
Figure 2. Degree of binocularity for rearing conditions (PoE model).

Binocularity was measured on a 7 point scale as in Shouval et al. [150]. Values 1 and 7 represent completely monocular responses while values in the middle correspond to at least somewhat binocular responses. (A) In the normal rearing condition, neurons had a range of binocular responses, although there were few completely monocular neurons. (B) In the stripe-reared condition, binocularity increased due to higher inter-ocular correlation caused by the reduction in off-axis spatial frequencies. Experiments have also reported varying amounts of increases in binocular responses. (C) In the orthogonal-reared condition, binocularity decreased. Experiments have also reported such a decrease. (D) In the monocular-reared condition, neurons developed responses primarily for the unoccluded eye, which led to strongly monocular responses for this eye. The primary experimental finding in this rearing condition has been the absence of responses to the occluded eye. (E) Alternating monocular rearing removes inter-ocular correlation as each eye is presented with stimuli only when the other eye is occluded. In the PoE model, this led to strongly monocular responses distributed equally between eyes. Experimentally, the primary finding has been a paucity of binocular responses, but equal responses to each eye. (F) Partial monocular rearing resulted in recovery of receptive fields for both eyes, albeit with fewer binocular neurons. Experimentally, a small amount of binocular experience has been found to result in a significant recovery of responses to the occluded eye, but also an increased degree of monocularity. (G) Strabismus decreases inter-ocular correlation, and thus led in the PoE model to increased monocularity. An increase in monocularity is the primary experimental finding of the effects of strabismus. Errorbars show the SEM. Each condition was repeated 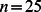 times. The binocularity distribution of all the modified rearing conditions were significantly different from the normal rearing condition (
times. The binocularity distribution of all the modified rearing conditions were significantly different from the normal rearing condition (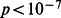 , Kolmogorov-Smirnov).
, Kolmogorov-Smirnov).