
Figure 5. Bridging in biological capsids. Number of NA segments that directly interact with positively charged ARM segments (interaction energy , blue squares) and bridging segments (interaction energy , purple circles).
The numbers are calculated at the optimal length for each capsid shown in Figure 4 using the base-paired model. For visual reference, the dashed line indicates a 1:1 correspondence between capsid charge and number of nucleotides.
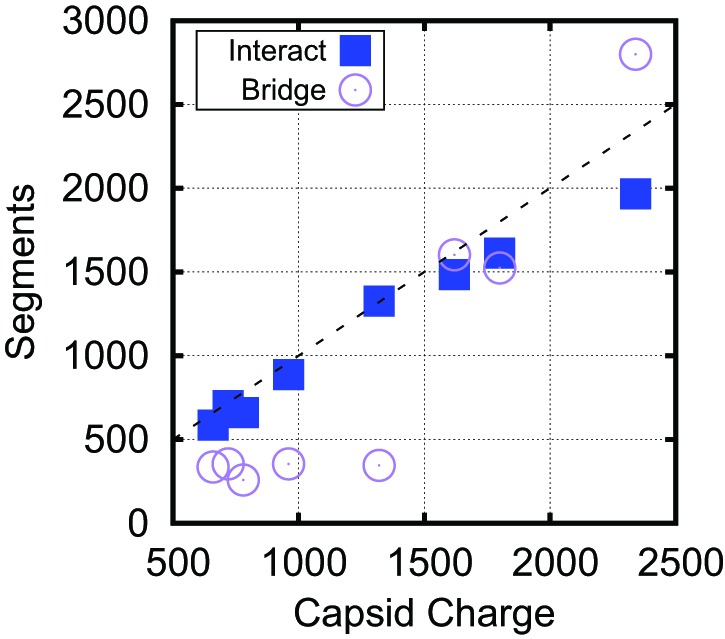
Figure 5—figure supplement 1. Radial density for linear polymer and ARM segments in the simple capsid (A) and CCMV (B).

The sharp peak in ARM density is due to the first ARM segment, which is rigidly attached to the capsid shell. In the simple capsid the polymer segments are concentrated within a few nm of the capsid shell, with lower densities in the capsid center. For CCMV, the longer arms result in a more diffuse distribution of positive charges within the capsid interior as compared to the basic capsid model. While there is some co-localization of positively charged ARM and combined neutral and negatively charged polymer segments, their densities peak at slightly different radii. The CCMV ARM sequence contains 48 segments, with 11 positive segments and 1 negative segment. Though the charges are not homogenously distributed throughout the sequence (9 occur within a 19 segment stretch), the degree of separation observed was unexpected.
Figure 5—figure supplement 2. Angular density of linear polymer segments (heat map) in the basic capsid model (A) and CCMV (B).

Green squares indicate the first ARM segment. Segment densities are averaged over radial distances of 5–6.25 nm (A) and 8.75–10 nm (B), as a function of the spherical angles, without angular averaging. For the simple capsid, the polymer more frequently resides in the vertices between subunits (between the clusters of 3 ARMs) as well as along the dodecahedral edges, and resides less frequently in the center of the subunit faces. The angular density is heterogeneous in CCMV, though to a lesser extent than found for the simple capsid.
Figure 5—figure supplement 3. Capsid radius and polymer bridging.

Number of polymer segments interacting with positive capsid charges (red inverted triangles), and number of polymer segments not interacting with positive charges (bridging segments, blue diamonds), using threshold interaction distance of 0.74 nm, which corresponds to a screened electrostatic interaction of . The numbers are calculated at the optimal polymer length as a function of capsid inradius for the simple capsid with constant ARM length (Figure 3B). The number of polymer segments strongly interacting with ARM charges is constant for nm, while the number of bridging segments increases to span the distances between arms. Hence, for capsids with nm, the observed dependence of on capsid size arises entirely due to bridging segments. For smaller capsids, there is a weak increase in the number of interacting segments with size as more conformational space around the ARMs becomes available.
Figure 5—figure supplement 4. Number of NA segments that directly interact with positively charged ARM segments and bridging segments, for both the linear and base-paired model.
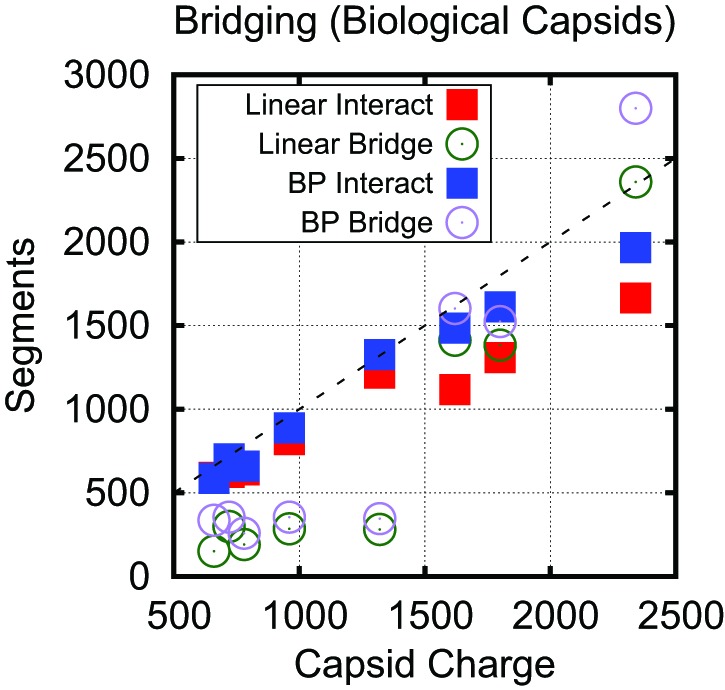
For visual reference, the dashed line indicates a 1:1 correspondence between capsid charge and number of nucleotides. This data shows that while the base-paired polymer increases the charge ratio it does so by increasing both the number of segments which are tightly bound and bridging.