be dependent both upon the increased expression of α4βδ GABARs on CA1 hippocampal pyramidal cells as well as on the direction of GABAergic current, which is hyperpolarizing at puberty (Shen et al., 2007). Thus, under these conditions, THP acts to reduce the tonic inhibitory current at puberty (Fig. 3) which was a post-synaptic effect because it is observed when action potentials were blocked with TTX in the presence of applied GABA (1 µM) (Shen et al., 2007). THP also increases neuronal excitability at puberty (Fig. 3), assessed both with cell-attached recordings of spontaneous spiking as well as by current clamp recordings (Shen et al., 2007). In the latter, THP reduced the threshold for spiking by increasing the input resistance of the neuron. These paradoxical excitatory effects of the steroid at puberty were not seen in the δ −/− mouse, implicating the increased expression of α4βδ GABARs at puberty as the mechanism. In contrast, the steroid reduced neuronal excitability before puberty, as expected for its role as a positive GABA modulator at other receptor subtypes where it is known to increase inhibition.
Figure 3. THP effects on CA1 hippocampal pyramidal cells at puberty.
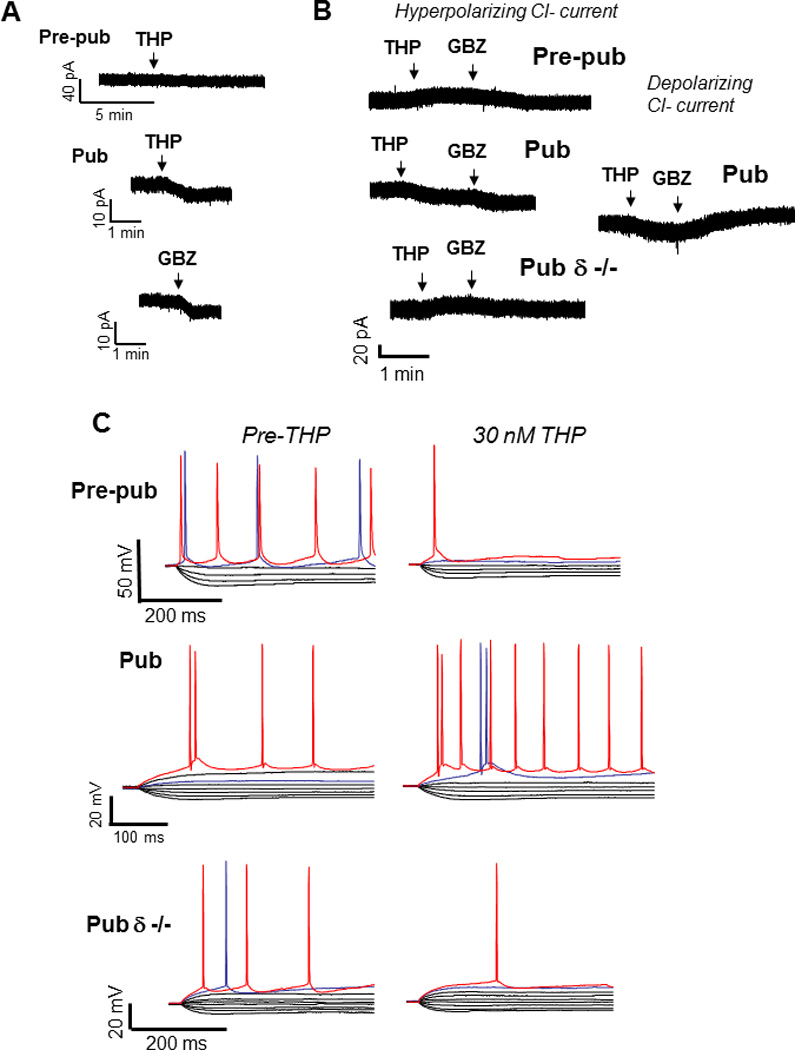
THP reduces the tonic GABAergic current and increases excitability of CA1 hippocampal pyramidal cells at puberty. A. THP effects on the tonic current recorded from CA1 hippocampal pyramidal cells in the slice using gramicidin perforated patch techniques to maintain the internal Cl− milieu. 1 µM TTX, 1 µM GABA and 2 mM kynurenic acid were added to isolate the GABAergic post-synaptic component. L-655, 708, CGP55845 and TEA were also added to block α5 and GABAB receptors and K+ channels, respectively. Pre-pub, pre-pubertal; Pub, pubertal; GBZ, gabazine, a GABA antagonist (presented for comparison). THP reduces the current in Pub slices. (Representative of 5 cells/group) B. Hyperpolarizing current recorded from CA1 hippocampal pyramidal cells in the slice using whole cell patch clamp techniques (ECl = –70 mV, –50 mV holding potential; pipette solution, K-gluconate; bath, 200 nM gabazine to block synaptic current and 2 mM kynurenic acid to block excitatory current). Inset, effects of THP on the depolarizing tonic current at puberty (ECl = −30 mV, pipet solution, CsCl). THP reduces hyperpolarizing current in wild-type but not δ knock-out mice, but potentiates depolarizing current. (Representative of 8 – 12 cells/group) C. Whole-cell current-clamp recordings reveal voltage responses recorded in response to increasing 0.3-nA current injection (initial current −1 nA). (The THP trace lacks the 800-pA current trace for ease of comparison.) THP lowers the current threshold for spiking of pyramidal cells at the onset of puberty in wild-type but not δ knock-out mice. Red trace, equivalent current injection, threshold for the less excitable state. Blue trace, equivalent current injection, threshold for the more excitable state. (Representative of 7 – 8 cells/group) (Revised and used with permission (Shen et al., 2007).)
2.2.3 The ultimate effect of THP on CA1 pyramidal cell function at puberty
THP effects at α4βδ GABARs are distinct because lower, physiological concentrations of this steroid (30 nM) would have a greater effect at α4βδ GABARs compared to other receptor sub-types, where significantly higher concentrations (>100 nM) are required (Belelli et al., 2009). Therefore, at puberty, THP effects (30 nM) on CA1 hippocampal pyramidal neurons would primarily impact the α4βδ GABAR population and the tonic current it generates (Fig. 4A). In addition, recent studies have shown that the GABAergic tonic current contributes a greater net effect on neuronal excitability than does the phasic current (Bai et al., 2000) because it produces a greater total charge transfer than the synaptic current (Fig. 4A). Although GABA is a partial agonist at α4βδ GABARs (Bianchi and Macdonald, 2003), it has greater potency at these receptors, and the 1 µM concentration of ambient GABA would preferentially activate these receptors (Brown et al., 2002). THP increases the input resistance (Rm) by accelerating the desensitization of α4βδ GABARs localized to the dendritic shaft, thus reducing this source of shunting inhibition. (Similar effects have been reported due to the knock-out of α4βδ GABARs (Chandra et al., 2006). This effect occurs within 10 ms (Fig. 2) because fast desensitization is rapid (Jones and Westbrook, 1995), and this is what renders the effect of THP inhibitory. In contrast, synaptic GABARs, such as α1β2γ2, are potentiated only to a modest degree by 30 nM THP (~10%)(Stell and Mody, 2003), but this has less of an impact on neuronal function because the total charge transfer is less than the corresponding change in the α4βδ GABAR population (Fig. 4A). In addition, although tonic current is also generated by α5β3γ2 GABARs (Caraiscos et al., 2004), these are relatively insensitive to THP (Shen et al., 2007) and thus would not be a factor in THP’s effects at puberty.
Figure 4. Relative changes in the total charge transfer and neuronal function produced by THP in pubertal hippocampus.
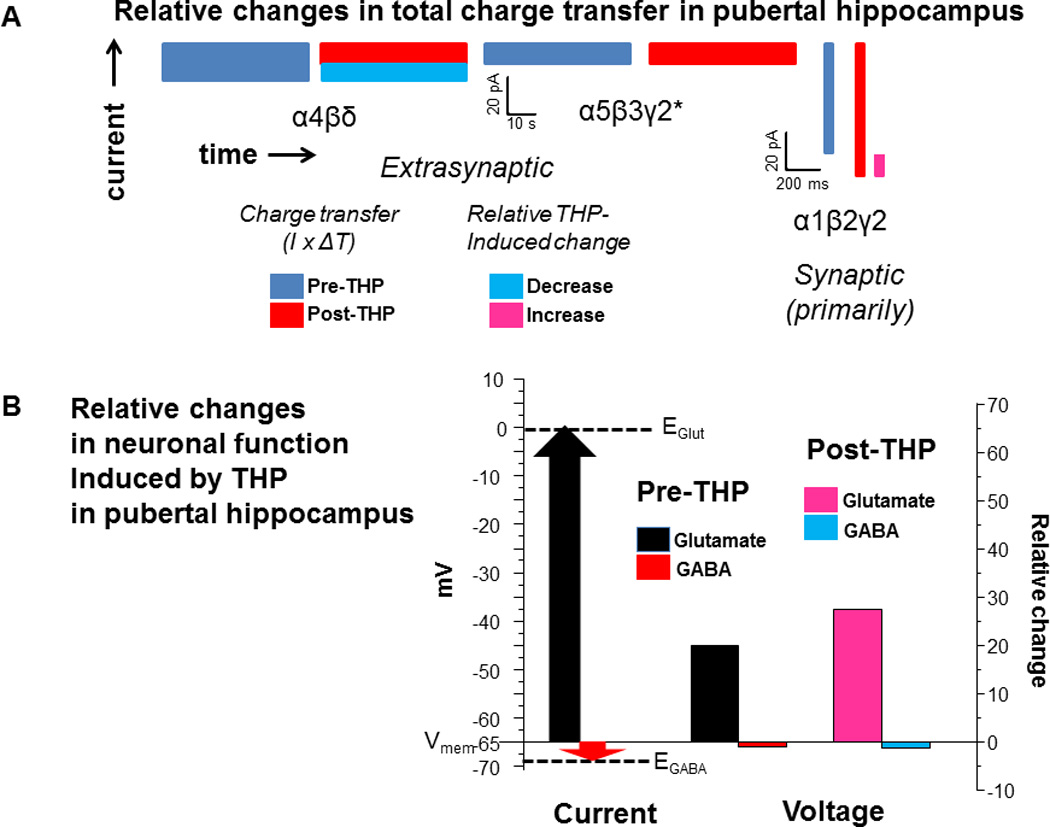
A, total charge transfer. Estimates of charge transfer based on average values obtained from whole cell patch clamp recordings of CA1 hippocampal pyramidal cells in the slice, before and after 30 nM THP across a 60 s time period (boxes indicate approximate total charge transfer: current (y axis) by total time (x axis). α4βδ, 20 pA tonic current × 60,000 ms; α5β3γ2, 10 pA tonic current × 60,000 ms; α1β2γ2, 60 pA, phasic current × 3600 ms (5 ms half-width × 12 Hz frequency). THP decreases the tonic current by 45% at puberty when α4βδ expression is high, has no effect on α5β3γ2 (*based on results from pre-pubertal mice, where α5β3γ2 represent the majority of the tonic current) and increases the half-width of the phasic current by ~10% (likely to represent α1β2γ2 GABARs). Therefore, the relative decrease in total charge transfer produced by THP would be approximately 20-fold greater for the tonic current compared to the corresponding increase in total charge transfer produced by THP on the phasic current. These values are similar to what has been reported for anesthetic effects (Bai et al., 2000). B, neuronal function. Y axis (left), Voltage (mV) indicated for the membrane potential (Vmem), reversal potential for glutamate (EGlut, dashed line) and reversal potential for GABA (EGABA, dashed line) for a typical mature CA1 hippocampal pyramidal cell. Because Vmem values are considerably further from EGlut compared to EGABA, opening of the channels on these receptors produces greater excitatory current relative to inhibitory GABAergic current (arrows). Y axis (right), relative changes in current and the resulting change in voltage. After 30 nM THP, there is a ~37% increase in input resistance (Shen et al., 2007). This produces a greater increase in the depolarization produced by glutamate (Glut) relative to the negligible decrease in hyperpolarization produced by GABA, calculated using Ohm’s Law. Thus, THP has an excitatory effect at puberty.
Figure 2. Chloride-dependent effects of THP at α4βδ GABAA receptors.
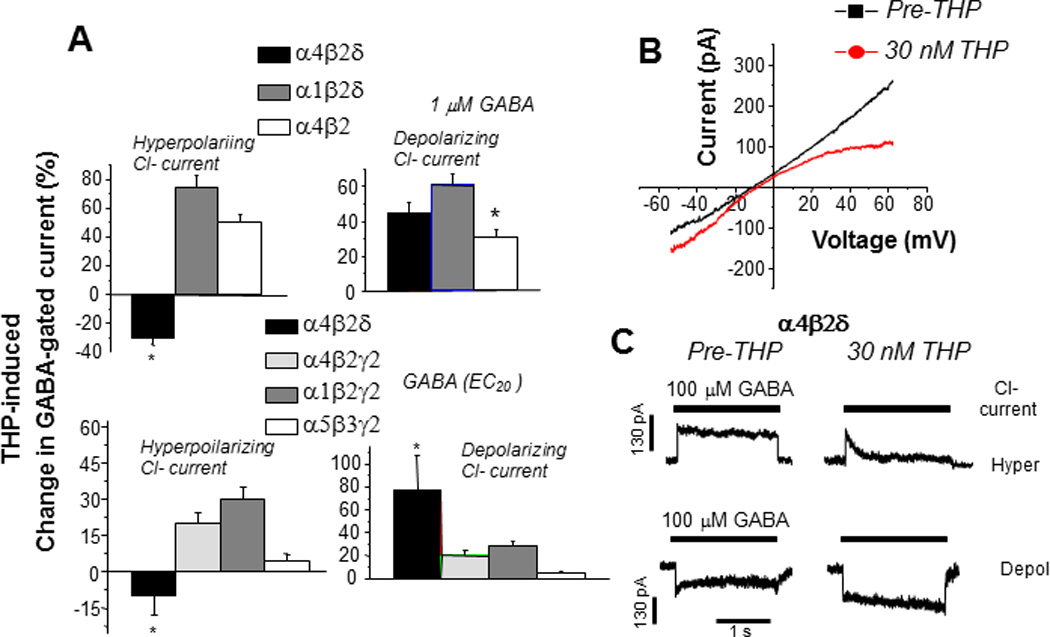
The neurosteroid THP decreases hyperpolarizing current gated by α4βδ GABAA receptors transiently expressed in HEK-293 cells and recorded with whole cell voltage clamp techniques. A. Mean effects of THP on hyperpolarizing and depolarizing currents in response to 1 µM GABA (upper panel) or the GABA EC20 (lower panel; α4β2δ, 0.1 µM; α4β2γ2, 5 µM; α1β2γ2, 10 µM; α5β3γ2, 5 µM) from 6–7 cells for each group (*P = 0.05 versus the other receptor subtypes). n= 6 – 8 cells/group. B. Effects of 30 nM THP on current generated by a voltage ramp over 400 ms where the ECl = 0 mV. THP increases depolarizing current (voltage < 0 mV) but decreases hyperpolarizing current (voltage > 0 mV). (Leak-subtracted current is presented as the average of three traces). n=5 – 6 cells/group. C. Effects of THP on desensitization of hypepolarizing (upper traces) and depolarizing (lower traces) current at α4βδ receptors (representative of 6 cells per group). THP accelerates desensitization of the hyperpolarizing current. (Used with permission (Shen et al., 2007).)
The decrease in the input resistance of CA1 pyramidal cells would permit greater voltage changes produced by both excitatory and inhibitory currents. However, the GABAergic currents in CA1 hippocampus produce a shunting inhibition because the reversal potential for chloride is close to the membrane potential (Xu and Sastry, 2007) and produces less of a driving force on inhibitory currents than exists for excitatory currents (Fig 4B). Therefore, a change in input resistance would have a greater impact on the ability of excitatory currents to generate an action potential (Fig. 4B). Thus, an increase in input resistance would lead to increased neuronal excitability.
2.2.4 Stress steroid-induced anxiety at puberty
The GABAR plays a pivotal role in the generation and modulation of anxiety behavior (Rudolph et al., 1999). Although several brain areas are linked with anxiety behavior, the hippocampus, part of the limbic circuitry which regulates emotion, is one which is directly implicated in THP’s effects on anxiety (Bannerman et al., 2004; Bitran et al., 1999). Therefore, one possible mediator of anxiety associated with puberty onset may be linked to the expression of this receptor in CA1 hippocampus. In order to test this possibility, female mice were administered THP (10 mg/kg, i.p.), acutely at puberty onset or before puberty, and tested for anxiety using the elevated plus maze, an animal model of anxiety. THP exerted effects which were opposite in the two groups: it paradoxically increased anxiety behavior at puberty (Fig. 5), but decreased anxiety in the pre-pubertal animals (Shen et al., 2007), similar to what has been reported in adults (Bitran et al., 1999). The anxiety-producing effects of THP were not seen in the δ knock-out mouse, implicating α4βδ GABARs. The effect of exogenous THP administration was replicated by subjecting the mice to 45 min of restraint stress, which is known to increase release of THP (Higashi et al., 2005). 20 min after the restraint stress, anxiety behavior was reduced in the pre-pubertal mice, but increased in the pubertal mice (Shen et al., 2007). This anxiety-producing effect of the steroid was not seen in the δ −/− mouse and prevented both by finasteride, which prevents formation of THP (Compagnone and Mellon, 2000), as well as by prior administration of 3β-OH-THP, an inactive steroid (Shen et al., 2007), which can prevent effects of THP on GABA-gated current. Taken together, these findings suggest that stress-induced release of THP produces anxiety at puberty in contrast to its well-established effect to decrease anxiety at other ages (Fig. 6, Summary).
Figure 5. THP paradoxically increases anxiety after the onset of puberty.

Alterations in anxiety produced by restraint stress or injection of THP (10 mg/kg, i.p.) are presented as a percentage change in open arm time in the elevated plus maze compared to mean values from a sham control group, age-and genotype-matched to the experimental group. To test the role of THP release in the stress response, in some cases the inactive 3β-OH isomer of THP (stress + 3β-OH-THP) or finasteride (a 5α-reductase inhibitor) were preadministered. Replacement THP (10 mg/kg, i.p., in oil, for 3 d) was also administered to prevent the decline in THP at puberty. n = 6 – 9 mice for each group. *P = 0.05 versus control, **P = 0.05 versus Pre-pub. (Used with permission (Shen et al., 2007).)
Figure 6. Summary of the effects of the stress steroid THP on CA1 hippocampal pyramidal cells at puberty.
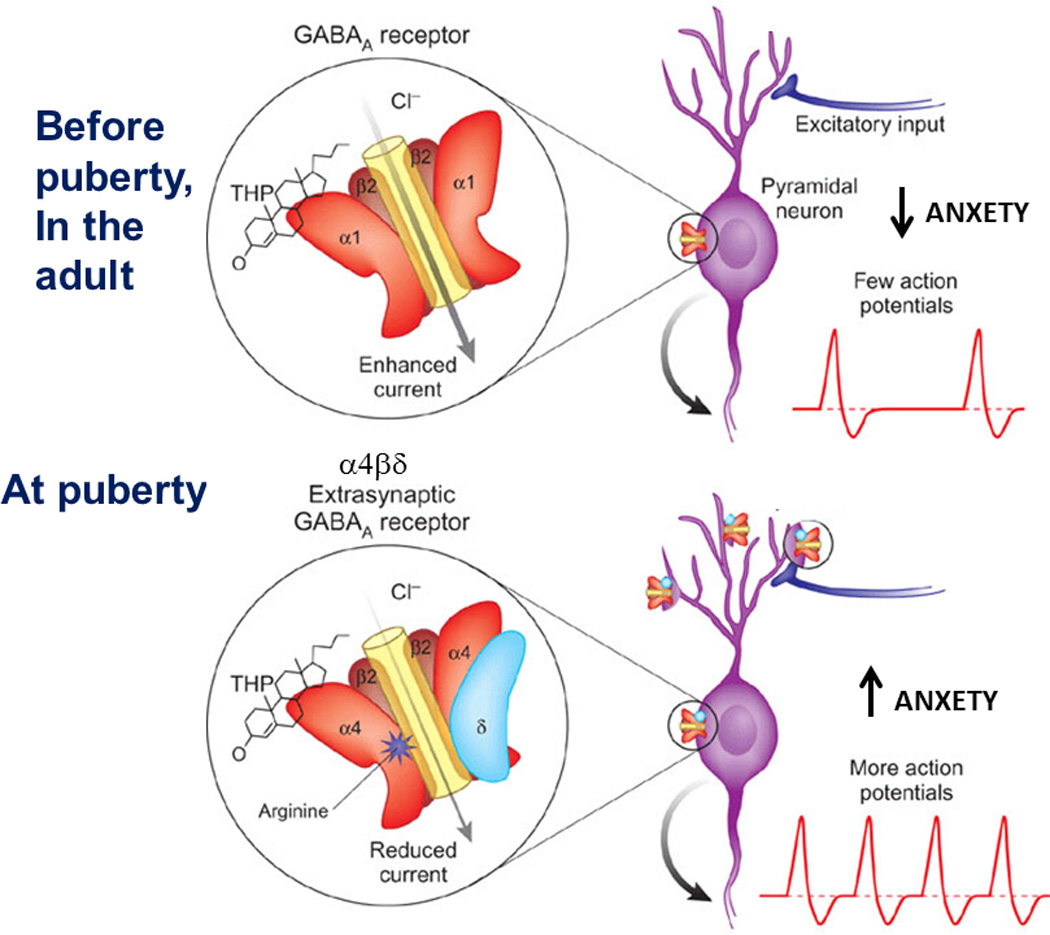
Before puberty and in the adult (upper panel) α4βδ GABAR expression is low. THP potentiates non- α4βδ GABARs, which would reduce the likelihood of action potentials. During the pubertal period (lower panel), however, α4βδ GABAR expression increases on the dendrites and spines of the pyramidal cells (lower panels). Current from these receptors is reduced by THP, which would increase neuronal excitability, increasing the likelihood of action potential generation. This effect of the stress steroid is linked both to increased anxiety, which may be a result of its effects at the α4βδ GABAR. (Revised and used with permission (McCarthy, 2007).)
Alternative mechanisms for THP-induced anxiety have been recently presented and include its actions on paraventricular (PVN) hypothalamic neurons which exhibit depolarizing GABA responses during stress when THP can release corticotrophin-releasing hormone (CRH) (Sarkar et al., 2011). This is in contrast to the typical increase in inhibition of these neurons mediated by THP (Belelli et al., 2009), which decreases release of CRH (Patchev et al., 1994). Although depolarizing responses of PVN neurons have not been shown to occur at puberty, these represent another potential mechanism for THP-generated anxiety. However, studies comparing effects of restraint stress in CRH knock-out mice have noted normal behavioral responses despite the lack of CRH release (Dunn and Swiergiel, 1999), suggesting that alternative stress mechanisms exist.
2.3 Paradoxical effects of THP and P on anxiety and dysphoric mood
The paradoxical anxiety-producing effect of THP observed at puberty onset in female mice which is linked with α4βδ GABAR expression (Shen et al., 2007) has also been observed with P, the parent compound for THP, in human studies at hormonal transitional states. In women with PMDD, negative mood is correlated with administration of E2 and P after cessation of normal ovarian cyclicity with an analog of luteinizing hormone (Schmidt et al., 1998). This is in contrast to normal women who experience positive mood changes in response to administration of these ovarian hormones. In another study, circulating levels of THP were positively correlated with negative mood systems in women with PMDD (Freeman et al., 2002), again in contrast to asymptomatic controls. In post-menopausal women, ovarian steroids are also correlated with negative mood symptoms when the circulating levels of P and THP are within a normal luteal range (Andreen et al., 2004), suggesting that these paradoxical effects of the steroid may be dose-dependent. Although one study which examined direct effects of THP on the acoustic startle response did not find effects in women with PMDD (Kask et al., 2009), this study did not find anxiety-reducing effects of the steroid in normal women, suggesting that other factors may be required. For instance, in rodent studies the effect of THP on anxiety is most robust in behavioral assays linked with an aversive stimulus (Smith et al., 2006) which was not included in the human study. There are several possibilities for the paradoxical effects of THP in these studies, including potential pre-synaptic effects to disinhibit principal cells, as seen in the periaqueductal grey, when α4βδ GABARs increase expression on diestrus (Brack and Lovick, 2007; Griffiths and Lovick, 2005b). Although it is not possible to determine potential mechanisms in human studies, the fact that women with PMDD have a decreased response to benzodiazepines (Sundstrom et al., 1997) is consistent with increased expression of α4βδ GABARs post-synaptically. Benzodiazepine modulation of the GABAR requires a γ2 subunit (Sigel, 2002) and an α subunit of the form α1–3,5. GABARs containing the α4 subunit lack a benzodiazepine binding site because of the histidine to arginine substitution at residue 99 in α4 (Wieland et al., 1992).
2.4 Puberty as a THP “withdrawal” state
Recent studies in both the human and mouse report that both P and its neuroactive metabolite THP are also increased during the pre-pubertal period from low levels seen in early development (Fadalti et al., 1999; Mannan and O'Shaughnessy, 1988; McCartney and et al., 2007). However, THP levels drop by 60–70% around the onset of puberty (Fadalti et al., 1999; Mannan and O'Shaughnessy, 1988; Shen et al., 2007) unlike most other steroids including E2, 17α-hydroxyprogesterone, 20α-dihydroprogesterone and androgens (androstenedione, T) which continue to increase during the pubertal period into adulthood (Mannan and O'Shaughnessy, 1988). Both ovarian production and hippocampal concentrations of THP were shown to drop precipitously at the onset of puberty in the mouse (Mannan and O'Shaughnessy, 1988; Shen et al., 2007). The decrease in ovarian release of THP around puberty onset was seen after exposure to the precursor steroid pregnenolone, when release of other steroids increased, suggesting that it is due to the relative reduction in activity of synthetic enzymes 5α-reductase and 3α-HSD, rather than due to a reduction in substrate (Mannan and O'Shaughnessy, 1988). Interestingly, the 3β-OH-isomer of THP (3β-OH-THP), which is inactive as a GABA modulator, is released at high levels by the ovary around PND 21 but is reduced to much lower levels of production before PND 29. Because this inactive steroid can block the effect of THP at GABARs (Shen et al., 2007), these findings suggest that the pre-pubertal period (~PND 29–35) may be a time when inhibitory processes in the brain may be enhanced.
This decline in THP (“THP withdrawal”) was shown to be responsible for the increased expression of α4βδ GABARs at puberty. Replacement THP (10 mg/kg, i.p. × 3) during pubertal onset prevented the increase in α4βδ expression (Shen et al., 2007), as well as THP’s effect to reduce the GABAergic tonic current. This also prevented the paradoxical excitatory effects of THP at this time on CA1 hippocampal pyramidal cell excitability and on anxiety behavior (Shen et al., 2007). THP withdrawal, induced by administration of the 5α-reductase blocker finasteride, has also been shown to increase α4βδ GABAR expression in hippocampus of pre-pubertal mice (Smith et al., 2006), when THP again decreases the tonic inhibitory current generated by these receptors.
Other studies (Griffiths and Lovick, 2005a; Lovick et al., 2005a) have shown that α4βδ expression in interneurons of the periaqueductal grey is increased after P withdrawal or on the stage of late diestrus of the estrous cycle, which follows a peak in circulating levels of P. In this case, increases in tonic inhibition of these interneurons would increase excitability of the output neurons, which are implicated in the circuitry of panic disorder (Lovick, 2000). In both this study and ours, THP withdrawal may represent a model of premenstrual dysphoric disorder (PMDD), which can occur during decreases in circulating levels of P (Cunningham et al., 2009; Rapkin and Winer, 2009), when stress-triggered anxiety and panic attacks are more likely to occur (Nillni et al., 2011; Vickers and McNally, 2004; Yonkers et al., 2008).
THP withdrawal may also be reflected by conditions during the post-partum period when THP levels decline precipitously, and alterations in α4βδ expression occur in dentate gyrus granule cells (Maguire et al., 2009; Maguire and Mody, 2009; Sanna et al., 2009). This change in receptor expression may represent a homeostatic response to maintain normal levels of neuronal excitability.
3.1 Cognition and adolescence: potential sex differences
It has been widely suggested that the onset of puberty may mark the end of a critical period for optimal learning of certain tasks, including motor learning and language acquisition. This has been reported for learning a second language (Johnson and Newport, 1989), including sign language (Newman et al., 2001), and appears to be a more severe boundary for those with pre-existing language deficits (Wright and Zecker, 2004). Although early development may be the most important period for language development, learning a language before puberty preferentially improves ability to detect semantic errors, a fundamental concept in learning (Weber-Fox and Neville, 2001). A number of studies have suggested similar critical periods for optimal cortical reorganization in the visual and auditory systems (Kral et al., 2001; Sharma et al., 2007; Voss et al., 2008) for those with visual and auditory deficits. Interestingly, both structural and functional changes are apparent in musicians who begin training before puberty compared to later learners and non-musicians (Bailey and Penhune, 2012; Imfeld et al., 2009; Schlaug et al., 1995), suggesting that the pubertal period may have important implications for brain plasticity.
In addition to studies implicating pubertal onset as demarcating the end of a period when CNS plasticity is optimal, other studies have suggested that early adolescence may be a time when learning of certain tasks is transiently impaired. Spatial learning with a computer video game is robust in pre-pubertal children (5th graders) (Subrahmanyam and Greenfield, 1994) and young adults (19–22 year olds) (Pepin and Dorval, 1986a), but not evident in young adolescents (12–13 year olds) (Pepin and Dorval, 1986b; Shavalier, 2004). One prospective study investigated latency to identify verbal and visual mismatches in facial expression across development (McGivern et al., 2002). Clear deficits were evident during early adolescence which recovered by late adolescence. In both of these spatial learning and mismatch detection studies, greater deficits can be seen in females, suggesting a potential gender difference in learning deficits at puberty. A more recent study (Gur et al., 2012) surveying a comprehensive battery of neurocognitive parameters reported global improvement in almost all tasks from age 8 – 21 in both sexes. However, spatial memory declined slightly in 12 – 15 year old females, consistent with pubertal onset, while certain measures of executive control (abstraction) and non-verbal reasoning declined in 12 – 13 year old males, suggesting a sex difference in adolescent performance. Interestingly, for two parameters (spatial memory and non-verbal reasoning), performance for both sexes was highest in 10 – 11 year old children and declined significantly in late adolescence and early adulthood, suggesting that these tasks are optimal before puberty onset. Unfortunately, for many of these studies the use of purely chronological age and the grouping of several age spans may at least partially mask pubertal effects on the measured end-points because the age of puberty onset is variable.
Sex differences in spatial learning have been widely reported, where performance in males is frequently better than for females. Several studies have suggested that these sex differences in spatial memory appear at or after the onset of puberty (Ardila et al., 2011; Gur et al., 2012; Kanit et al., 2000), although there are conflicting reports (Newhouse et al., 2007).
3.1.1 Spatial learning and the hippocampus
Although many CNS targets may contribute to these observed learning deficits in the pubertal period, the hippocampus is one site which plays a pivotal role in spatial learning (Bannerman et al., 2004; Burgess et al., 2002; Maguire et al., 1997) and memory consolidation (Pastalkova et al., 2006). In addition, the hippocampus has more recently been shown to be important for mismatch detection (Kumaran and Maguire, 2006) which is the CNS process which underlies error detection, a critical component of the learning process. Interestingly, hippocampal volume is also correlated with spatial learning during adolescence in humans (Herting and Nagel, 2012), and this parameter can be further correlated with aerobic fitness. These findings suggest the possibility that alterations in hippocampal function may play a role in learning deficits at puberty.
3.1.2 Effect of stress on learning in adolescence
Many studies have considered the effect of stress during adolescence on cognitive function. In general, high levels of chronic stress during adolescence impair cognition (Bellani et al., 2006; Sterlemann et al., 2010), an effect which may be due to the neurodegenerative effects of the stress hormone corticosterone (Watanabe et al., 1992). However, the effect of a stressor at this time would depend upon whether the stress is acute or chronic, the degree of stress and the cognitive state of the individual, as well as the perceived locus of control. In one study (Wolk and Bloom, 1978), middle school children were assessed in their performance on several academic tasks during stressors (strong environmental interruptions, disturbances and unpredictable obstacles) or no stress conditions. Those students with an internal locus of control exhibited improved performance during stress, while those with an external locus of control had diminished performance as stress level increased. The effect of stress on learning may also be different for simple versus complex learning tasks, as first described by the Yerkes-Dodson law (Yerkes and Dodson, 1908), which describes an inverted U relationship between stress and the learning of simple tasks (Lupien et al., 2007).
In animal studies, acute stress has also been shown to enhance learning during the pubertal period. In one study (Hodes and Shors, 2005), both male and female pubertal rats (35–40 days of age) exhibited improved performance on a trace eyeblink conditioning task 24 h following tailshock compared to unstressed animals. This effect was independent of the estrous cycle in females, and was not seen in pre-pubertal animals, despite robust increases in circulating corticosterone in response to the shock (Hodes and Shors, 2005). Facilitating effects of tailshock stress on learning was also not seen in adult females (Wood and Shors, 1998), suggesting that the pubertal period may represent a unique period for stress effects on cognition. Other studies have also reported facilitating effects of stress on learning in adolescence (Uysal et al., 2012), where a sex difference exists in terms of the shock intensity which produces optimal results. Although CRH and cortisol/corticosterone may be mediators of stress effects in many of these studies, especially those investigating effects of chronic stress, there are reported dissociations between CRH/cortisol release and stress effects (Dunn and Swiergiel, 1999; Hodes and Shors, 2005). Therefore other systems may also play a role in stress effects on learning during the pubertal period.
3.2 GABAA receptors and learning
Many studies have shown that drugs which act as positive modulators of the GABAR impair learning in adults. It is well known that benzodiazepine tranquilizers are amnestic, as are anesthetics such as propofol (Veselis et al., 2009). Both alcohol and the stress steroid THP impair spatial learning on the Morris Water Maze (Matthews et al., 2002). In the adult, GABARs containing the α5 subunit are the predominant extrasynaptic inhibitory receptor on the dendritic shaft (Brunig et al., 2002). Genetic knock-out or pharmacological knock-down of this receptor prevents learning impairment produced by the anesthetic etomidate (Cheng et al., 2006), and enhances a number of learned behaviors including fear conditioning (Chambers et al., 2003; Collinson et al., 2002; Crestani et al., 2002). These effects of positive GABA modulators are also evidenced in in vitro models of synaptic plasticity, including long-term potentiation (LTP). LTP can be reduced by a wide variety of these GABA modulators, including alcohol, THP and propofol (Izumi et al., 2005; Nagashima et al., 2005; Tokuda et al., 2011), suggesting a potent effect of inhibition in the hippocampus in limiting synaptic plasticity.
3.3 α4βδ GABARs and synaptic plasticity at puberty
The emergence of α4βδ GABARs on dendritic spines, as well as on the dendritic shaft, at puberty (Shen et al., 2010a) suggests a role for these receptors in regulating cognition during adolescence, because the dendritic spine is the CNS site for induction of synaptic plasticity. In adult CA1 hippocampus, GABARs localize to the soma (80%) or the dendritic shaft (20%) (Megias et al., 2001), where they can either be sub-synaptic or extrasynaptic, but they are not localized to the dendritic spine, in contrast to cortex where GABAergic innervation of spines is commonly found (Kubata et al., 2007). The predominant extrasynaptic GABARs in the adult are of the subtype α5β3γ2 (Brunig et al., 2002). However, at puberty (~PND 35), extrasynaptic α4βδ expression increases markedly from nearly undetectable levels (Shen et al., 2010a): the proportion of spines which are immunolabeled with α4 and δ is increased by 3 to 6-fold, respectively, while the immunoreactivity per spine is increased 4 to 8-fold, respectively, as determined with silver-intensified immunogold (SIG) labeling and electron microscopy in the proximal stratum radiatum (Fig. 7). It is estimated that 25% of spines may express α4βδ GABARs (Aoki et al., 2012) based on the analysis of immunolabeling. Similar increases in immunolabeling are seen on the dendritic shaft. Pubertal expression of α4βδ GABARs in these dendritic compartments is a transient event, and is reduced significantly by about PND 44 (Aoki et al., 2012), suggesting that high expression of extrasynaptic GABARs in the dendrites persists for a period of about 10 d.
Figure 7. α4 and β GABAA receptor subunit expression increases on dendritic spine membranes of CA1 hippocampal pyramidal cells at puberty.
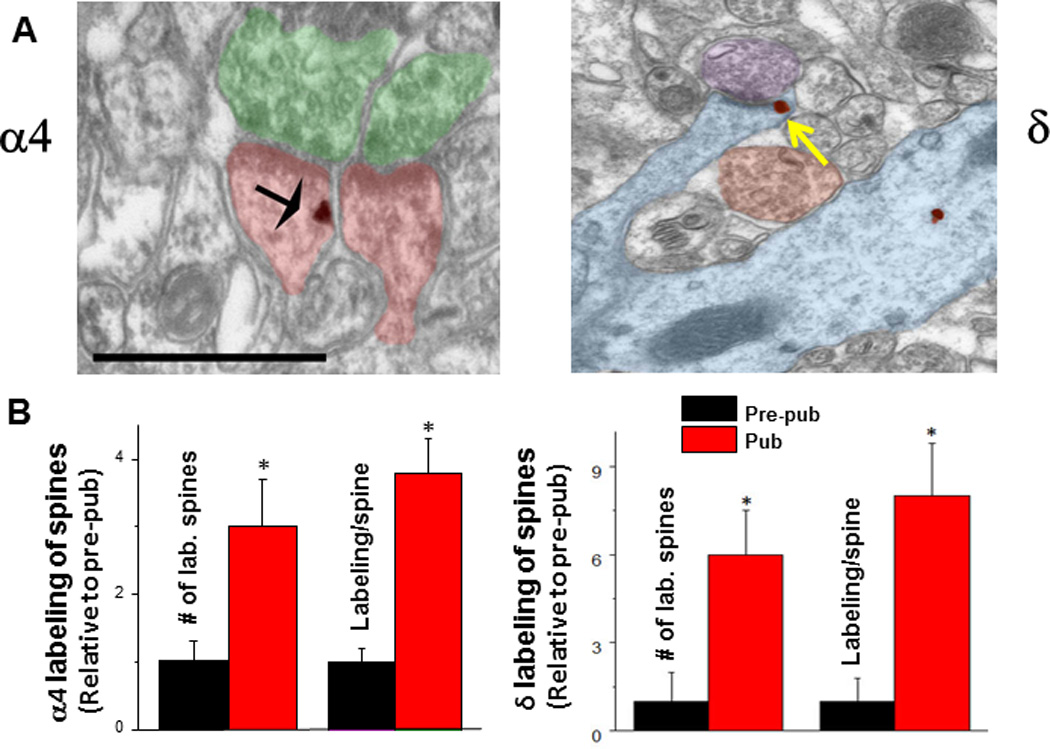
A. α4 (left) and δ (right) silver-intensified immunogold labeling (SIG) occurs along the plasma membrane of spines forming excitatory synapses. α4, black arrow; δ, yellow arrow. Scale, 100 µm. Shafts also exhibit immunoreactivity. B. Averaged data. # of labeled spines (α4, *P < 0.018; δ, **P = 0.002) and labeling per spine (α4, *P < 0.005, δ, **P = 0.00091) increase at puberty (Pub) relative to pre-puberty (Pre-pub). n = 50–80 spines.
α4βδ GABARs activated by ambient GABA produce a tonic inhibition in hippocampus at adolescence (Shen et al., 2007). At this time, activation of NMDA receptors is impaired (Shen et al., 2010a) most likely because the shunting inhibition produced by α4βδ receptors reduces the depolarization necessary for Mg+ unblock of the receptor (Herron et al., 1986). As a consequence, induction of long-term potentiation (LTP), an in vitro model of learning, produced by stimulation of the Schaffer collaterals to CA1 hippocampus with theta burst stimulation is nearly prevented (Shen et al., 2010a). This deficit in synaptic plasticity is not seen with total GABAR blockade or in the δ knock-out mouse, implicating α4βδ GABARs as the mediating factor. In contrast, LTP induction is robust in the hippocampus of pre-pubertal mice. Interestingly, selective blockade of only the synaptic GABARs with 200 nM gabazine does not facilitate LTP induction at puberty, suggesting that the deficit is solely due to the extrasynaptic GABAR population (Shen et al., 2010a). These findings are consistent with an earlier report suggesting that LTP induction is impaired in adolescence due to an increase in GABAergic inhibition (Meredith et al., 2003), although puberty onset and α4βδ were not identified. In contrast, activation of NMDA current is robust in the δ knock-out or after total GABAR blockade (Shen et al., 2010a), suggesting that the deficit in synaptic plasticity at puberty is due to increases in GABA inhibition rather than due to deficits in NMDA receptor expression.
3.3.1 E2 and α4βδ GABAR expression
The onset of puberty in the female is preceded by increases in circulating levels of the ovarian steroid E2, which in the mouse increase two-fold 7–8 days before vaginal opening (Ahima et al., 1997) and in the human appear well before menarche in the pre-pubertal period (Janfaza et al., 2006). Unlike P and THP, E2 levels continue to increase throughout the pubertal period (Ahima et al., 1997; Janfaza et al., 2006), but do not fluctuate (Hodes and Shors, 2005) despite changes in the morphology of cells in the vaginal lavage. The increase in α4βδ GABAR expression at puberty may also be a result of the high levels of circulating E2 because this steroid has been shown to facilitate expression of α4βδ GABARs (Shen et al., 2005). E2 acts via specific nuclear receptors, ERα and ERβ, which govern gene transcription of specific proteins, and these are localized both to the dendritic shaft and spine of hippocampal neurons of female rats and mice (Mitterling et al., 2010). Expression of ERα and ERβ is highest in spines of the CA1 hippocampal pyramidal cells (Mitterling et al., 2010), with ERα most prevalent in the proximal region of the stratum radiatum where expression of α4 and δ has been shown to increase at puberty (Shen et al., 2010a). In fact, expression here is 3 to 4-fold higher than observed in the dentate gyrus (Mitterling et al., 2010) where α4βδ GABAR expression is normally high (Benke et al., 1997; Wisden et al., 1992), suggesting the possibility that E2 may function as a modulator of receptor expression to a greater extent in CA1 hippocampus where α4βδ expression is highly plastic (Smith et al., 2007). Although its role in α4βδ expression is not yet known, E2 increases brain derived neurotrophic factor (BDNF) (Jezierski and Sohrabji, 2003; Sato et al., 2007; Scharfman et al., 2003) which has also been shown to increase activity of the α4 promotor (Roberts et al., 2006) as well as trafficking of δ to the surface membrane (Joshi and Kapur, 2009).
3.3.2 Hippocampal-dependent spatial learning, α4βδ GABARs and adolescence
It is well-known that the hippocampus is the primary site for spatial learning (Bannerman et al., 2004; Burgess et al., 2002; Pastalkova et al., 2006). It would thus be expected that the shunting inhibition produced by α4βδ GABARs on dendritic spines of CA1 pyramidal cells at puberty would impair spatial learning, as was observed. This was determined using a hippocampal-dependent spatial learning task employing active avoidance (Cimadevilla et al., 2001) which is dependent upon LTP (Pastalkova et al., 2006): for the task, mice are trained across three 10-min sessions to avoid a sector on a rotating platform which delivers a mild shock (~0.2 mA). The shock sector is stationary with respect to the room, but rotates with the platform, so that the mouse must move in order to avoid the shock (Fig. 10). (The degree of shock is sub-threshold for stress hormone release (Friedman et al., 1967) suggesting that this is a relatively unstressful task compared to other animal models of learning (Harrison et al., 2009).) For each trial, the latency to enter the shock sector is a measure of learning. Under these conditions, pubertal mice have impaired learning compared to pre-pubertal mice, because they fail to reach criterion (120 s latency to enter the shock sector) after 7 trials (Fig. 10), in contrast to the pre-pubertal mice who reach criterion in 2.5 trials (with a mean latency of 275 s) (Shen et al., 2010b). This learning deficit is not seen in δ knock-out mice at puberty, implicating α4βδ GABARs (Shen et al., 2010a).
Figure 10. Spatial learning is attenuated at puberty: reversal by the stress steroid THP and β knock-out.
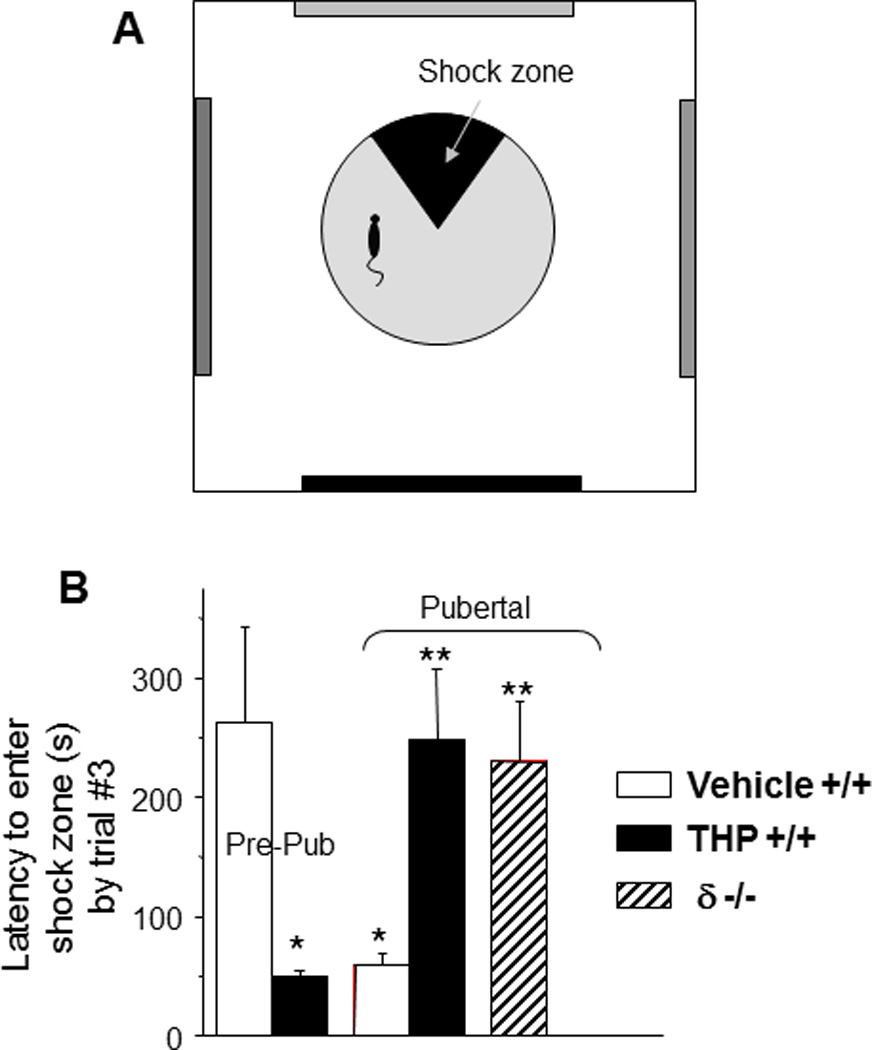
A. Spatial learning platform (shock zone, black sector). The platform rotates, but the shock zone is stationary, requiring the animal to move to actively avoid the shock. B. Latency to first entry of the shock zone, a measure of learning. Pre-pub mice attained the longest entry times. (n= 6 – 9 mice/group) *P<0.05 vs. pre-pub.
3.3.3 The effect of the stress hormone THP on synaptic plasticity at puberty
Because THP reduces hyperpolarizing current at α4βδ GABARs, its increased expression at puberty decreases the shunting tonic inhibition of CA1 hippocampal pyramidal cells (Shen et al., 2010a). This effectively removes the impediment to activation of NMDA receptors, and permits robust activation of NMDA current (Fig. 8), which is also correlated with reversal of the deficit in induction of LTP (Fig. 9), an NMDA receptor-dependent event (Herron et al., 1986). The effect of THP on LTP induction is observed when the steroid is restricted to the dendrites of the stratum radiatum during theta burst stimulation, suggesting that it facilitates LTP exclusively by facilitating induction (Shen et al., 2010a), rather than maintenance of LTP. This was confirmed by a study where THP was applied before induction, washed out and re-applied 5 min after theta burst stimulation, when it did not facilitate LTP. The facilitating effect of THP on LTP was not seen in the δ knock-out, suggesting that α4βδ GABARs are the target for this steroid’s effect on synaptic plasticity at puberty (Shen et al., 2010a).
Figure 8. NMDA current is decreased at puberty in CA1 hippocampal pyramidal cells: reversal by THP and GABAR blockade.
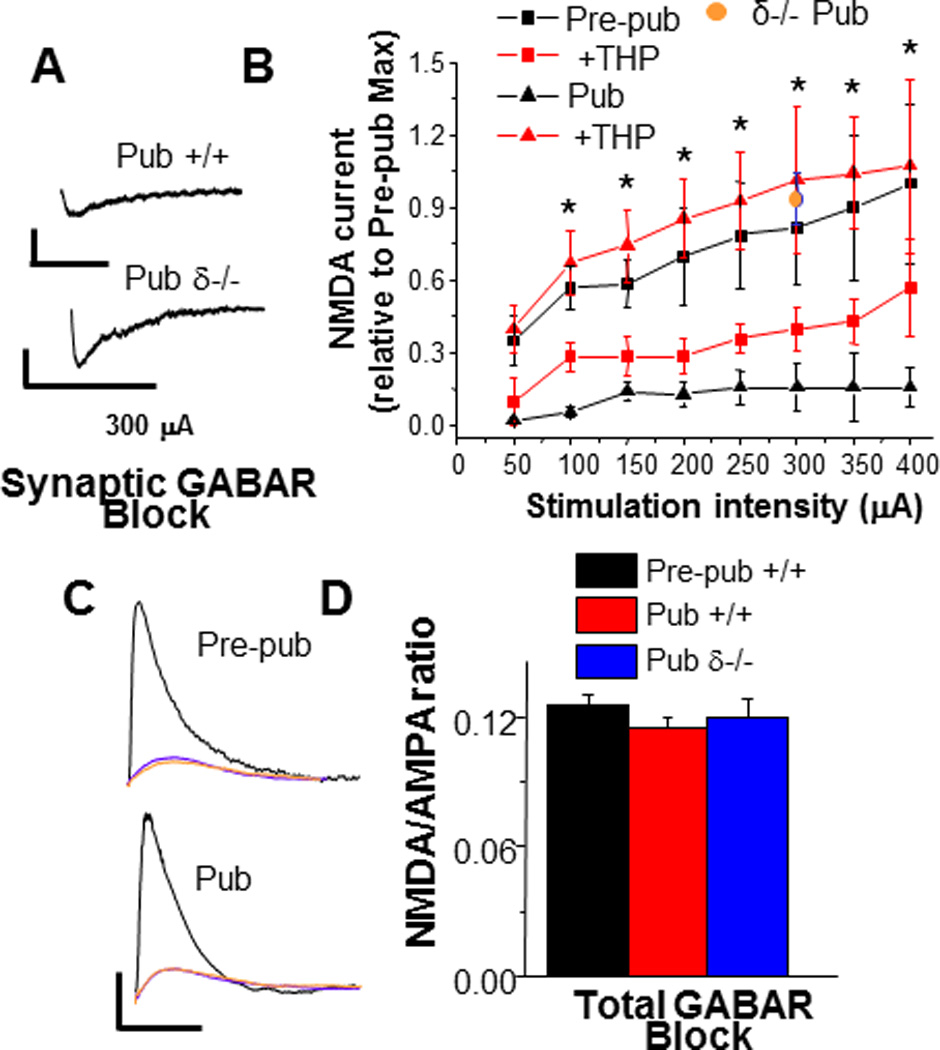
A. Representative traces, evoked (300 µA) NMDA current (0.05 Hz; +/+, above; δ −/−, below) recorded at −60 mV (1 mM MgCl2, bath) with synaptic GABAR blockade (200 nM gabazine) but extrasynaptic GABAR current intact. Scale for +/+, 20 pA, 100 ms; for δ −/−, 20 pA, 250 ms. B. NMDA EPSC amplitude with increasing stimulation intensities (Pre-pub, black squares; +THP, open squares; Pub, black triangle; +THP, open triangle; Pub δ−/−, orange). Amplitudes decreased at puberty, but were restored by THP and δ knock-out.*P < 0.05, Pre-pub versus Pub. C, D: Representative traces and summary data of evoked EPSPs (whole-cell current clamp, black) and NMDA EPSPs (blue; +THP, red) with total GABAR blockade. Scale, 1mV, 100 ms. The NMDA/AMPA ratio was unchanged with total GABAR blockade, suggesting that NMDA receptor expression is not altered by the onset of puberty. (*P < 0.05 versus all other groups, **P < 0.05 versus Pre-pub, Pub). n= 7 – 8 cells/group. (Revised and used with permission (Shen et al., 2010a).)
Figure 9. LTP induction is attenuated at puberty: reversal by the stress steroid THP and β knock-out.
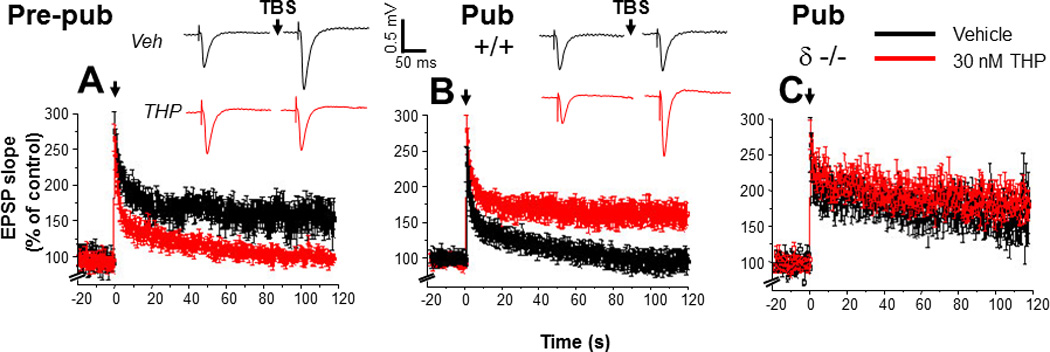
Theta burst stimulation (TBS - 0 time) induced LTP (black) before puberty (Pre-pub, A) but not in the pubertal (Pub, B) CA1 hippocampus. THP (red, 30 nM) permitted LTP induction at puberty. (Inset, Representative field EPSPs. TBS, arrow. Scale, 0.5 mV, 50 ms.) C. LTP, Pubertal δ−/−, where TBS induced LTP. n=6 – 9/group. (Used with permission (Shen et al., 2010a).)
In a similar manner, THP facilitates spatial learning using the hippocampal-dependent active avoidance task (Shen et al., 2010a). After THP administration, the pubertal mice reach criterion in less than 3 trials, with a mean latency of 250 s (Fig. 10), in contrast to their learning deficit in the absence of THP. Facilitating effects of THP on learning are not seen in the δ knock-out mouse, again suggesting that this receptor is the target for THP at puberty (Fig. 11, summary).
Figure 11. Summary of the effects of α4βδ GABARs on CA1 hippocampal pyramidal cells on plasticity at puberty.
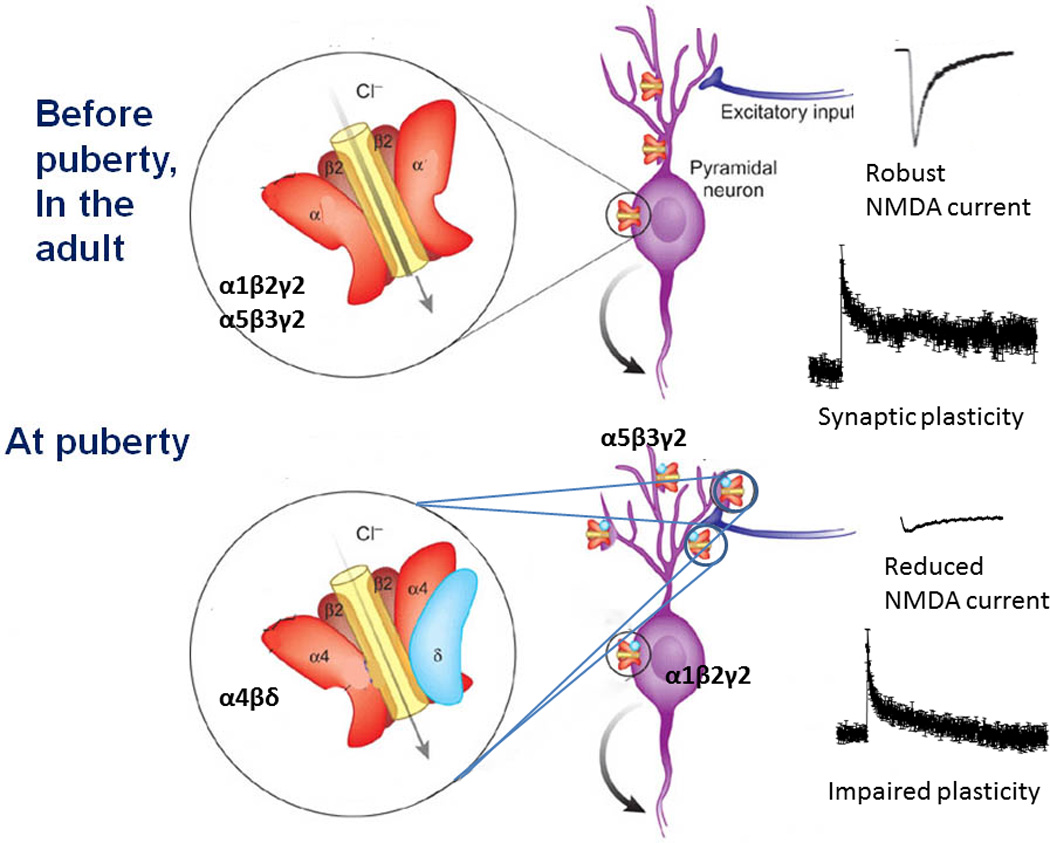
Before puberty and in the adult (upper panel) α4βδ GABAR expression is low. Because existing GABARs do not express on dendritic spines, NMDA receptors are easily activated and induction of LTP, a form of synaptic plasticity, is robust. At puberty (lower panel), α4βδ GABARs emerge on the dendritic spines, producing a shunting inhibition, which impairs NMDA receptor activation and synaptic plasticity. Thus, α4βδ GABAR expression at puberty plays a role in limiting the synaptic plasticity that underlies certain cognitive processes. (Revised and used with permission (McCarthy, 2007).)
3.4 α4βδ GABARs, puberty and eating disorders
Anorexia nervosa and bulimia are eating disorders with restricted or maladaptive patterns of food intake which most commonly begin in adolescence (Kaye, 2008). The disorder is most prevalent in females where the female:male ratio is 7:1 (Kaye, 2008). It has been estimated that, depending on the source, 2 – 7% of adolescent females in the United States have experienced an eating disorder (Kaye, 2008), compared with a 0.5% prevalence in the whole population. Although these disorders can potentially have fatal outcomes (Birmingham et al., 2005), treatment options are limited (Powers and Bruty, 2009). A number of studies have suggested that monoamine disturbance may contribute to these disorders, but the treatment outcomes are frequently equivocal (Powers and Bruty, 2009). One recent study (Aoki et al., 2012) has reported increased expression of α4βδ GABARs in CA1 hippocampus with an activity-based animal model of anorexia nervosa (ABA). In this model, female rats are conditioned during puberty with both food restriction and running wheel activity to mimic conditions underlying anorexia in humans. Expression of α4 and δ on spines of CA1 hippocampal pyramidal cells of control female rats increases at the onset of puberty (PND 32–36), as it does in mice, and remains elevated for about 10 d, declining around PND 42 (Aoki et al., 2012). However, α4 and δ expression remains elevated on dendritic spines of ABA rats, such that, by PND 44, expression is 2 – 6-fold higher than corresponding levels of expression in the control animals (Aoki et al., 2012). In contrast, animals which experience only food restriction or running wheel activity alone do not show increased α4βδ expression on hippocampal spines. This prolonged period of increased expression of α4βδ GABARs on spines has implications for both stress-triggered anxiety and cognitive impairment, as already shown for pubertal female mice (Shen et al., 2007; Shen et al., 2010b). In fact, studies in humans show extensive co-morbidity of anxiety disorders with anorexia nervosa and bulimia (Kaye et al., 2004; Swinbourne et al., 2012). In addition, a recent study has reported decreased cognitive ability in adolescents with anorexia (Andres-Perpina et al., 2011), which is correlated with hyperactivity in several brain regions, but predominantly in the temporal lobe, during a working memory task (Castro-Fornieles et al., 2010). These findings suggest that increased activity in this region may be required to attain normal levels of working memory. The findings are also consistent with increased levels of shunting inhibition in hippocampus in anorexia which would require greater depolarization to activate NMDA receptors, as we have shown in pubertal mice when expression of α4βδ GABARs increases on dendritic spines of CA1 hippocampus (Shen et al., 2010a). Taken together, these findings suggest a link between anxiety and cognitive deficits and eating disorders, which is consistent with increased expression of α4βδ GABARs.
4.1 Conclusions
The onset of puberty demarcates a vulnerable period when mood changes occur and certain cognitive processes can be compromised. The many hormonal events which converge at the onset of puberty may contribute to these changes in mood and cognition that characterize early adolescence. However, the emergence of α4βδ GABARs during early adolescence play a pivotal role in these changes, and act as a target for one stress steroid which can trigger anxiety and alter cognitive processes (Summary Figs. 4,6,11). The outcomes resulting from altered stress responses at puberty are known to contribute to psychopathologies later in development, suggesting that unraveling the cellular mechanisms for these events is an important avenue of investigation.
Highlights.
Puberty onset increases hippocampal expression of α4βδ GABARs.
The stress steroid THP decreases hippocampal tonic inhibitory current at puberty.
THP effects α4βδ GABARs are dependent on the chloride gradient.
THP and restraint stress produce greater anxiety at puberty.
α4βδ expression on CA1 pyramidal cells at puberty reduces synaptic plasticity.
α4βδ GABARs play a role in mood and cognition at puberty in female mice.
Abbreviations
- ABA
activity-based animal model of anorexia nervosa
- AMPA
α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid
- BDNF
brain-derived neurotrophic factor
- CNS
central nervous system
- CRH
corticotrophin releasing hormone
- DHEA
dehydroepiandrosterone
- DHEAS
dehydroepiandrosterone sulfate
- E2
17β-estradiol (an estrogen)
- EPSC/EPSP
excitatory post-synaptic current/potential
- ER
estrogen receptor
- fMRI
functional magnetic resonance imaging
- GABA
γ-amino-butyric-acid
- GABAR
GABAA receptor
- 3α-HSD
3α-hydroxysteroid oxidoreductase
- i.p.
intraperitoneal
- KCC2
K+-Cl− co-transporter
- LTP
long-term potentiation
- NKCC1
Na+-K+-Cl+ co-transporter
- NMDA
N-methyl-D-aspartate
- P
progesterone
- PMDD
premenstrual dysphoric disorder
- PND
post-natal day
- PVN
paraventricular nucleus (hypothalamus)
- SIG
silver-intensified gold
- T
testosterone
- TBS
theta-burst stimulation
- TEA
tetra-ethylammonium
- THDOC
3α,21-dihydroxy-5α-pregnan-20-one
- THP
3α-OH-5[α]β-pregnan-20-one or [allo]pregnanolone
- TTX
tetrodotoxin
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Reference List
- Agis-Balboa RC, Pinna G, Zhubi A, Maloku E, Veldic M, Costa E, Guidotti A. Characterization of brain neurons that express enzymes mediating neurosteroid biosynthesis. Proc Natl Acad Sci. 2006;103:14602–14607. doi: 10.1073/pnas.0606544103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahima RS, Dushay J, Flier SN, Prabakaran D, Flier JS. Leptin accelerates the onset of puberty in normal female mice. J Clin Invest. 1997;99:391–395. doi: 10.1172/JCI119172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alger BE, Nicoll RA. Pharmacological evidence for two kinds of GABA receptors on rat hippocampal pyramidal cells studies in vitro. J Physiol. 1982;328:125–141. doi: 10.1113/jphysiol.1982.sp014256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreen L, Sundstrom-Poromaa I, Bixo M, Andersson A, Nyberg S, Backstrom T. Relationship between allopregnanolone and negative mood in postmenopausal women taking sequential hormone replacement therapy with vaginal progesterone. Psychoneuroendocrinology. 2004;30:212–224. doi: 10.1016/j.psyneuen.2004.07.003. [DOI] [PubMed] [Google Scholar]
- Andres-Perpina S, Lozano-Serra E, Puig O, Lera-Miguel S, Lazaro L, Castro-Fornieles J. Clinical and biological correlates of adolescent anorexia nervosa with impaired cognitive profile. Eur Child Adolesc Psychiatry. 2011;20:541–549. doi: 10.1007/s00787-011-0216-y. [DOI] [PubMed] [Google Scholar]
- Aoki C, Sabaliauskas N, Chowdhury T, Min JY, Colacino AR, Laurino K, Barbarich-Marsteller NC. Adolescent female rats exhibiting activity-based anorexia express elevated levels of GABA(A) receptor alpha4 and delta subunits at the plasma membrane of hippocampal CA1 spines. Synapse. 2012;66:391–407. doi: 10.1002/syn.21528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Apter D. The role of leptin in female adolescence. Ann N Y Acad Sci. 2003;997:64–76. doi: 10.1196/annals.1290.008. [DOI] [PubMed] [Google Scholar]
- Apter D, Hermanson E. Update on female pubertal development. Curr Opinion Obstet Gynecol. 2002;14:475–481. doi: 10.1097/00001703-200210000-00006. [DOI] [PubMed] [Google Scholar]
- Ardila A, Rosselli M, Matute E, Inozemtseva O. Gender differences in cognitive development. Dev Psychol. 2011;47:984–990. doi: 10.1037/a0023819. [DOI] [PubMed] [Google Scholar]
- Bai D, Zhu G, Pennefather P, Jackson MF, Macdonald JF, Orser BA. Distinct functional and pharmacological properties of tonic and quantal inhibitory postsynaptic currents mediated by y-aminobutyric acid(A) receptors in hippocampal neurons. Molec Pharmac. 2000;59:814–824. doi: 10.1124/mol.59.4.814. [DOI] [PubMed] [Google Scholar]
- Bailey J, Penhune VB. A sensitive period for musical training: contributions of age of onset and cognitive abilities. Ann N Y Acad Sci. 2012;1252:163–170. doi: 10.1111/j.1749-6632.2011.06434.x. [DOI] [PubMed] [Google Scholar]
- Bannerman DM, Rawlins JNP, McHugh SB, Deacon RMJ, Yee BK, Bast T, Zhang W-N, Pothuizen HHJ, Feldon J. Regional dissociations within the hippocampus--memory and anxiety. Neurosci Biobehav Rev. 2004;28:273–283. doi: 10.1016/j.neubiorev.2004.03.004. [DOI] [PubMed] [Google Scholar]
- Barbaccia ML, Serra M, Purdy RH, Biggio G. Stress and neuroactive steroids. Int Rev Neurobiol. 2001;46:243–272. doi: 10.1016/s0074-7742(01)46065-x. [DOI] [PubMed] [Google Scholar]
- Belelli D, Casula A, Ling A, Lambert JJ. The influence of subunit composition on the interaction of neurosteroids with GABA(A) receptors. Neuropharm. 2002;43:651–661. doi: 10.1016/s0028-3908(02)00172-7. [DOI] [PubMed] [Google Scholar]
- Belelli D, Herd MB. The contraceptive agent Provera enhances GABA(A) receptor-mediated inhibitory neurotransmission in the rat hippocampus: evidence for endogenous neurosteroids? J Neurosci. 2003;23:10013–10020. doi: 10.1523/JNEUROSCI.23-31-10013.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belelli D, Peden DR, Rosahl TW, Wafford K, Lambert JJ. Extrasynaptic GABA-A receptors for thalamocortical neurons: A molecular target for hypnotics. J Neurosci. 2005;25:11513–11520. doi: 10.1523/JNEUROSCI.2679-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belelli D, Harrison NL, Maguire J, Macdonald RL, Walker MC, Cope DW. Extrasynaptic GABA-A receptors: form, pharmacology, and function. J Neurosci. 2009;29:12757–12763. doi: 10.1523/JNEUROSCI.3340-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellani R, Luecken LJ, Conrad CD. Peripubertal anxiety profile can predict predisposition to spatial memory impairments following chronic stress. Behav Brain Res. 2006;166:263–270. doi: 10.1016/j.bbr.2005.08.006. [DOI] [PubMed] [Google Scholar]
- Benke D, Michel C, Mohler H. GABA-A receptors containing the a4-subunit: prevalence, distribution, pharmacology, and subunit architecture in situ. J Neurochem. 1997;69:806–814. doi: 10.1046/j.1471-4159.1997.69020806.x. [DOI] [PubMed] [Google Scholar]
- Bentley PJ. Comparative Vertebrate Endocrinology. Cambridge: Cambridge University Press; 1998. [Google Scholar]
- Bianchi MT, Haas KF, Macdonald RL. Alpha1 and alpha6 subunits specify distinct desensitization, deactivation and neurosteroid modulation of GABA(A) receptors containing the delta subunit. Neuropharm. 2002;43:492–502. doi: 10.1016/s0028-3908(02)00163-6. [DOI] [PubMed] [Google Scholar]
- Bianchi MT, Macdonald RL. Neurosteroids shift partial agonist activation of GABA(A) receptor channels from low-to high-efficacy gating patterns. J Neurosci. 2003;23:10934–10943. doi: 10.1523/JNEUROSCI.23-34-10934.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birmingham CL, Su J, Hlynsky JA, Goldner EM, Gao M. The mortality rate from anorexia nervosa. Int J Eat Disord. 2005;38:143–146. doi: 10.1002/eat.20164. [DOI] [PubMed] [Google Scholar]
- Bitran D, Dugan M, Renda P, Ellis R, Foley M. Anxiolytic effects of the neuroactive steroid pregnanolone (3alpha-OH-5beta-pregnan-20-one) after microinjection in the dorsal hippocampus and lateral septum. Brain Res. 1999;850:217–224. doi: 10.1016/s0006-8993(99)02150-2. [DOI] [PubMed] [Google Scholar]
- Blanton RE, Cooney RE, Joormann J, Eugene F, Glover GH, Gotlib IH. Pubertal stage and brain anatomy in girls. Neuroscience. 2012;217:105–112. doi: 10.1016/j.neuroscience.2012.04.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowers JM, Waddell J, McCarthy MM. A developmental sex difference in hippocampal neurogenesis is mediated by endogenous oestradiol. Biol Sex Differ. 2010;1:8. doi: 10.1186/2042-6410-1-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brack KE, Lovick TA. Neuronal excitability in the periaqueductal grey matter during the estrous cycle in female Wistar rats. Neuroscience. 2007;144:325–335. doi: 10.1016/j.neuroscience.2006.08.058. [DOI] [PubMed] [Google Scholar]
- Bright DP, Renzi M, Bartram J, McGee TP, MacKenzie G, Hosie AM, Farrant M, Brickley SG. Profound desensitization by ambient GABA limits activation of delta-containing GABAA receptors during spillover. J Neurosci. 2011;31:753–763. doi: 10.1523/JNEUROSCI.2996-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown N, Kerby J, Bonnert TP, Whiting PJ, Wafford KA. Pharmacological characterization of a novel cell line expressing human alpha(4)beta(3)delta GABA(A) receptors. Br J Pharmacol. 2002;136:965–974. doi: 10.1038/sj.bjp.0704795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brunig I, Scotti E, Sidler C, Fritschy JM. Intact sorting, targeting, and clustering of gamma-aminobutyric acid A receptor subtypes in hippocampal neurons in vitro. J.Comp.Neurol. 2002;443:43–55. doi: 10.1002/cne.10102. [DOI] [PubMed] [Google Scholar]
- Buchanan CM, Eccles JS, Becker JB. Are adolescents the victims of raging hormones: evidence for activational effects of hormones on moods and behavior at adolescence. Psychol Bull. 1992;111:62–107. doi: 10.1037/0033-2909.111.1.62. [DOI] [PubMed] [Google Scholar]
- Burgard EC, Tietz EI, Neelands TR, Macdonald RL. Properties of recombinant gamma-aminobutyric acid A receptor isoforms containing the alpha 5 subunit subtype. Mol Pharmacol. 1996;50:119–127. [PubMed] [Google Scholar]
- Burgess N, Maguire EA, O'Keefe J. The human hippocampus and spatial and episodic memory. Neuron. 2002;35:625–641. doi: 10.1016/s0896-6273(02)00830-9. [DOI] [PubMed] [Google Scholar]
- Callachan H, Cottrell GA, Hather NY, Lambert JJ, Nooney JM, Peters JA. Modulation of the GABAA receptor by progesterone metabolites. Proc R Soc Lond B Biol Sci. 1987;231:359–369. doi: 10.1098/rspb.1987.0049. [DOI] [PubMed] [Google Scholar]
- Caraiscos VB, Elliott EM, You-Ten KE, Cheng VY, Belelli D, Newell JG, Jackson MF, Lambert JJ, Rosahl TW, Wafford K, MacDonald JF, Orser BA. Tonic inhibition in mouse hippocampal CA1 pyramidal neurons is mediated by alpha 5 subunit-containing gamma-aminobutyric acid type A receptors. Proc Natl Acad Sci. 2004;101:3662–3667. doi: 10.1073/pnas.0307231101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrion VG, Weems CF, Ray R, Reiss AL. Toward an empirical definition of pediatric PTSD: the phenomenology of PTSD symptoms in youth. J Am Acad Child Adolesc Psychiatry. 2002;41:166–173. doi: 10.1097/00004583-200202000-00010. [DOI] [PubMed] [Google Scholar]
- Castro-Fornieles J, Caldu X, Andres-Perpina S, Lazaro L, Bargallo N, Falcon C, Plana MT, Junque C. A cross-sectional and follow-up functional MRI study with a working memory task in adolescent anorexia nervosa. Neuropsychologia. 2010;48:4111–4116. doi: 10.1016/j.neuropsychologia.2010.10.003. [DOI] [PubMed] [Google Scholar]
- Chambers MS, Atack JR, Broughton HB, Collinson N, Cook S, Dawson GR, Hobbs SC, Marshall G, Maubach KA, Pillai GV, Reeve AJ, MacLeod AM. Identification of a novel, selective GABA(A) alpha-5 receptor inverse agonist which enhances cognition. J.Med.Chem. 2003;46:2227–2240. doi: 10.1021/jm020582q. [DOI] [PubMed] [Google Scholar]
- Chandra D, Jia F, Liang J, Peng Z, Suryanarayanan A, Werner DF, Spigelman I, Houser CR, Olsen RW, Harrison NL, Homanics GE. GABA-A receptor alpha-4 subunits mediate extrasynaptic inhibition in thalamus and dentate gyrus and the action of gaboxadol. Proc Natl Acad Sci. 2006;103:15230–15235. doi: 10.1073/pnas.0604304103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang Y, Wang R, Barot S, Weiss DS. Stoichiometry of a recombinant GABA-A receptor. J Neurosci. 1990;16:534–541. doi: 10.1523/JNEUROSCI.16-17-05415.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng VY, Martin LJ, Elliott EM, Kim JH, Mount HT, Taverna FA, Roder JC, MacDonald JF, Bhambri A, Collinson N, Wafford K, Orser BA. Alpha5GABAA receptors mediate the amnestic but not sedative-hypnotic effects of the general anesthetic etomidate. J Neurosci. 2006;26:3713–3720. doi: 10.1523/JNEUROSCI.5024-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cimadevilla JM, Wesierska M, Fenton AA, Bures J. Inactivating one hippocampus impairs avoidance of a stable room-defined place during dissociaiton of arena cues from room cues by rotation of the arena. Proc Natl Acad Sci. 2001;98:3532–3536. doi: 10.1073/pnas.051628398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collinson N, Kuenzi FM, Jarolimek W, Maubach KA, Cothliff R, Sur C, Smith A, Otu FM, Jowell O, Atack JR, McKernan RM, Seabrook GR, Dawson GR, Whiting PJ, Rosahl TW. Enhanced learning and memory and altered GABAergic synaptic transmission in mice lacking the alpha 5 subunit of the GABA-A receptor. J.Neurosci. 2002;22:5572–5580. doi: 10.1523/JNEUROSCI.22-13-05572.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Compagnone NA, Mellon SH. Neurosteroids: Biosynthesis and Function of These Novel Neuromodulators. Frontiers in Neuroendocrinology. 2000;21:1–56. doi: 10.1006/frne.1999.0188. [DOI] [PubMed] [Google Scholar]
- Corcoran CM, Smith C, McLaughlin D, Auther A, Malaspina D, Cornblatt B. HPA axis function and symptoms in adolescents at clinical high risk for schizophrenia. Schizophr Res. 2012;135:170–174. doi: 10.1016/j.schres.2011.11.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costello EJ, Mustillo S, Erkanli A, Keeler G, Angold A. Prevalence and development of psychiatric disorders in childhood and adolescence. Arch Gen Psychiatry. 2003;60:837–844. doi: 10.1001/archpsyc.60.8.837. [DOI] [PubMed] [Google Scholar]
- Crestani F, Keist R, Fritschy J-M, Benke D, Vogt K, Prut L, Bluthmann H, Mohler H, Rudolph U. Trace fear conditioning involves hippocampal alpha-5 GABA(A) receptors. Proc Natl Acad Sci. 2002;99:8980–8985. doi: 10.1073/pnas.142288699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham J, Yonkers KA, O'Brien S, Eriksson E. Update on research and treatment of premenstrual dysphoric disorder. Harv Rev Psychiatry. 2009;17:120–137. doi: 10.1080/10673220902891836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dahl RE. Adolescent brain development: a period of vulnerabilities and opportunities. Keynote address. Ann N Y Acad Sci. 2004;1021:1–22. doi: 10.1196/annals.1308.001. [DOI] [PubMed] [Google Scholar]
- Davidson RJ, McEwen BS. Social influences on neuroplasticity: stress and interventions to promote well-being. Nat Neurosci. 2012;15:689–695. doi: 10.1038/nn.3093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dorn LD, Chrousos GP. The neurobiology of stress: understanding regulation of affect during female biological transitions. Semin Reprod Endocrinol. 1997;15:19–35. doi: 10.1055/s-2008-1067965. [DOI] [PubMed] [Google Scholar]
- Droogleever Fortuyn HA, van Broekhoven F, Span PN, Backstrom T, Zitman FG, Verkes RJ. Effects of PhD examination stress on allopregnanolone and cortisol plasma levels and peripheral benzodiazepine receptors density. Psychoneuroendocrinol. 2004;29:1341–1344. doi: 10.1016/j.psyneuen.2004.02.003. [DOI] [PubMed] [Google Scholar]
- Dunn AJ, Swiergiel AH. Behavioral responses to stress are intact in CRF-deficient mice. Brain Res. 1999;845:14–20. doi: 10.1016/s0006-8993(99)01912-5. [DOI] [PubMed] [Google Scholar]
- Dzhala V, Valeeva G, Glykys J, Khazipov R, Staley K. Traumatic alterations in GABA signaling disrupt hippocampal network activity in the developing brain. J Neurosci. 2012;32:4017–4031. doi: 10.1523/JNEUROSCI.5139-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ewart CK, Kolodner KB. Social competence interview for assessing physiological reactivity in adolescents. Psychosom Med. 1991;53:289–304. doi: 10.1097/00006842-199105000-00003. [DOI] [PubMed] [Google Scholar]
- Fadalti M, Petraglia F, Luisi S, Bernardi F, Casarosa E, Ferrari E, Luisi M, Saggese G, Genazzani AR, Bernasconi S. Changes of serum allopregnanolone levels in the first 2 years of life and during pubertal development. Pediatr Res. 1999;46:323–327. doi: 10.1203/00006450-199909000-00013. [DOI] [PubMed] [Google Scholar]
- Freeman EW, Frye CA, Rickels K, Martin PA, Smith SS. Allopregnanolone levels and symptom improvement in severe premenstrual syndrome. J Clin Psychopharmacol. 2002;22:516–520. doi: 10.1097/00004714-200210000-00013. [DOI] [PubMed] [Google Scholar]
- Friedman SB, Ader R, Grota LJ, Larson T. Plasma corticosterone response to parameters of electric shock stimulation in the rat. Psychosomatic Med. 1967;29:323–328. doi: 10.1097/00006842-196707000-00003. [DOI] [PubMed] [Google Scholar]
- Frye CA. The neurosteroid 3alpha,5alpha-THP has antiseizure and possible neuroprotective effects in an animal model of epilepsy. Brain Res. 1995;696:113–120. doi: 10.1016/0006-8993(95)00793-p. [DOI] [PubMed] [Google Scholar]
- Frye CA, Duffy CK, Walf AA. Estrogens and progestins enhance spatial learning of intact and ovariectomized rats in the object placement task. Neurobiol Learn Mem. 2007;88:208–216. doi: 10.1016/j.nlm.2007.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frye CA, Walf AA. Depression-like behavior of aged male and female mice is ameliorated with administration of testosterone or its metabolites. Physiol Behav. 2009a;97:266–269. doi: 10.1016/j.physbeh.2009.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frye CA, Walf AA. Progesterone reduces depression-like behavior in a murine model of Alzheimer's Disease. Age (Dordr) 2009b;31:143–153. doi: 10.1007/s11357-009-9091-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garber J, Keiley MK, Martin C. Developmental trajectories of adolescents' depressive symptoms: predictors of change. J Consult Clin Psychol. 2002;70:79–95. doi: 10.1037//0022-006x.70.1.79. [DOI] [PubMed] [Google Scholar]
- Ge Y, Dong Z, Bagot RC, Howland JG, Phillips AG, Wong TP, Wang YT. Hippocampal long-term depression is required for the consolidation of spatial memory. Proc Natl Acad Sci U S A. 2010;107:16697–16702. doi: 10.1073/pnas.1008200107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gee KW, Chang W-C, Brinton RE, McEwen BS. GABA-dependent modulation of the Clionophore by steroids in rat brain. Eur J Pharmac. 1987;136:419–423. doi: 10.1016/0014-2999(87)90317-7. [DOI] [PubMed] [Google Scholar]
- Girdler SS, Beth Mechlin M, Light KC, Morrow AL. Ethnic differences in allopregnanolone concentrations in women during rest and following mental stress. Psychophysiology. 2006;43:331–336. doi: 10.1111/j.1469-8986.2006.00410.x. [DOI] [PubMed] [Google Scholar]
- Griffiths J, Lovick T. Withdrawal from progesterone increases expression of alpha4, beta1 and delta GABA(A) receptor subunits in neurons in the periaqueductal gray matter in female Wistar rats. J Comp Neurol. 2005a;486:89–97. doi: 10.1002/cne.20540. [DOI] [PubMed] [Google Scholar]
- Griffiths JL, Lovick TA. GABAergic neurones in the rat periaqueductal grey matter express alpha4, beta1 and delta GABAA receptor subunits: plasticity of expression during the estrous cycle. Neuroscience. 2005b;136:457–466. doi: 10.1016/j.neuroscience.2005.08.013. [DOI] [PubMed] [Google Scholar]
- Gur RC, Richard J, Calkins ME, Chiavacci R, Hansen JA, Bilker WB, Loughead J, Connolly JJ, Qiu H, Mentch FD, Abou-Sleiman PM, Hakonarson H, Gur RE. Age group and sex differences in performance on a computerized neurocognitive battery in children age 8–21. Neuropsychology. 2012;26:251–265. doi: 10.1037/a0026712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guyer AE, McClure-Tone EB, Shiffrin ND, Pine DS, Nelson EE. Probing the neural correlates of anticipated peer evaluation in adolescence. Child Dev. 2009;80:1000–1015. doi: 10.1111/j.1467-8624.2009.01313.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haas KF, Macdonald RL. GABAA receptor subunit gamma2 and delta subtypes confer unique kinetic properties on recombinant GABAA receptor currents in mouse fibroblasts. J Physiol. 1999;514:27–45. doi: 10.1111/j.1469-7793.1999.027af.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison FE, Hosseini AH, McDonald MP. Endogenous anxiety and stress responses in water maze and Barnes maze spatial memory tasks. Behav Brain Res. 2009;198:247–251. doi: 10.1016/j.bbr.2008.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison NL, Majewska MD, Harrington JW, Barker JL. Structure-activity relationships for steroid interaction with a g-aminobutyric acidA receptor complex. J Pharmacol Exp Ther. 1987;241:346–353. [PubMed] [Google Scholar]
- Hayward C, Sanborn K. Puberty and the emergence of gender diferences in psychopathology. J Adolescent Health. 2002;30S:49–58. doi: 10.1016/s1054-139x(02)00336-1. [DOI] [PubMed] [Google Scholar]
- Hayward C, Killen JD, Hammer LD, Litt IF, Wilson DM, Simmonds B, Taylor CB. Pubertal stage and panic attack history in sixth- and seventh-grade girls. Am J Psychiatry. 1992;149:1239–1243. doi: 10.1176/ajp.149.9.1239. [DOI] [PubMed] [Google Scholar]
- Heim C, Plotsky PM, Nemeroff CB. Importance of studying the contributions of early adverse experience to neurobiological findings in depression. Neuropsychopharmacology. 2004;29:641–648. doi: 10.1038/sj.npp.1300397. [DOI] [PubMed] [Google Scholar]
- Herd MB, Haythornthwaite AR, Rosahl TW, Wafford K, Homanics GE, Lambert JJ, Belelli D. The expression of GABA-A Beta subunit isoforms in synaptic and extrasynaptic receptor populations of mouse dentate gyrus granule cells. J Physiol. 2008;586:989–1004. doi: 10.1113/jphysiol.2007.146746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herron CE, Lester RA, Coan EJ, Collingridge GL. Frequency-dependent involvement of NMDA receptors in the hippocampus: a novel synaptic mechanism. Nature. 1986;322:265–268. doi: 10.1038/322265a0. [DOI] [PubMed] [Google Scholar]
- Herting MM, Nagel BJ. Aerobic fitness relates to learning on a virtual Morris Water Task and hippocampal volume in adolescents. Behav Brain Res. 2012;233:517–525. doi: 10.1016/j.bbr.2012.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higashi T, Takido N, Shimada K. Studies on neurosteroids XVII. Analysis of stress-induced changes in neurosteroid levels in rat brains using liquid chromatography-electron capture atmosperic pressure chemical ionization-mass spectrometry. Steroids. 2005;70:1–11. doi: 10.1016/j.steroids.2004.08.001. [DOI] [PubMed] [Google Scholar]
- Hodes GE, Shors TJ. Distinctive stress effects on learning during puberty. Horm Behav. 2005;48:163–171. doi: 10.1016/j.yhbeh.2005.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosie AM, Wilkins ME, da Silva HM, Smart TG. Endogenous neurosteroids regulate GABA-A receptors through two discrete transmembrane sites. Nature. 2006;444:486–489. doi: 10.1038/nature05324. [DOI] [PubMed] [Google Scholar]
- Houston CM, Bright DP, Sivilotti LG, Beato M, Smart TG. Intracellular chloride ions regulate the time course of GABA-mediated inhibitory synaptic transmission. J Neurosci. 2009;29:10416–10423. doi: 10.1523/JNEUROSCI.1670-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huerta R, Brizuela-Gamino OL. Interaction of pubertal status, mood and self-esteem in adolescent girls. J Reprod Med. 2002;47:217–225. [PubMed] [Google Scholar]
- Imfeld A, Oechslin MS, Meyer M, Loenneker T, Jancke L. White matter plasticity in the corticospinal tract of musicians: a diffusion tensor imaging study. Neuroimage. 2009;46:600–607. doi: 10.1016/j.neuroimage.2009.02.025. [DOI] [PubMed] [Google Scholar]
- Isomura Y, Sugimoto M, Fujiwara-Tsukamoto Y, Yamamoto-Muraki S, Yamada J, Fukuda A. Synaptically activated Cl− accumulation responsible for depolarizing GABAergic responses in mature hippocampal neurons. J Neurophysiol. 2003;90:2752–2756. doi: 10.1152/jn.00142.2003. [DOI] [PubMed] [Google Scholar]
- Izumi Y, Nagashima K, Murayama K, Zorumski CF. Acute effects of ethanol on hippocampal long-term potentiation and long-term depression are mediated by different mechanisms. Neuroscience. 2005;136:509–517. doi: 10.1016/j.neuroscience.2005.08.002. [DOI] [PubMed] [Google Scholar]
- Janfaza M, Sherman TI, Larmore KA, Brown-Dawson J, Klein KO. Estradiol levels and secretory dynamics in normal girls and boys as determined by an ultrasensitive bioassay: a 10 year experience. J Pediatr Endocrinol Metab. 2006;19:901–909. doi: 10.1515/jpem.2006.19.7.901. [DOI] [PubMed] [Google Scholar]
- Jezierski MK, Sohrabji F. Estrogen enhances retrograde transport of brain-derived neurotrophic factor in the rodent forebrain. Endocrinology. 2003;144:5022–5029. doi: 10.1210/en.2003-0724. [DOI] [PubMed] [Google Scholar]
- Johnson JS, Newport EL. Critical period effects in second language learning: The influence of maturational state on the acquisition of English as a second language. Cognitive Psychology. 1989;21:60–99. doi: 10.1016/0010-0285(89)90003-0. [DOI] [PubMed] [Google Scholar]
- Jones MV, Westbrook GL. Desensitized states prolong GABAA channel responses to brief agonist pulses. Neuron. 1995;15:181–191. doi: 10.1016/0896-6273(95)90075-6. [DOI] [PubMed] [Google Scholar]
- Joshi S, Kapur J. Slow intracellular accumulation of GABA(A) receptor delta subunit is modulated by brain-derived neurotrophic factor. Neuroscience. 2009;164:507–519. doi: 10.1016/j.neuroscience.2009.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanit L, Taskiran D, Yilmaz OA, Balkan B, Demirgoren S, Furedy JJ, Pogun S. Sexually dimorphic cognitive style in rats emerges after puberty. Brain Res Bull. 2000;52:243–248. doi: 10.1016/s0361-9230(00)00232-x. [DOI] [PubMed] [Google Scholar]
- Karavolas HJ, Hodges DR. Neuroendocrine metabolism of progesterone and related progestins. Ciba Found Symp. 1990;153:22–44. doi: 10.1002/9780470513989.ch3. [DOI] [PubMed] [Google Scholar]
- Kask K, Backstrom T, Lundgren P, Sundstrom PI. Allopregnanolone has no effect on startle response and prepulse inhibition of startle response in patients with premenstrual dysphoric disorder or healthy controls. Pharmacol Biochem Behav. 2009;92:608–613. doi: 10.1016/j.pbb.2009.02.014. [DOI] [PubMed] [Google Scholar]
- Kaye W. Neurobiology of anorexia and bulimia nervosa. Physiol Behav. 2008;94:121–135. doi: 10.1016/j.physbeh.2007.11.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaye WH, Bulik CM, Thornton L, Barbarich N, Masters K. Comorbidity of anxiety disorders with anorexia and bulimia nervosa. Am J Psychiatry. 2004;161:2215–2221. doi: 10.1176/appi.ajp.161.12.2215. [DOI] [PubMed] [Google Scholar]
- Kelly MM, Tyrka AR, Anderson GM, Price LH, Carpenter LL. Sex differences in emotional and physiological responses to the Trier Social Stress Test. J Behav Ther Exp Psychiatry. 2008;39:87–98. doi: 10.1016/j.jbtep.2007.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kessler RC, Berglund P, Demler O, Jin R, Merikangas KR, Walters EE. Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey Replication. Arch Gen Psychiatry. 2005;62:593–602. doi: 10.1001/archpsyc.62.6.593. [DOI] [PubMed] [Google Scholar]
- Khan A, Berger RG, deCatanzaro D. The onset of puberty in female mice as reflected in urinary steroids and uterine/ovarian mass: interactions of exposure to males, phyto-oestrogen content of diet, and ano-genital distance. Reproduction. 2008;135:99–106. doi: 10.1530/REP-07-0314. [DOI] [PubMed] [Google Scholar]
- Kral A, Hartmann R, Tillein J, Heid S, Klinke R. Delayed maturation and sensitive periods in the auditory cortex. Audiol Neurootol. 2001;6:346–362. doi: 10.1159/000046845. [DOI] [PubMed] [Google Scholar]
- Kubota Y, Hatada S, Kondo S, Karube F, Kawaguchi Y. Neocortical inhibitory terminals innervate dendritic spines targeted by thalamocortical afferents. J Neurosci. 2007;27:1139–1150. doi: 10.1523/JNEUROSCI.3846-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumaran D, Maguire EA. An unexpected sequence of events: Mismatch detection in the human hippocampus. PLOS Biology. 2006;4:2372–2382. doi: 10.1371/journal.pbio.0040424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuver A, Shen H, Smith SS. Regulation of the surface expression of alpha4beta2delta GABA(A) receptors by high efficacy states. Brain Res. 2012;1463:1–20. doi: 10.1016/j.brainres.2012.04.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambert NA, Borroni AM, Grover LM, Teyler TJ. Hyperpolarizing and depolarizing GABA(A) receptor-mediated dendritic inhibition in area CA1 of the rat hippocampus. J.Neurophysiol. 1991;66:1538–1548. doi: 10.1152/jn.1991.66.5.1538. [DOI] [PubMed] [Google Scholar]
- Lancel M, Faulhaber J, Schiffelholz T, Romeo E, Di Michele F, Holsboer F, Rupprecht R. Allopregnanolone affects sleep in a benzodiazepine-like fashion. J Pharmacol Exp Ther. 1997;282:1213–1218. [PubMed] [Google Scholar]
- Leen-Feldner EW, Reardon LE, Zvolensky MJ. Pubertal status and emotional reactivity to a voluntary hyperventilation challenge predicting panic symptoms and somatic complaints: a laboratory-based multi-informant test. Behav Modif. 2007;31:8–31. doi: 10.1177/0145445506295058. [DOI] [PubMed] [Google Scholar]
- Lenroot RK, Giedd JN. Sex differences in the adolescent brain. Brain Cogn. 2010;72:46–55. doi: 10.1016/j.bandc.2009.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang SW, Jemerin JM, Tschann JM, Irwin CE, Jr, Wara DW, Boyce WT. Life events, cardiovascular reactivity, and risk behavior in adolescent boys. Pediatrics. 1995;96:1101–1105. [PubMed] [Google Scholar]
- Lovick T. Panic disorder: a malfunction of multiple transmitter control systems within the midbrain periaqueductal gray matter? Neuroscientist. 2000;6:48–59. [Google Scholar]
- Lovick TA, Griffiths JL, Dunn SM, Martin IL. Changes in GABA(A) receptor subunit expression in the midbrain during the oestrous cycle in Wistar rats. Neuroscience. 2005a;131:397–405. doi: 10.1016/j.neuroscience.2004.11.010. [DOI] [PubMed] [Google Scholar]
- Lovick TA, Griffiths JL, Dunn SM, Martin IL. Changes in GABA(A) receptor subunit expression in the midbrain during the oestrous cycle in Wistar rats. Neuroscience. 2005b;131:397–405. doi: 10.1016/j.neuroscience.2004.11.010. [DOI] [PubMed] [Google Scholar]
- Lui P, Padow VA, Franco D, Hall BS, Park B, Klein ZA, Romeo RD. Divergent stress-induced neuroendocrine and behavioral responses prior to puberty. Physiol Behav. 2012;107:104–111. doi: 10.1016/j.physbeh.2012.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lupien SJ, Maheu F, Tu M, Fiocco A, Schramek TE. The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition. Brain Cogn. 2007;65:209–237. doi: 10.1016/j.bandc.2007.02.007. [DOI] [PubMed] [Google Scholar]
- Maconochie DJ, Zempel JM, Steinbach JH. How quickly can GABAA receptors open? Neuron. 1994;12:61–71. doi: 10.1016/0896-6273(94)90152-x. [DOI] [PubMed] [Google Scholar]
- Maguire EA, Frackowiak RSJ, Firth CD. Recalling routes around London: Activation of the right hippocampus in taxi drivers. J Neurosci. 1997;17:7103–7110. doi: 10.1523/JNEUROSCI.17-18-07103.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maguire JL, Ferando I, Simonsen C, Mody I. Excitability changes related to GABA-A receptor plasticity during pregnancy. J Neurosci. 2009;29:9592–9601. doi: 10.1523/JNEUROSCI.2162-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maguire JL, Mody I. Neurosteroid synthesis-mediated regulation of GABA(A) receptors: relevance to the ovarian cycle and stress. J Neurosci. 2007;27:2155–2162. doi: 10.1523/JNEUROSCI.4945-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maguire JL, Mody I. GABA(A)R plasticity during pregnancy: relevance to postpartum depression. Neuron. 2009;59:207–213. doi: 10.1016/j.neuron.2008.06.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maguire JL, Stell BM, Rafizadeh M, Mody I. Ovarian cycle-linked changes in GABA(A) receptors mediating tonic inhibition alter seizure susceptibility and anxiety. Nat Neurosci. 2005;8:797–804. doi: 10.1038/nn1469. [DOI] [PubMed] [Google Scholar]
- Majewska MD, Harrison NL, Schwartz RD, Barker JL, Paul SM. Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor. Science. 1986;232:1004–1007. doi: 10.1126/science.2422758. [DOI] [PubMed] [Google Scholar]
- Maninger N, Wolkowitz OM, Reus VI, Epel ES, Mellon SH. Neurobiological and neuropsychiatric effects of dehydroepiandrosterone (DHEA) and DHEA sulfate (DHEAS) Front Neuroendocrinol. 2009;30:65–91. doi: 10.1016/j.yfrne.2008.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mannan MA, O'Shaughnessy PJ. Ovarian steroid metabolism during post-natal development in the normal mouse and in the adult hypogonadal (hpg) mouse. J Reprod Fertil. 1988;82:727–734. doi: 10.1530/jrf.0.0820727. [DOI] [PubMed] [Google Scholar]
- Matthews DB, Morrow AL, Tokunaga S, McDaniel JR. Acute ethanol administration and acute allopregnanolone administration impair spatial memory in the Morris water task. Alc Clin Exp Res. 2002;26:1747–1751. doi: 10.1097/01.ALC.0000037219.79257.17. [DOI] [PubMed] [Google Scholar]
- Matthews KA, Woodall KL, Stoney CM. Changes in and stability of cardiovascular responses to behavioral stress: results from a four-year longitudinal study of children. Child Dev. 1990;61:1134–1144. [PubMed] [Google Scholar]
- McCarthy MM. GABA receptors make teens resistant to input. Nat Neurosci. 2007;10:397–399. doi: 10.1038/nn0407-397. [DOI] [PubMed] [Google Scholar]
- McCartney CR, et al. Obesity and sex steroid changes across puberty: Evidence for marked hyperandrogenemia in pre- and early pubertal obese girls. J Clin Endocrinol Metab. 2007;92:430–436. doi: 10.1210/jc.2006-2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McEwen BS. Physiology and neurobiology of stress and adaptation: central role of the brain. Physiol Rev. 2007;87:873–904. doi: 10.1152/physrev.00041.2006. [DOI] [PubMed] [Google Scholar]
- McEwen BS, Eiland L, Hunter RG, Miller MM. Stress and anxiety: structural plasticity and epigenetic regulation as a consequence of stress. Neuropharmacology. 2012;62:3–12. doi: 10.1016/j.neuropharm.2011.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGivern RF, Andersen J, Byrd D, Mutter KL, Reilly J. Cognitive efficiency on a match to sample task decreases at the onset of puberty in children. Brain Cogn. 2002;50:73–89. doi: 10.1016/s0278-2626(02)00012-x. [DOI] [PubMed] [Google Scholar]
- Meera P, Wallner M, Otis T. Molecular basis for the high THIP/gaboxadol sensitivity of extrasynaptic GABA-A receptors. J Neurophysiology. 2011;106:2057–2011. doi: 10.1152/jn.00450.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Megias M, Emri ZS, Freund mTF, Gulyas AI. Total number and distribution of inhibitory and excitatory synapses on hippocampal CA1 pyramidal cells. Neurosci. 2001;102:527–540. doi: 10.1016/s0306-4522(00)00496-6. [DOI] [PubMed] [Google Scholar]
- Mellon SH, Vaudry H. Biosynthesis of neurosteroids and regulation of their synthesis. Int Rev Neurobiol. 2001;46:33–78. doi: 10.1016/s0074-7742(01)46058-2. [DOI] [PubMed] [Google Scholar]
- Meredith RM, Floyer-Lea AM, Paulsen O. Maturation of long-term potentiation induction rules in rodent hippocampus: role of GABAergic inhibition. J Neurosci. 2003;23:11142–11146. doi: 10.1523/JNEUROSCI.23-35-11142.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller WL. Androgen synthesis in adrenarche. Rev Endocr Metab Disord. 2009;10:3–17. doi: 10.1007/s11154-008-9102-4. [DOI] [PubMed] [Google Scholar]
- Mitterling KL, Spencer JL, Dziedzic N, Shenoy S, McCarthy K, Waters EM, McEwen BS, Milner TA. Cellular and subcellular localization of estrogen and progestin receptor immunoreactivities in the mouse hippocampus. J Comp Neurol. 2010;518:2729–2743. doi: 10.1002/cne.22361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Modesti PA, Pela I, Cecioni I, Gensini GF, Serneri GG, Bartolozzi G. Changes in blood pressure reactivity and 24-hour blood pressure profile occurring at puberty. Angiology. 1994;45:443–450. doi: 10.1177/000331979404500605. [DOI] [PubMed] [Google Scholar]
- Mukai H, Higashi T, Nagura Y, Shimada K. Studies on neurosteroids XXV. Influence of 5alpha-reductase inhibitor, finasteride, on rat brain neurosteroid levels and metabolism. Biol Pharm Bull. 2008;31:1646–1650. doi: 10.1248/bpb.31.1646. [DOI] [PubMed] [Google Scholar]
- Nagashima K, Zorumski CF, Izumi Y. Propofol inhibits long-term potentiation but not long-term depression in rat hippocampal slices. Anesthesiology. 2005;103:318–326. doi: 10.1097/00000542-200508000-00015. [DOI] [PubMed] [Google Scholar]
- Neufang S, Specht K, Hausmann M, Gunturkun O, Herpertz-Dahlmann B, Fink GR, Konrad K. Sex differences and the impact of steroid hormones on the developing human brain. Cereb Cortex. 2009;19:464–473. doi: 10.1093/cercor/bhn100. [DOI] [PubMed] [Google Scholar]
- Newhouse P, Newhouse C, Astur RS. Sex differences in visual-spatial learning using a virtual water maze in pre-pubertal children. Behav Brain Res. 2007;183:1–7. doi: 10.1016/j.bbr.2007.05.011. [DOI] [PubMed] [Google Scholar]
- Newman AJ, Bavelier D, Corina D, Jezzard P, Neville HJ. A critical period for right hemispheric recruitment in American Sign Language processing. Nature Neuroscience. 2001;5:76–80. doi: 10.1038/nn775. [DOI] [PubMed] [Google Scholar]
- Nillni YI, Toufexis DJ, Rohan KJ. Anxiety sensitivity, the menstrual cycle, and panic disorder: a putative neuroendocrine and psychological interaction. Clin Psychol Rev. 2011;31:1183–1191. doi: 10.1016/j.cpr.2011.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nunez JL, McCarthy MM. Evidence for an extended duration of a GABA-mediated excitation in the developing male versus female hippocampus. Dev Neurobiol. 2007;67:1879–1890. doi: 10.1002/dneu.20567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olsen RW, Sieghart W. GABA A receptors: subtypes provide diversity of function and pharmacology. Neuropharmacology. 2009;56:141–148. doi: 10.1016/j.neuropharm.2008.07.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olsen RW, Snowman A. Chloride-dependent enhancment by barbiturates of gamma-aminobutyric acid receptor binding. J Neuroscience. 1982;2:1812–1823. doi: 10.1523/JNEUROSCI.02-12-01812.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ordaz S, Luna B. Sex differences in physiological reactivity to acute psychosocial stress in adolescence. Psychoneuroendocrinology. 2012;37:1135–1157. doi: 10.1016/j.psyneuen.2012.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pastalkova E, Serrano P, Pinkhasova D, Wallace E, Fenton AA, Sacktor TC. Storage of spatial information by the maintenance mechanism of LTP. Science. 2006;313:1141–1144. doi: 10.1126/science.1128657. [DOI] [PubMed] [Google Scholar]
- Patchev VK, Shoaib M, Holsboer F, Almeida OF. The neurosteroid tetrahydroprogesterone counteracts corticotropin-releasing hormone-induced anxiety and alters the release and gene expression of corticotropin-releasing hormone in the rat hypothalamus. Neuroscience. 1994;62:265–271. doi: 10.1016/0306-4522(94)90330-1. [DOI] [PubMed] [Google Scholar]
- Pepin M, Dorval M. Effect of playing a video game on a measure of spatial visualization. Percept Mot Skills. 1986a;62:159–162. doi: 10.2466/pms.1986.62.1.159. [DOI] [PubMed] [Google Scholar]
- Pepin M, Dorval M. Effect of playing a video game on adults' and adolescents' spatial visualization. Educational Resources Information Center. 1986b:2–18. ED273266. [Google Scholar]
- Pirker S, Schwarzer C, Wieselthaler A, Sieghart W, Sperk G. GABA(A) receptors: immunocytochemical distribution of 13 subunits in the adult rat brain. Neuroscience. 2000;101:815–850. doi: 10.1016/s0306-4522(00)00442-5. [DOI] [PubMed] [Google Scholar]
- Powers PS, Bruty H. Pharmacotherapy for eating disorders and obesity. Child Adolesc Psychiatr Clin N Am. 2009;18:175–187. doi: 10.1016/j.chc.2008.07.009. [DOI] [PubMed] [Google Scholar]
- Purdy RH, Morrow AL, Moore PH, Jr, Paul SM. Stress-induced elevations of gamma-aminobutyric acid type A receptor-active steroids in the rat. PNAS. 1991;88:4553–4557. doi: 10.1073/pnas.88.10.4553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rapkin AJ, Winer SA. Premenstrual syndrome and premenstrual dysphoric disorder: quality of life and burden of illness. Expert Rev Pharmacoecon Outcomes Res. 2009;9:157–170. doi: 10.1586/erp.09.14. [DOI] [PubMed] [Google Scholar]
- Reardon LE, Leen-Feldner EW, Hayward C. A critical review of the empirical literature on the relation between anxiety and puberty. Clin Psychol Rev. 2009;29:1–23. doi: 10.1016/j.cpr.2008.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy DS, Kaur G, Kulkarni SK. Sigma (sigma1) receptor mediated anti-depressant-like effects of neurosteroids in the Porsolt forced swim test. Neuroreport. 1998;9:3069–3073. doi: 10.1097/00001756-199809140-00028. [DOI] [PubMed] [Google Scholar]
- Rivera C, Voipio J, Payne JA, Ruusuvuori E, Lahtinen H, Lamsa K, Saarma M, Kaila K. The K-Cl cotransporter KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature. 1999;397:251–255. doi: 10.1038/16697. [DOI] [PubMed] [Google Scholar]
- Roberts DS, Hu Y, Lund IV, Brooks-Kayal AR, Russek SJ. Brain-derived neurotrophic factor (BDNF)-induced synthesis of early growth response factor 3 (Egr3) controls the levels of type A GABA receptor alpha4 subunits in hippocampal neurons. J Biol Chem. 2006;281:29431–29435. doi: 10.1074/jbc.C600167200. [DOI] [PubMed] [Google Scholar]
- Rudolph U, Crestani F, Benke D, Brünig I, Benson JA, Fritschy J-M, Martin JR, Bluethmann H, Möhler H. Benzodiazepine actions mediated by specific g-aminobutyric acidA receptor subtypes. Nature. 1999;401:796–800. doi: 10.1038/44579. [DOI] [PubMed] [Google Scholar]
- Safranski TJ, Lamberson WR, Keisler DH. Correlations among three measures of puberty in mice and relationships with estradiol concentration and ovluation. Biology of Reproduction. 1993;48:669–673. doi: 10.1095/biolreprod48.3.669. [DOI] [PubMed] [Google Scholar]
- Sanna E, Mostallino MC, Murru L, Carta M, Talani G, Zucca S, Mura ML, Maciocco E, Biggio G. Changes in expression and function of extrasynaptic GABA-A receptors in the rat hippocampus during pregnancy and after delivery. J Neurosci. 2009;29:1755–1765. doi: 10.1523/JNEUROSCI.3684-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarkar J, Wakefield S, MacKenzie G, Moss SJ, Maguire J. Neurosteroidogenesis is required for the physiological response to stress: role of neurosteroid-sensitive GABAA receptors. J Neurosci. 2011;31:18198–18210. doi: 10.1523/JNEUROSCI.2560-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato K, Akaishi T, Matsuki N, Ohno Y, Nakazawa K. beta-Estradiol induces synaptogenesis in the hippocampus by enhancing brain-derived neurotrophic factor release from dentate gyrus granule cells. Brain Res. 2007;1150:108–120. doi: 10.1016/j.brainres.2007.02.093. [DOI] [PubMed] [Google Scholar]
- Scharfman HE, Mercurio TC, Goodman JH, Wilson MA, MacLusky NJ. Hippocampal excitability increases during the estrous cycle in the rat: a potential role for brain-derived neurotrophic factor. J Neurosci. 2003;23:11641–11652. doi: 10.1523/JNEUROSCI.23-37-11641.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlaug G, Jancke L, Huang Y, Staiger JF, Steinmetz H. Increased corpus callosum size in musicians. Neuropsychologia. 1995;33:1047–1055. doi: 10.1016/0028-3932(95)00045-5. [DOI] [PubMed] [Google Scholar]
- Schmidt P, Nieman L, Danaceau M, Adams L, Rubinow D. Differential behavioral effects of gonadal steroids in women with premenstrual syndrome. New England J Med. 1998;338:209–216. doi: 10.1056/NEJM199801223380401. [DOI] [PubMed] [Google Scholar]
- Sharma A, Gilley PM, Dorman MF, Baldwin R. Deprivation-induced cortical reorganization in children with cochlear implants. Int J Audiol. 2007;46:494–499. doi: 10.1080/14992020701524836. [DOI] [PubMed] [Google Scholar]
- Shavalier M. The effects of CAD-like software on the spatial ability of middle school students. J Educat Comput Res. 2004;31:37–49. [Google Scholar]
- Shen H, Gong QH, Aoki C, Yuan M, Ruderman Y, Dattilo M, Williams K, Smith SS. Reversal of neurosteroid effects at alpha4-beta2-delta GABA-A receptors triggers anxiety at puberty. Nat Neurosci. 2007;10:469–477. doi: 10.1038/nn1868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen H, Gong QH, Yuan M, Smith SS. Short-term steroid treatment increases delta GABA-A receptor subunit expression in rat CA1 hippocampus: pharmacological and behavioral effects. Neuropharmacology. 2005;49:573–586. doi: 10.1016/j.neuropharm.2005.04.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen H, Sabaliauskas N, Sherpa A, Fenton AA, Stelzer A, Aoki C, Smith SS. A critical role for a4bd GABA-A receptors in shaping learning deficits at puberty in mice. Science. 2010a;327:1515–1518. doi: 10.1126/science.1184245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen H, Sabaliauskas N, Sherpa A, Fenton AA, Stelzer A, Aoki C, Smith SS. A critical role for alpha4-beta-delta GABA-A receptors in shaping learning deficits at puberty in mice. Science (Supp Online Material) 2010b;327:1515–1518. doi: 10.1126/science.1184245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sigel E. Mapping of the benzodiazepine recognition site on GABA(A) receptors. Curr Top Med Chem. 2002;2:833–839. doi: 10.2174/1568026023393444. [DOI] [PubMed] [Google Scholar]
- Smith SS, Ruderman Y, Frye CA, Homanics GE, Yuan M. Steroid withdrawal in the mouse results in anxiogenic effects of 3alpha,5alpha-THP: A possible model of premenstrual dysphoric disorder. Psychopharmacology. 2006;186:323–333. doi: 10.1007/s00213-005-0168-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith SS, Shen H, Gong QH, Zhou X. Neurosteroid regulation of GABA(A) receptors: Focus on the alpha4 and delta subunits. Pharm and Therapeutics. 2007;116:58–76. doi: 10.1016/j.pharmthera.2007.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith SS, Waterhouse BD, Woodward DJ. Locally applied progesterone metabolites alter neuronal responsiveness in the cerebellum. Brain Research Bulletin. 1987a;18:739–747. doi: 10.1016/0361-9230(87)90209-7. [DOI] [PubMed] [Google Scholar]
- Smith SS, Waterhouse BD, Woodward DJ. Progesterone alters GABA and glutamate responsiveness: A possible mechanism for its anxiolytic action. Brain Res. 1987b;400:353–359. doi: 10.1016/0006-8993(87)90634-2. [DOI] [PubMed] [Google Scholar]
- Staley KJ, Mody I. Shunting of excitatory input to dentate gyrus granule cells by a depolarizing GABAA receptor-mediated postsynaptic conductance. J Neurophysiol. 1992;68:197–212. doi: 10.1152/jn.1992.68.1.197. [DOI] [PubMed] [Google Scholar]
- Staley KJ, Proctor WR. Modulation of mammalian dendritic GABA(A) receptor function by the kinetics of Cl− and HCO3− transport. J Physiol. 1999;519:693–712. doi: 10.1111/j.1469-7793.1999.0693n.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stell BM, Brickley SG, Tang CY, Farrant M, Mody I. Neuroactive steroids reduce neuronal excitability by selectively enhancing tonic inhibition mediated by delta subunit-containing GABA-A receptors. Proc Natl Acad Sci. 2003;100:14439–14444. doi: 10.1073/pnas.2435457100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stell BM, Mody I. Receptors with different affinities mediate phasic and tonic GABA(A) conductances in hippocampal neurons. J Neurosci. 2002;22:RC223. doi: 10.1523/JNEUROSCI.22-10-j0003.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sterlemann V, Rammes G, Wolf M, Liebl C, Ganea K, Muller MB, Schmidt MV. Chronic social stress during adolescence induces cognitive impairment in aged mice. Hippocampus. 2010;20:540–549. doi: 10.1002/hipo.20655. [DOI] [PubMed] [Google Scholar]
- Subrahmanyam K, Greenfield P. Effect of video game practice on spatial skills in girls and boys. J Applied Devel Psychol. 1994;15:15–32. [Google Scholar]
- Sumter SR, Bokhorst CL, Miers AC, Van PJ, Westenberg PM. Age and puberty differences in stress responses during a public speaking task: do adolescents grow more sensitive to social evaluation? Psychoneuroendocrinology. 2010;35:1510–1516. doi: 10.1016/j.psyneuen.2010.05.004. [DOI] [PubMed] [Google Scholar]
- Sundstrom I, Nyberg S, Backstrom T. Patients with premenstrual syndrome have reduced snsitivity to midazolam compared to control subjects. Neuropsychopharm. 1997;17:370–381. doi: 10.1016/S0893-133X(97)00086-9. [DOI] [PubMed] [Google Scholar]
- Sundstrom-Poromaa I, Smith DH, Gong Q, Sabado TN, Li X, Light A, Wiedmann M, Williams K, Smith S. Hormonally regulated a4b2d GABA-A receptors are a target for alcohol. Nat Neurosci. 2002;5:721–722. doi: 10.1038/nn888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Susman EJ, Nottelmann ED, Dorn LD, Inoff-Germain G, Chrousos GP. Physiological and behavioral aspects of stress in adolescence. Adv Exp Med Biol. 1988;245:341–352. doi: 10.1007/978-1-4899-2064-5_27. [DOI] [PubMed] [Google Scholar]
- Swinbourne J, Hunt C, Abbott M, Russell J, St CT, Touyz S. The comorbidity between eating disorders and anxiety disorders: prevalence in an eating disorder sample and anxiety disorder sample. Aust N Z J Psychiatry. 2012;46:118–131. doi: 10.1177/0004867411432071. [DOI] [PubMed] [Google Scholar]
- Syme C, Abrahamowicz M, Leonard GT, Perron M, Richer L, Veillette S, Xiao Y, Gaudet D, Paus T, Pausova Z. Sex differences in blood pressure and its relationship to body composition and metabolism in adolescence. Arch Pediatr Adolesc Med. 2009;163:818–825. doi: 10.1001/archpediatrics.2009.92. [DOI] [PubMed] [Google Scholar]
- Tena-Sempere M. Neuroendocrinology of puberty: recent milestones and new challenges. Mol Cell Endocrinol. 2010;324:1–2. doi: 10.1016/j.mce.2010.05.006. [DOI] [PubMed] [Google Scholar]
- Tinggaard J, Mieritz MG, Sorensen K, Mouritsen A, Hagen CP, Aksglaede L, Wohlfahrt-Veje C, Juul A. The physiology and timing of male puberty. Curr Opin Endocrinol Diabetes Obes. 2012;19:197–203. doi: 10.1097/MED.0b013e3283535614. [DOI] [PubMed] [Google Scholar]
- Tokuda K, Izumi Y, Zorumski CF. Ethanol enhances neurosteroidogenesis in hippocampal pyramidal neurons by paradoxical NMDA receptor activation. J Neurosci. 2011;31:9905–9909. doi: 10.1523/JNEUROSCI.1660-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres JM, Ruiz E, Ortega E. Effects of CRH and ACTH administration on plasma and brain neurosteroid levels. Neurochem Res. 2001;26:555–558. doi: 10.1023/a:1010925331768. [DOI] [PubMed] [Google Scholar]
- Trincavelli ML, Da PE, Daniele S, Martini C. The GABAA-BZR complex as target for the development of anxiolytic drugs. Curr Top Med Chem. 2012;12:254–269. doi: 10.2174/1568026799078787. [DOI] [PubMed] [Google Scholar]
- Uysal N, Sisman AR, Dayi A, Ozbal S, Cetin F, Baykara B, Aksu I, Tas A, Cavus SA, Gonenc-Arda S, Buyuk E. Acute footshock-stress increases spatial learning-memory and correlates to increased hippocampal BDNF and VEGF and cell numbers in adolescent male and female rats. Neurosci Lett. 2012;514:141–146. doi: 10.1016/j.neulet.2012.02.049. [DOI] [PubMed] [Google Scholar]
- Veselis RA, Pryor KO, Reinsel RA, Li Y, Mehta M, Johnson R., Jr Propofol and midazolam inhibit conscious memory processes very soon after encoding: an event-related potential study of familiarity and recollection in volunteers. Anesthesiology. 2009;110:295–312. doi: 10.1097/ALN.0b013e3181942ef0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vetter-O'Hagen CS, Spear LP. Hormonal and physical markers of puberty and their relationship to adolescent-typical novelty-directed behavior. Dev Psychobiol. 2012;54:523–535. doi: 10.1002/dev.20610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vickers K, McNally RJ. Is premenstrual dysphoria a variant of panic disorder? A review. Clin Psychol Rev. 2004;24:933–956. doi: 10.1016/j.cpr.2004.08.001. [DOI] [PubMed] [Google Scholar]
- Voss P, Gougoux F, Zatorre RJ, Lassonde M, Lepore F. Differential occipital responses in early-and late-blind individuals during a sound-source discrimination task. Neuroimage. 2008;40:746–758. doi: 10.1016/j.neuroimage.2007.12.020. [DOI] [PubMed] [Google Scholar]
- Waddell J, Kim J, Alger BE, McCarthy MM. The depolarizing action of GABA in cultured hippocampal neurons is not due to the absence of ketone bodies. PLoS One. 2011;6:e23020. doi: 10.1371/journal.pone.0023020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walf AA, Frye CA. Conjugated equine estrogen enhances rats' cognitive, anxiety, and social behavior. Neuroreport. 2008;19:789–792. doi: 10.1097/WNR.0b013e3282fe209c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallner M, Hanchar HJ, Olsen RW. Ethanol enhances alpha 4-beta 3-delta and alpha 6-beta 3-delta gamma-aminobutyric acid type A receptors at low concentrations known to affect humans. Proc Natl Acad Sci. 2003;100:15218–15223. doi: 10.1073/pnas.2435171100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe Y, Gould E, McEwen BS. Stress induces atrophy of apical dendrites of hippocampal CA3 pyramidal neurons. Brain Res. 1992;588:341–345. doi: 10.1016/0006-8993(92)91597-8. [DOI] [PubMed] [Google Scholar]
- Weber-Fox C, Neville HJ. Sensitive periods differentiate processing of open- and closed-class words: an ERP study of bilinguals. J Speech Lang Hear Res. 2001;44:1338–1353. doi: 10.1044/1092-4388(2001/104). [DOI] [PubMed] [Google Scholar]
- Wieland HA, Luddens H, Seeburg PH. A single histidine in GABA-A receptors is essential for benzodiazepine agonist binding. J Biol Chem. 1992;267:1426–1429. [PubMed] [Google Scholar]
- Wisden W, Laurie DJ, Monyer H, Seeburg P. The distribution of 13 GABA-A receptor subunit mRNAs in the rat brain. I. Telencephalon, diencephalon, mesencephalon. J Neurosci. 1992;12:1040–1062. doi: 10.1523/JNEUROSCI.12-03-01040.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wohlfarth KM, Bianchi MT, Macdonald RL. Enhanced neurosteroid potentiation of ternary GABA(A) receptors containing the delta subunit. J Neurosci. 2002;22:1541–1549. doi: 10.1523/JNEUROSCI.22-05-01541.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolk S, Bloom D. The interactive effects of locus of control and situational stress upon performance accuracy and time. J Pers. 1978;46:279–298. doi: 10.1111/j.1467-6494.1978.tb00180.x. [DOI] [PubMed] [Google Scholar]
- Wood GE, Shors TJ. Stress facilitates classical conditioning in males, but impairs classical conditioning in females through activational effects of ovarian hormones. Proc Natl Acad Sci U S A. 1998;95:4066–4071. doi: 10.1073/pnas.95.7.4066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright BA, Zecker SG. Learning problems, delayed development, and puberty. Proc Natl Acad Sci. 2004;101:9942–9946. doi: 10.1073/pnas.0401825101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu Y, Wang W, Richerson G. Vigabatrin induces tonic inhibition via GABA transporter reversal without increasing vesicular GABA release. J Neurophysiol. 2003;89:2021–2034. doi: 10.1152/jn.00856.2002. [DOI] [PubMed] [Google Scholar]
- Xu JY, Sastry BR. Theta-bursts induce a shift in reversal potentials for GABA-A receptor-mediated postsynaptic currents in rat hippocampal CA1 neurons. Exp Neurol. 2007;204:836–839. doi: 10.1016/j.expneurol.2007.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yerkes RM, Dodson JD. The relation of strength of stimulus to rapidity of habit-formation. J Comp Neurol and Psych. 1908;18:459–482. [Google Scholar]
- Yonkers KA, O'Brien PM, Eriksson E. Premenstrual syndrome. Lancet. 2008;371:1200–1210. doi: 10.1016/S0140-6736(08)60527-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zgourides GD, Warren R. Prevalence of panic in adolescents: a brief report. Psychol Rep. 1988;62:935–937. doi: 10.2466/pr0.1988.62.3.935. [DOI] [PubMed] [Google Scholar]
- Zheleznova N, Sedelnikova A, Weiss DS. Alpha1-beta2-delta, a silent GABA-A receptor: recruitment by tracazolate and neurosteroids. Brit J Pharmacol. 2008;153:1062–1071. doi: 10.1038/sj.bjp.0707665. [DOI] [PMC free article] [PubMed] [Google Scholar]