

Abstract
The present study was undertaken to investigate the protective effect of the filamentous cyanobacterium Spirulina platensis (S. platensis) on mercury (II) chloride (HgCl2)-induced oxidative damages and histopathological alterations in the testis of Wistar albino rats. The animals were divided into four equal groups, i ) control, ii ) HgCl2, iii ) S. platensis and iv ) combination of HgCl2+S. platensis. Oxidative stress, induced by a single dose of HgCl2 (5 mg/kg, bw; subcutaneously, s.c.), substantially decreased (P<0.01) the activity level of testicular key enzymatic antioxidant biomarkers (superoxide dismutase, SOD; catalase, CAT and glutathione peroxidase, GPx), oxidative stress makers (blood hydroperoxide; testicular reduced glutathione, GSH and malondialdehyde, MDA), and testicular mercury levels. Moreover, HgCl2 administration resulted in a significant (P<0.01) increase in the number of sperms with abnormal morphology and decrease in epididymal sperm count, motility, plasma testosterone level and testicular cholesterol. Furthermore, HgCl2 exposure induced histopathological changes to the testis including morphological alterations of the seminiferous tubules, and degeneration and dissociation of spermatogenic cells. Notably, oral pretreatment of animals with Spirulina (300 mg/kg, bw) lowered the extent of the observed HgCl2-mediated toxicity, whereby significantly reducing the resulting lipid peroxidation products, mercury accumulation in the testis, histopathological changes of the testes and spermatozoal abnormalities. In parallel, the pretreatment with Spirulina also completely reverted the observed Hg-Cl2-induced inhibition in enzymatic activities of antioxidant biomarkers (SOD, CAT and GPx) back to control levels. The pretreatment of rats with S. platensis significantly recovered the observed HgCl2-mediated decrease in the weight of accessory sex organs. Taken together, our findings clearly highlight the role of S. platensis as a protective modulator of HgCl2-induced testicular injuries and suggest some therapeutic potential in mammals. Further investigation of therapeutic strategies employing Spirulina against heavy metals toxicity in humans is therefore warranted.
Introduction
Heavy metals have become one of many contaminants found in our environment. Many of these metals, including lead, mercury, cobalt, cadmium, and chromium are known to exert toxic effects on testicular function, while others such as zinc, manganese, and selenium have been shown to be essential for normal functioning of the testis [1], [2], [3]. Mercury is a ubiquitous element in the environment causing oxidative stress in the exposed individuals leading to tissue damage. Its contamination and toxicity has posed a serious hazard to human health. The release of mercury from dental amalgam dominates exposure to inorganic mercury and may have an acceptable risk among the general population [4]. Human exposure to mercury can occur by inhalation, ingestion and consumption via food chain. Adverse effects of metals on human reproduction and development continue to be a demanding challenge for researchers. Mercury compounds are known to affect testicular spermatogenic and steroidogenic functions in experimental animals [5]. Oral exposure of mercuric chloride produced adverse effects on the reproductive performance of mice [6]. Mercury affects accessory sex glands function in rats and mice through androgen deficiency [7], [8]. Decrements in sperm count, motility and morphology have been reported in methyl mercury exposed monkeys and rodents [8], [9]. The activity of superoxide dismutase, glutathione peroxidase and glutathione reductase of sperm incubated in mercury decreased while thiobarbituric reactive substances (TBARS) levels and H2O2 generation were increased [10]. The treatment of rats with Hg led to a decrease in GSH levels in different tissues [11]. Hg induced oxidative stress in the testicular tissue of the rat as manifested by a decrease in SOD and catalase and an increase of malondialdehyde (MDA) levels [12].
Mercuric chloride is one of the most toxic forms of mercury because it easily forms organomercury complexes with proteins [13]. The inorganic ionic mercury has great affinity for SH groups of biomolecules, such as glutathione (GSH) and sulfhydryl proteins, which may contribute to its toxicity [14]. Once bound to GSH, Hg can leave the cell to circulate in serum or lymph and be deposited in other organs or tissues [13]. Mercuric chloride is considered to be one of the pro-oxidants that induce oxidative stress [6]. Oxidative stress occurs when the production of reactive oxygen species (ROS) such as, superoxide anion (•O− 2), hydrogen peroxide (H2O2) and the hydroxyl radical (•OH) exceeds the body's defense mechanism, causing damage to macromolecules such as DNA, proteins and lipids [15] and trigger many pathological processes in the male reproductive system [16]. There is evidence that ROS may have a detrimental effect on critical components of the steroidogenic pathway [17]. Excessive production of ROS above normal levels results in lipid peroxidation and membrane damage leading to loss of sperm motility [7], inactivation of glycolytic enzymes and damage to the acrosomal membranes [18] which render the sperm cell unable to fertilize the oocyte, or produce a viable pregnancy [19].
Spirulina platensis, recently renamed Arthrospira, is a filamentous cyanobacterium (blue-green alga) that belongs to the Oscillatoraceae family and has a long history for use as food. It is rich in proteins, lipids, carbohydrates and some vital elements like zinc, magnesium, manganese, selenium, ß-carotene, riboflavin, α-tocopherol and α-linoleic acid [20]. The antioxidant properties of spirulina and its capacity to scavenge hydroxyl radicals [21], and to inhibit lipid peroxidation [22] have attracted the attention of many researches. Spirulina species exhibited various biological activities such as antihypertensive and antihyperlipidemic [23]. Moreover, Spirullina fusiformis provides protection against mercuric chloride induced oxidative stress and alteration of antioxidant defense mechanism in the liver [24]. These activities were largely related to phycocyanin, an active protein of Spirullina [25]. It was reported that feeding of mice a diet supplemented with 30% of Spirulina maxima for 13 weeks did not produce any toxic effects [26]. Moreover, Spirulina maxima treatment was not associated with any adverse effects on reproductive performance, including male and female fertility and duration of gestation in rats [27]. Furthermore, no significant alteration was observed in the motility and shape of sperm of rats treated with Spirulina maxima (30%) incorporated into the diet for 5 days [28]. Oral administration of Spirullina fusiformis to mice can modulate mercury induced alteration in testicular acid and alkaline phosphatase activities [29]. To our knowledge, there is no available report until now describing the protective properties of S. platensis against mercury-induced injuries to male reproductive system.
In the present study, an attempt has been made to evaluate the protective potential of S. platensis against mercury-induced oxidative damage to testes of male rats by studying testicular antioxidant defense system, sperm quality and plasma testosterone level.
Materials and Methods
Animals
Male Wistar albino rats (Rattus norvegicus), weighing 180–200 g, were obtained from the animal house, Faculty of Pharmacy, King Saud University. The animals were housed throughout the experiment in polypropylene cages (each cage housing eight animals) and allowed to acclimatize to laboratory environment for seven days before the beginning of the experiment. Animals were maintained under controlled conditions of temperature (23±1°c), humidity (50±15%) and normal photoperiod (12–12 h light-dark cycles). Rats were allowed free access to standard dry pellet diet and water ad libitum. This study was conducted in the Zoology Department, Faculty of Science, King Saud University, Saudi Arabia. The care and handling of experimental animals were carried out according to the animal ethical committee of the College of Pharmacy, King Saud University.
Test chemicals
Mercury in the form of HgCl2 was purchased from Merck (Darmstadt, Germany) (Product no. 104417), while S. platensis was obtained from Alibaba Company (China) in the form of tablets. 5–5-dithio-bis(2-nitrobenzoic acid) (DTNB), thiobarbituric acid and reduced glutathione were purchased from Sigma-Aldrich Corp. (St. Louis, MO USA).
Treatments
The animals were divided into four groups, each containing eight rats. Animals in group I were used as control group and no treatment was given to these rats. Animals in group II were given S. platensis daily by gavage for 60 consecutive days at dose level of 300 mg/kg b.wt [30] suspended in water. Animals in group III were administered a single dose (5 mg/kg bw) of mercury (II) chloride (HgCl2) by subcutaneous (s.c.) route with the help of Hamilton syringe [31], dissolved in water three-time a week for 60 days. The s.c. route was chosen to ensure the exact and the proper uptake of HgCl2. The administered dose of 5 mg/kg bw (i.e., 15 mg/kg bw/wk) has been chosen based on previous studies which employed HgCl2 doses of 5, 10 and 20 mg/kg bw/day. These doses were optimal for the induction of testicular histopathological changes testes without observed toxicity [32] Animals in group IV were given S. platensis (300 mg/kg bw) by gavage for 10 consecutive days before mercuric chloride administration and continued up to 60 days along with HgCl2 treatment.
Collection of samples
At the end of the experimental period, blood was drawn from the animals by puncturing retro-orbital venous sinus. Whole blood was used for the determination of hydroperoxide level, while separated plasma was used to determine testosterone level. After collection of blood samples, the animals from all groups were autopsied under light ether anesthesia. Subsequently, testes, vas deferens, epididymis, prostate glands and seminal vesicles were excised from surrounding tissues and placed into tube. Thus, organs were dried between two sheets of filter paper and their wet weight was determined. Next, the relative organ weight was calculated by use of the formula: organ weight/body weight ×100. Left epididymis was weighed, while right epididymis was rinsed in warm phosphate buffered saline (PBS) and incubated at 37°C for the evaluation of sperm quality. Whereas, left testes were processed for histological study, right testes were processed for determination of MDA, GSH, SOD, CAT, GPx and cholesterol.
Biochemical studies
Determination of lipid peroxide levels
Blood hydroperoxide level was evaluated using an analytical system (Iram, Parma, Italy). The test is a colorimetric test that takes advantage of the ability of hydroperoxide to generate free radicals after reacting with transitional metals, when buffered chromogenic substance is added; a colored complex appears. This complex was measured spectrophotometrically. Lipid peroxidation level in the testis was measured by a method [33] using thiobarbituric acid reactive substances (TBARS) with some modifications as previously described [34]. Testis was homogenized in ice cold 0.15 M KCl (10%) and the concentration of TBARS was expressed as nmol of MDA per mg protein using 1,1,3,3-tetramethoxypropane as standard. The absorbance was read at 532 nm.
Determination of testicular mercury
Total mercury concentration was determined in testes using a previously reported protocol [35]. The samples were heated with 7 ml nitric acid and perchloric acid (2v/1v) in boiling water bath until evaporation. After digestion, the samples were diluted by definite volume of distilled water and filtered. The mercury concentration was measured in samples by use of atomic absorption, Model RA-2 (Tokyo, Japan). Data are presented as total mercury (µg/g of tissue dry).
Determination of testicular reduced glutathione (GSH)
Reduced form of glutathione was determined using Ellman's reagent 5-5-dithio-bis (2-nitrobenzoic acid) (DTNB) as a coloring reagent [36]. The absorbance was read at 412 nm by spectrophotometer. GSH concentration was calculated from a standard curve.
Determination of testicular superoxide dismutase (SOD)
Testicular superoxide dismutase was assayed by the method of Asada [37], which involves the inhibition of photochemical reduction of nitro blue tetrazolium (NBT) at pH 8.0. A single unit of enzyme is defined as the quantity of superoxide dismutase required to produce 50% inhibition of photochemical reduction of NBT. The absorbance was read at 580 nm against a blank using UV – Vis spectrophotometer. The activity was expressed as U/mg protein.
Determination of testicular catalase (CAT)
Catalase activity was estimated in testis homogenate by the method reported by Aebi [38]. The specific activity of catalase has been expressed as µmoles of H2O2 consumed/min/mg protein. The difference in absorbance at 240 nm per unit time is a measure of catalase activity.
Determination of testicular glutathione peroxidase (GPX)
The activity of the antioxidant enzyme glutathione peroxidase was determined using glutathione reductase and NADPH. This method is based on the oxidation of NADPH at 25°C, which is indicated by the decrease in absorbance at 340 nm [39]. Results are expressed in U/mg protein.
Determination of testicular cholesterol
The estimation of testicular cholesterol was carried out by the method of Zlatki [40]. To test tubes containing 5 ml of working FeCl3 solution, 0.2 ml of testis homogenate prepared in glacial acetic acid was added. The contents were mixed and 3 ml of concentrated H2SO4. The optical density after color development was read at 540 nm on a spectrophotometer and expressed as mg/100 mg tissue wt.
Hormonal assay
Plasma testosterone concentration was measured by enzyme immunoassay using commercial kit from Diagnostic products Co. (Los Angeles, CA USA).
Sperm analysis
Collection and incubation of epididymal sperm
Spermatozoa were obtained from the fresh right epididymis of adult rat described by Narayana [41]. Briefly, epididymis was cut into small pieces with a sharp razor blade and dispersed in 3 ml of phosphate buffered saline (pH 7.2) to obtain a suspension with gentle stirring. Dispersed sperm samples were kept in an incubator.
Sperm motility
Ten µl of the suspension was placed in a warm slide and the percentage of motile sperm was counted under Nikon binocular microscope with warmed stage and about 300 spermatozoa were evaluated. Motility was then expressed as the percentage of motile spermatozoa.
Sperm count
The suspension was filtered. An aliquot from the suspension (up to 0.5 ml) was taken in leukocyte micropipette of hemocytometer and diluted with phosphate buffered solution up to the mark 11. The suspension was well-mixed and charged into Neubauer's counting chamber. The total sperm count in 8 big squares was determined and multiplied by 3×5×104 [42] to express the number of spermatozoa/epididymis (million/epididymis).
Sperm abnormalities
For the evaluation of the sperm morphology the filtrate (obtained as described above in Section 2.7.3) was stained with 1% eosin and morphological defects were analyzed [42]. Briefly, the spermatozoa in the smears were visualized under light microscope (400×) and any abnormalities of either heads or tails were noted. Three hundred sperms were screened for each animal and total abnormality was expressed as percentage of incidence/300 sperm/animal.
Histological determination
For microscopic evaluation, testis tissues were fixed in 10% formol saline, embedded in paraffin, sectioned at 5 µm and stained with hematoxylin/eosin. Sections were studied under light microscope at 400× magnification.
Statistical analysis
All values from duplicate measures for each sample (n = 8) were expressed as means with ± S.E. Statistical analysis of data was performed using two-way ANOVA followed by Fisher's least significant difference (LSD) procedure for comparison of various treatments using the IBM SPSS software (version 20.0, SPSS Inc., Chicago, IL). Difference in statistics at P<0.05 were considered significant.
Results
Lipid peroxidation products and testicular mercury concentration
Results presented in Table 1 indicate that there was a statistically significant decrease (P<0.01) in the level of blood hydroperoxide in Spirulina-treated animals (Group II), while the reduction in MDA level was not significant. Mercuric chloride (HgCl2) administration resulted in a significant (P<0.01) elevation in the levels of blood hydroperoxide and testicular MDA by 2.15 and 1.36 folds, respectively, as compared to values in the control group I. The blood hydroperoxide level also increased significantly (P<0.01) in the combined treatment of S. platensis with HgCl2 by 1.60 fold, while testicular MDA did not change significantly compared to the control values. Moreover, the hydroperoxide or MDA level of S. platensis+HgCl2 group was significantly (P<0.01) lower than that of exposed to HgCl2 alone. Although, mercury level in testis exhibited a significant elevation (P<0.01) in both HgCl2 and S. platensis+HgCl2 groups; its level in the later group was significantly less than that of the HgCl2 group.
Table 1. Effect of S. platensis on lipid peroxidation products and testicular mercury concentration in experimental HgCl2-exposed rats as compared to control group.
| Groups | ||||
| Spirulina+HgCl2 | HgCl2 | Spirulina | Control | Parameters |
| 45.96±2.89**b | 61.23±5.15** | 24.11±0.32** | 28.42±0.63 | Blood hydroperoxide level (mg/100 ml) |
| 5.20±0.05b | 6.96±0.05** | 4.81±0.13 | 5.10±0.06 | Testicular MDA (nmol/mg protein) |
| 1.40±0.02**b | 2.15±0.08** | 00.00 | 00.00 | Mercury concentration (µg/g dry tissue) |
The relative organ weight was calculated by use of the formula: organ weight/body weight ×100. Each value represents the mean ± S.E., n = 8. Values marked with an asterisk (*) differ significantly from control value (** P <0.01), while those marked with letter differ significantly from HgCl2 group (P<0.01).
Testicular reduced glutathione and enzymatic antioxidant biomarkers
A significant (P<0.01) increase in reduced glutathione (GSH) and SOD (P<0.05) was observed in Spirulina group (Table 2). Although HgCl2 exposure alone resulted in a significant (P<0.01) inhibition in the level of testicular GSH and activity levels of testicular antioxidant enzymes (SOD, CAT and GPx,) as compared to the control group, S. platensis administration to HgCl2-exposed rats prevented the observed decrease of these parameters.
Table 2. Effect of S. platensis on testicular glutathione content and activities of antioxidant biomarker enzymes in experimental HgCl2-exposed rats as compared to control group.
| Groups | ||||
| Spirulina+HgCl2 | HgCl2 | Spirulina | Control | Parameters |
| 338.23±2.50b | 231.92±1.23** | 374.12±2.50** | 342.48±2.05 | GSH (nmol/mg protein) |
| 0.83±0.007b | 0.56±0.01** | 0.84±0.01 | 0.85±0.01 | GPX (µ/mg protein) |
| 5.59±0.11b | 3.63±0.09** | 6.12±0.10 | 5.90±0.08 | CAT (µ mol H2O2/min/mg protein) |
| 23.35±0.28b | 14.11±0.22** | 25.92±0.27* | 24.39±0.47 | SOD (µ/mg protein |
Each values represents the mean ± S.E., n = 8. Values marked with an asterisk(*) differ significantly from control value (* P<0.05,** P<0.01), while those marked with a letter (b) differ significantly from HgCl2 group (** P<0.01).
Testicular cholesterol and plasma testosterone
Mean level of testicular cholesterol significantly (P<0.01) decreased in both HgCl2-exposed and S. platensis+HgCl2 groups by 47% and 7%, respectively (Table 3). Moreover, the concentration of plasma testosterone was significantly (P<0.01) blunted in HgCl2 and Spirulina+HgCl2 groups by 41% and 17%, respectively. However, the mean values of the two parameters in S. platensis+HgCl2 group were significantly higher (P<0.01) than those of HgCl2 group (Table 3).
Table 3. Effect of S. platensis on testicular cholesterol and plasma testosterone concentrations in experimental HgCl2-exposed rats as compared to control group.
| Groups | ||||
| Spirulina+HgCl2 | HgCl2 | Spirulina | Control | Parameters |
| 0.28±0.003**b | 0.16±0.002** | 0.31±0.01 | 0.30±0.004 | Testicular cholesterol (mg/100 g bw) |
| 2.10±0.15*b | 1.49±0.06** | 2.68±0.07 | 2.54±0.08 | Plasma testosterone (ng/ml) |
Each values represents the mean ± S.E., n = 8. Values marked with an asterisk (*) differ significantly from control value (* P<0.05,** P<0.01), while those marked with a letter differ (b) significantly from HgCl2 group (* P<0.01).
Weights of reproductive organ
The values of organ weights are shown in Table 4. The weights of testes and accessory sex organs decreased significantly (P<0.01) in both HgCl2-exposed and S. platensis+HgCl2 groups. However, the values of accessory sex organs of S. platensis+HgCl2 group were significantly higher than those of the HgCl2-exposed group.
Table 4. Effect of S. platensis on reproductive organ weights (g) relative to body weight in experimental HgCl2-exposed rats as compared to control group.
| Groups | ||||
| Spirulina+HgCl2 | HgCl2 | Spirulina | Control | Organs |
| 1.06±0.03** | 1.07±0.02** | 1.18±0.02 | 1.23±0.01 | Testis |
| 0.05±0.003**a | 0.04±0.003** | 0.06±0.005 | 0.07±0.003 | Vas deferens |
| 0.24±0.003b | 0.21±0.007** | 0.24±0.008 | 0.25±0.003 | Epididymis |
| 0.23±0.01**a | 0.19±0.01** | 0.29±0.01 | 0.31±0.02 | Prostate |
| 0.33±0.02**b | 0.26±0.01** | 0.50±0.04 | 0.52±0.01 | Seminal vesicle |
The relative organ weight was calculated by use of the formula: organ weight/body weight ×100. Each values is the mean ± S.E., n = 8. Values marked with an asterisk (*) differ significantly from control value; **P<0.01. Those marked with a letter differ significantly from HgCl2 group; Pa<0.05; Pb<0.01.
Epididymal sperm characteristics
The results of sperm analysis are presented in Table 5. Both sperm motility and concentration of S. platensis group increased significantly (P<0.01) as compared to control values. Although HgCl2 treatment caused a significant decrease (P<0.01) in sperm motility and concentration, it resulted in a significant (P<0.01) increase in head, tail and total sperm abnormalities when compared with the values of control group. The administration of S. platensis+HgCl2 in combination tended to increase sperm motility along with sperm concentration and to decrease abnormal sperm rates as compared to HgCl2 group.
Table 5. Effect of S. platensis on sperm morphological quality parameters in experimental HgCl2-exposed rats as compared to control group.
| Groups | ||||
| Spirulina+HgCl2 | HgCl2 | Spirulina | Control | Parameters |
| 79.32±1.02**b | 63.87±1.57** | 89.08±0.47** | 84.50±0.56 | Sperm motility (%) |
| 29.49±1.29b | 21.69±0.84** | 33.67±1.41** | 27.39±0.75 | Sperm count per epididymis (million/epididymis) |
| Abnormal sperm rate (%)+ | ||||
| 5.60±0.36**b | 8.25±0.25** | 2.42±0.22 | 2.11±0.13 | Head |
| 2.45±0.32a | 3.40±0.24** | 2.26±0.18 | 2.06±0.10 | Tail |
| 7.99±0.58**b | 11.66±0.62** | 4.68±0.27 | 4.17±0.19 | Total |
Normal sperms were characterized with hook-shaped head and straight tail in control animals. Morphological abnormalities observed in HgCl2-exposed spermatozoa were absence of head, head winding around tail, coiled tails and kinks in midpiece and tail regions, bent and curved tail. Each values is the mean ± S.E., n = 8. Values marked with an asterisk (*) differ significantly from control value: **P<0.01, those marked with letter differ significantly from HgCl2 group; Pa<0.05, Pb<0.01.
Histopathology
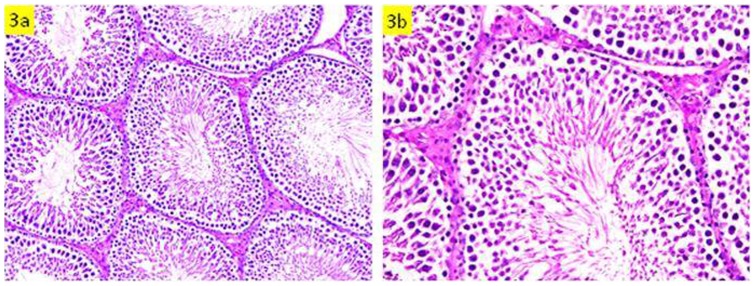
Histopathological examination revealed that rats administered with HgCl2 alone exhibited testicular lesions in some seminiferous tubules comprising decrease of luminal spermatozoa, irregular basement membrane, disorganization and degeneration of some spermatogenic cells and hemorrhage in interstitial tissues, as compared to the control group (Figure 1 & 2). Pre-administration of S. platensis protected the testicular tissue of rats exposed to HgCl2, as evidenced by the appearance of normal structure of testicular seminiferous tubules (Figure 3).
Figure 1. Testicular section of control rat which show normal spermatogenesis and cell arrangement in the seminiferous tubules (a: H&E x200, b: H&Ex400).
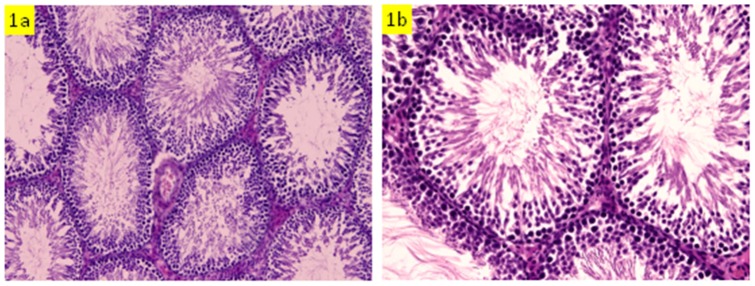
Figure 2. Testicular sections of HgCl2-exposed group.
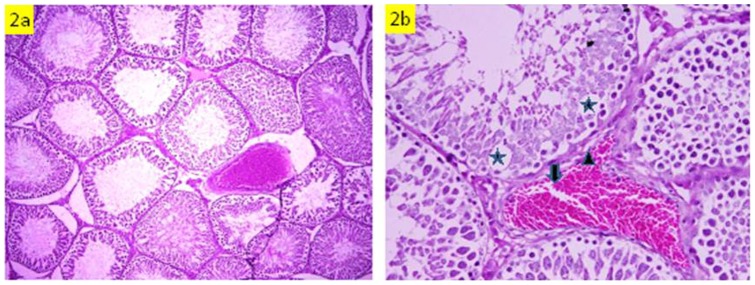
Note impaired spermatogenesis in some seminiferous tubules, degenerative areas in spermatogenic layers (Star), irregular vacuolized basement membrane (head of arrow) and hemorrhage in interstitial space (Arrow). (a: H&E ×200, b: H&Ex400).
Figure 3. Testicular section of Spirulina+HgCl2 group which show normal spermatogenesis and cell arrangement (a: H&E ×300, b: H&E×400).
Discussion
Several in vivo and in vitro studies have reported that the exposure of animals to inorganic or organic forms of mercury are accompanied by induction of oxidative stress [43] and elevation of production of reactive oxygen species (ROS) which lead to cell death [44]. Oxidative damage may result from decreased clearance of ROS by scavenging mechanisms. In the present study, HgCl2 exposure correlated with increased levels of oxidative stress and antioxidant biomarkers in the testis of rats, represented by decreased levels of GSH along with antioxidant enzymes (SOD, CAT, and GPx) and the increase of testicular MDA and blood hydroperoxide (Tables 1 & 2). MDA is one of the major products of peroxidized polyunsaturated fatty acids and increased MDA content is an important indicator of lipid peroxidation. It has been shown previously that HgCl2 increase MDA level in testicular tissue [12]. Treatment of rats with a combination of Spirulina and HgCl2 attenuated the HgCl2– induced increase in MDA level, which indicates that Spirulina may have a beneficial effect in reducing HgCl2 toxicity. Moreover, Spirulina reduced the accumulation of mercury in the testis. It has been reported that Spirulina contains vitamin E and selenium (Se) which play a crucial rule in hindering the absorption of mercury in the gastrointestinal tract [11], [45]. The most important role for Se as an essential trace element is acting as an antioxidant agent because it an essential component of the selenoprotein glutathione peroxidase (GPx). Cell membranes are protected by GPx from ROS-mediated oxidative damage resulting from the formation of hydrogen peroxide (H202) during normal metabolism in the cells' mitochondria. In this context, GPx converts H202 into water before it can produce damaging ROS. Vitamin E works in a similar way to prevent ROS-mediated membrane damage by binding up ROS within the cell membrane. In this way, Se and vitamin E act in concert to prevent damage to cell membranes, DNA and other cellular structures from damage by free radicals [46].
Various mechanisms have been proposed to explain the biological toxicity of HgCl2, including oxidative stress. Numerous studies on mercury toxicities have revealed that it generates ROS and has a great affinity for SH groups of biomolecules, thus depleting intracellular thiols including reduced glutathione [14]. It is postulated that the antioxidant GSH depletion by mercury may be a trigger for the production of reactive oxygen species (ROS) that induce lipid, protein, and DNA oxidation. Generation of ROS in the cytoplasm of cells may increase the hydrogen peroxide production and lipid peroxidation of mitochondrial membrane, resulting in loss of membrane integrity and finally cell necrosis or apoptosis [47]–[49]. In the present investigation, Mercury administration depleted GSH content in testicular tissues that made spermatogenic cells more susceptible to oxidative damage, especially during increased free radical production. GSH is an important intracellular antioxidant that spontaneously neutralizes several reactive oxygen species [50]. In the present investigation, Spirulina elevated testicular content of GSH. It has been reported that GSH, both as a carrier of mercury and an antioxidant agent, has specific roles in protecting the body from mercury toxicity. Glutathione, specifically binds with methyl mercury, forms a complex that prevents mercury from binding to cellular proteins and causing damage to both enzymes and tissue [51]. Glutathione–mercury complexes also reduce intracellular damage by preventing mercury from entering tissue and cells, and becoming an intracellular toxin [51].
Cells have a number of mechanisms to protect themselves from the toxic effect of ROS. SOD removes superoxide radical by converting it into H2O2 which is rapidly converted to water by CAT or GPx. Moreover, GPx reduces lipid hydroperoxides to alcohols. Therefore, any alteration in the activity of these enzymes may result in a number of deleterious effects due to accumulation of superoxide radicals and hydrogen peroxide. In the present study, the activities of antioxidant enzymes, SOD, CAT, and GPx in testicular tissues were concomitantly restored to normal level by Spirulina supplementation to HgCl2, which is indicative of the ROS-scavenging role that Spirulina could play whereby lessening tissue damage and subsequently improving the activities of these antioxidant enzymes. In Spirulina+HgCl2 group, Spirulina ameliorated the antioxidant status of testicular tissues in terms of increased GSH and antioxidant enzymes with decreased MDA contents. Hence, it could be concluded that Spirulina might have a potential role in preventing Hg-Cl2-induced testicular injuries. It could also be proposed that the beneficial effects of Spirulina may be due to its antioxidant properties that could revive endogenous cellular antioxidant defense system. Spirulina is considered a valuable additional food source of some macro- and micro-nutrients including high-quality protein, iron, gamma-linolenic fatty acids, carotenoids and vitamins [52]. The metalloprotective role of Spirulina may be attributed to the presence of potent antioxidant components as β-carotene, vitamin C, E, selenium and brilliant blue polypeptide pigment phycocyanin. It has been reported that β-carotene of Spirulina may reduce cell damage, especially the damage to DNA molecules, thus playing the role in the repair of regeneration process of damaged cells [53]. β- Carotene of Spirulina may scavenge free radicals generated by HgCl2, thus reducing lipid peroxidation. The antioxidant mechanism of β -carotene has been suggested to be occur via quenching of singlet oxygen, free radical scavenging and chain breaking during lipid peroxidation [54]. Vitamin E prevents lipid peroxidation and maintains GSH and ascorbic acid levels in damaged tissue by inhibiting free radical formation [55]. Moreover, these findings have been recently supported by Al-Attar [56] who showed that the administration of vitamin E protects against heavy metals-induced renal and testicular oxidative stress and injuries. Vitamin C reduces the chemical toxicity by decreasing the covalent binding of reactive intermediates and eliminating free radical metabolites [57]. Selenium present in Spirulina induces the selenium-containing enzyme GPx, proteins or compounds such as selenodiglutathione, selenocysteine and dimethylselenide, which are known to modulate the toxic effects of heavy metals [58]. It has been reported that phycocyanin content of Spirulina significantly inhibited peroxyl radical-induced lipid peroxidation [59].
Concerning the hormone level, a fall in the plasma testosterone level in the animals treated with mercury was observed (Table 3). The decrease in the testosterone level seems to be due to a reduction in the activity of enzymes involved in the biosynthesis of testosterone [60] or due to the decrease in testicular cholesterol, a precursor of testosterone synthesis. After mercury treatment, sperm concentration in the epididymis decreased (Table 5), perhaps due to a low level of sperm production in the testis, which could be related to a low level of testosterone, a prime regulator for sperm production [61]. As growth of accessory sex glands requires testosterone [62], the decrease in the weight of these glands due to mercury treatment can result from the reduction in the testosterone level (Table 4). The decrease in epididymal sperm concentration is consistent with histologic examination of the seminiferous tubules, which showed a decrease in luminal spermatozoa in HgCl2-exposed rats (Figure 2). In the present investigation, reduction in sperm number/epididymis weight and motility was associated with an increase of sperm abnormalities in rats exposed to HgCl2, which suggests the HgCl2 may interfere with spermatogenesis by crossing the blood – testis barrier and gaining access to germinal cells. The adverse effects of HgCl2 on mammalian testicular tissue have been reported with marked testicular spermatogenic degeneration at the spermatocyte level in rats [7]. The spermatozoa membranes are rich in polyunsaturated fatty acids, so they are susceptible to ROS attack and lipid peroxidation [63] as a result of exposure to mercury. Lipid peroxidation reaction causes membrane damage which leads to a decrease in sperm motility, presumably by a rapid loss of intracellular ATP, and an increase in sperm morphology defects [64], [65]. Mercury compounds have been reported to cause DNA breaks by means of free-radical mediated reactions [44] that may cause the increase in the frequency of spermatozoa with abnormal heads. Several active components in Spirulina [54], [55], [59] may scavenge ROS generated by mercury, reduce lipid peroxidation and enhance the activity of antioxidant enzymes whereby leading to protection against mercury-induced testicular damages which are manifested by an increase in sperm abnormalities and fall in testosterone level. Thus, the antioxidative properties of Spirulina may play a positive role in the defense against oxidative stress induced by HgCl2.
It has been observed that the addition of S. platensis to a diet in boars leads to an increase in sperm concentration, viability and lactate dehydrogenase activity, which has been correlated with sperm motility [44]. The present investigation clearly demonstrated that the treatment of rats with S. platensis positively impact upon sperm quality parameters, as manifested by an increase of sperm motility and count. The improvement of sperm quality may be due to the antioxidant components of Spirulina, such as α – tochopherol (vitamin E), ascorbic acid (vitamin C) and selenium that improve testicular functions and sperm quality [66], [67]. Vitamin C is a well-known antioxidant that is present in the testis protecting it from oxidative damage [68]. Thus, it has been recently reported that the decrease in the testicular level of vitamin C are correlated with methylparathion-mediated effects on sperm quality and count in rats [42].
Moreover, ß-carotene, a component of Spirulina, has been reported to enhance reproductive functions and growth in mice [69]. Moreover, scientific studies have shown that the synergistic action exhibited by a broad spectrum of antioxidants is more efficient than the activity of a single antioxidant. In addition, antioxidants from natural sources (primarily food) have a higher bioavailability and, therefore, higher protective efficacy than the synthetic ones [70].
In conclusion, the study demonstrates that S. platensis may protect testes against HgCl2-induced testicular damage, as evidenced by its effective restoration potency of oxidative stress markers, activities of enzymatic antioxidant biomarkers and histopathological alterations back to control level. Hence, it might be postulated that the protection observed by Spirulina could be attributed to its rich antioxidants content whereby scavenging HgCl2-induced ROS.
Funding Statement
The authors extend their appreciation to the Deanship of Scientific Research at King Saud University for funding the work through the research group project No RGP-VPP-130. The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Allouche L, Hamadouche M, Touabti A (2009) Chronic effects of low lead levels on sperm quality, gonadotropins and testosterone in albino rats. Exp Toxicol Pathol 61: 503–510. [DOI] [PubMed] [Google Scholar]
- 2.Gunn S, Gould T (1970) Cadmium and other mineral elements. In: Johnson AD, Gomes WR, Vademark NL, editors. The testis, vol.111. Influencing factors. New York: Academic Press p. 378–481.
- 3. Anderson M, Pedigo N, Katz R, George W (1992) Histopathology of testis from mice chronically treated with cobalt. Reprod Toxicol 6: 41–50. [DOI] [PubMed] [Google Scholar]
- 4. Ekstrand J, Bjorkman L, Edlund C, Sandborgh E (1998) Toxicological aspects on the release and systemic uptake of mercury from dental amalgam. Euro J Oral Sci 106: 678–686. [DOI] [PubMed] [Google Scholar]
- 5. Fossato da Silva D, Teixeira C, Scarano W, Favareto A, Fernandez C, et al. (2011) Effects of methyl mercury on male reproductive functions in Wistar rats. Reprod Toxicol 31: 431–439. [DOI] [PubMed] [Google Scholar]
- 6. Khan A, Atkinson A, Graham T, Thompson S, Ali S, et al. (2004) Effects of inorganic mercury on reproductive performance of mice. Food Chem Toxicol 42: 571–577. [DOI] [PubMed] [Google Scholar]
- 7. Vachhrajani K, Makhija S, Chinoy N, Chowdhury A (1988) Structural and functional alterations in testis of rats after mercuric chloride treatment. J Reprod Bio Compara Endocrinol 8: 97–104. [Google Scholar]
- 8. Rao M (1989) Histophysiological changes of sex organs in methyl mercury intoxicated mice. Endocrinologia Experimentalis 23: 60–65. [PubMed] [Google Scholar]
- 9. Mohamed M, Burbacher T, Mottet M (1987) Effects of methyl mercury on testicular functions in Macaca fascicularis monkey. Pharmacol Toxicol 60: 29–36. [DOI] [PubMed] [Google Scholar]
- 10. Rao M, Gangadharan B (2008) Antioxidative potential of melatonin against mercury induced intoxication in spermatozoa in vitro. Toxicol in vitro 22: 935–942. [DOI] [PubMed] [Google Scholar]
- 11. Su L, Wang M, Yin S, Wang H, Chen L, et al. (2008) The interaction of selenium and mercury in the accumulations and oxidative stress of rat tissues. Ecotoxicol Environ Safety 70: 483–489. [DOI] [PubMed] [Google Scholar]
- 12. Boujbiha MA, Hamden K, Guermazi F, Bouslama A, Omezzine A, et al. (2009) Testicular toxicity in mercuric chloride treated rats: Association with oxidative stress. Reprod Toxicol 28: 81–89. [DOI] [PubMed] [Google Scholar]
- 13. Lorschieder FL, Vimy MJ, Summers AO (1995) Mercury exposure from “silver” tooth filling: emerging evidence questions a traditional dental paradigm. FASEB J 9: 504–508. [PubMed] [Google Scholar]
- 14. Hansen JM, Zhang H, Hones DP (2006) Differential oxidation of thio-redoxin-1, thioredoxin-2, and glutathione by metal ions. Free Radic Biol Med 40: 138–145. [DOI] [PubMed] [Google Scholar]
- 15. Valko M, Morris H, Cronin MTD (2005) Metals, toxicity and oxidative stress. Curr Med Chem 12: 161–208. [DOI] [PubMed] [Google Scholar]
- 16. Agarwal A, Saleh RA, Bedaiwy MA (2003) Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil Steril 79: 829–43. [DOI] [PubMed] [Google Scholar]
- 17. Diemer T, Allen JA, Hales KH, Hales DB (2003) Reactive oxygen disrupts mitochondria in MA-10 tumor Leydig cells and inhibits steroidogenic acute regulatory (StAR) protein and steroidogenesis. Endocrinol 144: 2882–2891. [DOI] [PubMed] [Google Scholar]
- 18. Popescu HI (1978) Poisoning with alkyl mercuric compounds. Br Med J 1: 23–47.620134 [Google Scholar]
- 19. Keck C, Bergman M, Ernst E, Muller C, Kliensch S, et al. (1993) Autometallographic detection of mercury in testicular tissue of infertile men exposed to mercury vapour. Reprod Toxicol 7: 469–75. [DOI] [PubMed] [Google Scholar]
- 20.Cohen Z (1997) The chemicals of Spirulina, In: Vonshak, A. (Ed), Spirulina platensis (Arthrospira): Physiology, Cell-biology and Biotechnology, Taylor and francis Ltd, 175–204.
- 21. Peter C (2008) Antioxidant potential of Spirulina platensis preparations. Phytotherapy Res 22: 627–33. [DOI] [PubMed] [Google Scholar]
- 22. Karadeniz A, Yildiri A, Simsek N, Kalkan Y, Celebi F (2008) Spirulina platensis protects against gentamicin-induced nephrotoxicity in rats. Phytotherapy Res 22: 1506–1510. [DOI] [PubMed] [Google Scholar]
- 23. Torress-Duran PV, Ferreira-Hermosillo A, Juarez-Oropeza MA (2007) Antihyperlipemic and antihypertensive effects of Spirulina maxima in an open sample of Mexican population: a preliminary report. Lipids health disease 6: 33–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Sharma MK, Sharma A, Kumar A, Kumar M (2007) Spirulina fusiforms provides protection against mercuric chloride induced oxidative stress in Swiss albino mice. Food Chem Toxicol 45: 2412–2419. [DOI] [PubMed] [Google Scholar]
- 25. Romay C, Armesto J, Remikrez D, Gonzalez R, Ledon L, et al. (1998) Antioxidant and anti-inflammatory properties of c-phycocyanin from blue-green algae. Inflamm Res 47: 36–41. [DOI] [PubMed] [Google Scholar]
- 26. Salazar M, Martinez E, Madrigal E, Ruiz LE, Chamorro GA (1998) Subchronic toxicity in mice fed Spirulina maxima. . J Ethenopharmacol 62: 235–241. [DOI] [PubMed] [Google Scholar]
- 27. Salazar M, Chamorro GA, Salazar S, Steele CE (1996) Effect of Spirulina maxima consumption on reproduction and peri- and postnatal development in rats. Food Chem Toxicol 34: 353–359. [DOI] [PubMed] [Google Scholar]
- 28. Chamorro GA, Salazar M, Pages N (1996) Dominant lethal study of Spirulina maxima in male and female rats after short term feeding. Phytotherapy Res 10: 28–32. [Google Scholar]
- 29. Saxena PS, Kumar M (2004) Modulatory potential of Spirulina fusiformis on testicular phosphatases in Swiss albino mice against mercury intoxication. Indian J Exp Biol 42: 998–1002. [PubMed] [Google Scholar]
- 30. Simsek N, Karadeniz A, Kalkan Y, Keles ON, Unal B (2009) Spirulina platensis feeding inhibited the anemia and leucopenia-induced lead and cadmium in rats. J Hazar Mat 164: 1304–1309. [DOI] [PubMed] [Google Scholar]
- 31. Peixoto NC, Roza T, Flores EMM, Pereira ME (2003) Effects of zinc and cadmium on HgCl2-δ-ALA-D inhibition and Hg levels in tissues of suckling rats. Toxicol Lett 146: 17–25. [DOI] [PubMed] [Google Scholar]
- 32. Mathur N, Pandey G, Jain GC (2010) Male Reproductive Toxicity of Some Selected Metals: A Review. J Biol Sci 10: 396–404. [Google Scholar]
- 33. Ohkawa H, Ohishi N, Yagi K (1979) Assay of lipid peroxides in animal tissues by thiobarbituric acid reaction. Analyt Biochem 95: 351–358. [DOI] [PubMed] [Google Scholar]
- 34. Aboul-Soud MAM, Al-Othman AM, El-Desoky GE, Al-Othman ZA, Yusuf K, et al. (2011) Hepatoprotective effects of vitamin E/selenium against malathion-induced injuries on the antioxidant status and apoptosis-related gene expression in rats. J Toxicol Sci 36: 285–296. [DOI] [PubMed] [Google Scholar]
- 35. Shinyashiki M, Kumagai Y, Nakajima H, Nagafune J, Homma-Takeda S, et al. (1998) Differential changes in rat brain nitric oxide synthase in vivo and in vitro by methylmercury. Brain Res 798: 147–155. [DOI] [PubMed] [Google Scholar]
- 36. Moron MS, Depierre JW, Mannervik B (1979) Levels of GSH, GR and GST activities in rat lung and liver. Biochimica et Biophysica Acta 582: 67–78. [DOI] [PubMed] [Google Scholar]
- 37. Asada K, Takahashi M, Nagate M (1974) Assay and inhibitors of spinach superoxide dismutase. Agric Biol Chem 38: 471–473. [Google Scholar]
- 38. Aebi H (1984) Catalase in vitro . Method Enzymol 105: 121–126. [DOI] [PubMed] [Google Scholar]
- 39. Paglia DE, Valentine WN (1967) Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med 70: 158–169. [PubMed] [Google Scholar]
- 40.Zlatki S, Azaki B, Boyle GJ (1953) In:Harold V, editor. Practical clinical biochemistry. London: Arnol Heinemann Publication.
- 41. Narayana K, D'Souza U, Rao K (2002) Ribavirin induced sperm shape abnormalities in Wistar rat. Mutat Res 513: 193–196. [DOI] [PubMed] [Google Scholar]
- 42. Narayana K, Prashanthi N, Nayanatar A, Kumar HHC, Abhilash K, et al. (2005) Effects of methyl parathion (o,o-dimethyl o-4-nitropheny phosphorothioate) on rat sperm morphology and sperm count, but not fertility, are associated with decreased ascorbic acid level in the testis. Mutat Res 588: 28–34. [DOI] [PubMed] [Google Scholar]
- 43.Nogueira CW, Soares FA, Nascimento PC, Muller D, Rocha JBT (2003) 2, 3- Dimercaptopropane -1-sulfonic acid and meso-2,3-dimercaptosuccinic acid increase mercury- and cadmium-induced inhibition of σ-amino levulinate dehydratase. Toxicol 184, 85–95. [DOI] [PubMed]
- 44. Park E-J, Park K (2007) Induction of reactive oxygen species and apoptosis in BEAS-2B cells by mercuric chloride. Toxicol In Vitro 21: 789–790. [DOI] [PubMed] [Google Scholar]
- 45. Rao MV, Sharma PSN (2001) Protective effect of Vitamin E against mercuric chloride reproductive toxicity in male mice. Reprod Toxicol 15: 705–712. [DOI] [PubMed] [Google Scholar]
- 46. Al-Othman AM, Al-Numair KS, El-Desoky GE, Yusuf K, Al-Othman ZA, et al. (2011) Protection of α -tocopherol and selenium against acute effects of malathion on liver and kidney of rats. Afr J Pharm Pharmaco 10: 1263–1271. [Google Scholar]
- 47. Jezek P, Hlavata L (2005) Mitochondria in homeostasis of reactive oxygen species in cell, tissues, and organism. Int J Biochem Cell Biol 37: 2478–503. [DOI] [PubMed] [Google Scholar]
- 48.Kaur P, Aschner M, Syversen T (2006) Glutathione modulation influences methyl mercury induced neurotoxicity in primary cell cultures of neurons and astrocytes. Neurotoxicol 2:, 492–500. [DOI] [PubMed]
- 49. Oh IS, Datar S, Koch CJ, Shapiro IM, Shenker BJ (1997) Mercuric compounds inhibit human monocyte function by inducing apoptosis: Evidence for formation of reactive oxygen species, development of mitochondrial membrane permeability transition and loss of reductive reserve. Toxicol 124: 211–224. [DOI] [PubMed] [Google Scholar]
- 50.Lu SC (1999) Regulation of hepatic glutathione synthesis: current concepts and controversies. FASEB J 1:, 1169–1175. [PubMed]
- 51. Kromidas L, Trombetta LD, Jamall IS (1990) The protective effects of glutathione against methyl mercury cytotoxicity. Toxicol Let 51: 67–80. [DOI] [PubMed] [Google Scholar]
- 52. Mazo VK, Gmoshinskii IV, Zilova IS (2004) Microalgae Spirulina in human nutrition. Voprosy Pitaniia 73: 45–53. [PubMed] [Google Scholar]
- 53. Luxia AS, Monica S, Ornella C, Plizzala B, Laura R, et al. (1996) Effect of b-carotene on cell cycle progression of human fibroblasts. Mutagenesis 17: 2395–2401. [DOI] [PubMed] [Google Scholar]
- 54. Gerster H (1993) Anticarcinogenic effect of common carotenoids. Inter J for Vit Nut Res 63: 93–121. [PubMed] [Google Scholar]
- 55. Garg MC, Chaudhary DP, Bansal DD (2005) Effect of vitamin E supplementation on diabetes induced oxidative stress in experimental diabetes in rats. Ind J of Exp Bio 43: 177–80. [PubMed] [Google Scholar]
- 56.Al-Attar AM (2011) Antioxidant effect of vitamin E treatment on some heavy metals-induced renal and testicular injuries in male mice. Saudi J Biol Sci 18, 63–72. [DOI] [PMC free article] [PubMed]
- 57. Traber MG, Stevens JF (2011) Vitamins C and E: Beneficial effects from a mechanistic perspective. Free Radic Biol Med 51: 1000–1013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. El-Demerdash FM (2001) Effects of selenium and mercury on the enzymatic activities and lipid peroxidation in brain, liver and blood of rats. J Environ Sci Health, Part-B 36: 489–99. [DOI] [PubMed] [Google Scholar]
- 59. Bhat VB, Madyastha KM (2000) C-phycocyanin: a potent peroxyl radical scavenger in vivo and in vitro . Biochem Biophy Res Comm 275: 20–25. [DOI] [PubMed] [Google Scholar]
- 60. McVey MJ, Cooke GM, Curran IH, Chan HM, Kubow S, et al. (2008) An investigation of the effects of methyl mercury in rats fed different dietary fats and proteins: testicular steroidogenic enzymes and serum testosterone levels. Food Chem Toxicol 46: 270–279. [DOI] [PubMed] [Google Scholar]
- 61.Steinberger E (1975) Hormonal regulation of the seminiferous tubule function. In: French FS, Hansson V, Ritzen EM, Neyfeh SN, editors. Hormonal regulation of spermatogenesis. New York: Plenum Press p. 337–352.
- 62. Barkeley MS, Goldmass BD (1977) A quantitative study of serum testosterone. Sex accessory organ growth and the development of inter male aggression in the mouse. Horm Behav 8: 208–218. [DOI] [PubMed] [Google Scholar]
- 63. Mandava V, Rao BG (2008) Antioxidative potential of melatonin against mercury induced intoxication in spermatozoa in vitro . Toxicol In Vitro 22: 935–942. [DOI] [PubMed] [Google Scholar]
- 64. De Lamirande, E, Gagnon C (1992) Reactive oxygen species and human spermatozoa. I. Effects on the motility of intact spermatozoa and on sperm axonemes; and II. Depletion of adenosine triphosphate plays an important role in the inhibition of sperm motility. J Androl 13: 368–386. [PubMed] [Google Scholar]
- 65. Kistanova E, Marchev Y, Nedeva R, Kacheva D, Shumkov K, et al. (2009) Effect of the Spirulina platensis induced in the main diet on boar sperm quality. Biotech animal husband 25(5–6): 547–57. [Google Scholar]
- 66. Mohammadi S, Movahedin M, Mowla SJ (2008) Antioxidant effects of selenium on sperm parameters and testicular structure in young and aged mice. J reprod. Infertility 9: 229–237. [Google Scholar]
- 67. Yousef MI, Abdallah GA, Kamel KI (2003) Effect of ascorbic acid and vitamin E supplementation on semen quality and biochemical parameters of male rabbits. Animal Reprod Sci 76: 99–111. [DOI] [PubMed] [Google Scholar]
- 68. Sonmez M, Turk G, Yuce A (2005) The effect of ascorbic acid supplementation on sperm quality, lipid peroxidation and testosterone levels of male Wistar rats. Theriogenology 63: 2063–2072. [DOI] [PubMed] [Google Scholar]
- 69. Nagasawa H, Konishi R, Yamamoto K, Ben-Amotz A (1989) Effects of beta-carotene-rich algae on reproduction and body growth in mice. In Vivo 3: 79–81. [PubMed] [Google Scholar]
- 70. Gey KF (1998) Vitamins E plus C and interacting co-nutrients required for optimal health. A critical and constructive review of epidemiology and supplementation data regarding cardiovascular disease and cancer. Biofactors 7: 113–174. [DOI] [PubMed] [Google Scholar]