
Fig. 2.
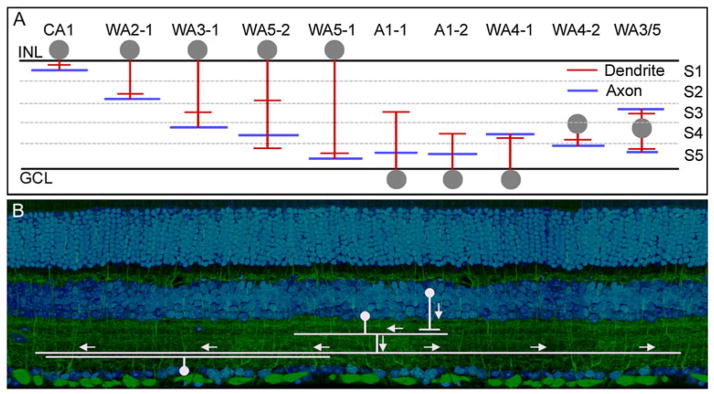
(A) Polyaxonal amacrine cells of the mouse retina. Dendrites are represented in red and axons in blue. Note that these cells are present in most layers of the IPL, and that some have axons and dendrites in different layers. It is not known whether this is an exhaustive list or whether more polyaxonal cells remain to be discovered. These cells were first well described in the retina of the rabbit (Vaney et al., 1988; Famiglietti, 1992a,b,c). Lin and Masland (2006) surveyed GFP-M retinas (Feng et al., 2000) for the presence of long-range amacrine cells. Badea and Nathans (2004) and Gustincich et al. (1997) used different genetically based strategies and their results are included here. No standard nomenclature exists; for details, see the original publications. The lateral spread of the processes is not represented, except to indicate that the spread of the dendritic arbor in these cells is narrower than the spread of the axonal arbor. (B) Polyaxonal cells transmit information from one or a few types of bipolar and amacrine cells to specific other groups of cells, here illustrated for one type of bipolar cell and one type of ganglion cell. This is shown by both the laminar structure of the polyaxonal cells, and the fact that their physiological effects are restricted to subsets of ganglion cells (Olveczky et al., 2003; Roska & Werblin, 2003).