

Abstract
We recently demonstrated a diurnal pattern to insulin action (i.e., insulin sensitivity [SI]) in healthy individuals with higher SI at breakfast than at dinner. To determine whether such a pattern exists in type 1 diabetes, we studied 19 subjects with C-peptide–negative diabetes (HbA1c 7.1 ± 0.6%) on insulin pump therapy with normal gastric emptying. Identical mixed meals were ingested during breakfast, lunch, and dinner at 0700, 1300, and 1900 h in randomized Latin square of order on 3 consecutive days when measured daily physical activity was equal. The triple tracer technique enabled measurement of glucose fluxes. Insulin was administered according to the customary insulin:carbohydrate ratio for each participant. Although postprandial glucose excursions did not differ among meals, insulin concentration was higher (P < 0.01) and endogenous glucose production less suppressed (P < 0.049) at breakfast than at lunch. There were no differences in meal glucose appearance or in glucose disappearance between meals. Although there was no statistical difference (P = 0.34) in SI between meals in type 1 diabetic subjects, the diurnal pattern of SI taken across the three meals in its entirety differed (P = 0.016) from that of healthy subjects. Although the pattern in healthy subjects showed decreasing SI between breakfast and lunch, the reverse SI pattern was observed in type 1 diabetic subjects. The results suggest that in contrast to healthy subjects, SI diurnal pattern in type 1 diabetes is specific to the individual and cannot be extrapolated to the type 1 diabetic population as a whole, implying that artificial pancreas algorithms may need to be personalized.
An optimal closed-loop control system will need to take into account alterations in physiological parameters that modulate glucose concentrations, including changes in insulin sensitivity (SI) related to meals and physical activity. A better understanding of how these factors are involved in glucose homeostasis is crucial to developing physiological models to improve glucose control, minimize glucose variability, and reduce morbidity and complications in individuals with diabetes, especially type 1 diabetes. We recently showed the presence of a diurnal pattern to postprandial insulin action and secretion in healthy individuals with both of these parameters to be highest at breakfast than later in the day under controlled experimental conditions (1). Although most, if not all, individuals with type 1 diabetes do not secrete insulin in response to a meal, a diurnal pattern to postprandial insulin action, if present, would need to be incorporated into an ideal closed-loop control algorithm. Furthermore, although studies investigating diurnal variations in insulin action in individuals with and without type 2 diabetes (2–8) exist, such information is scarce in type 1 diabetes. The purpose of the present study, therefore, was to determine whether there are diurnal changes in postprandial glucose tolerance, SI, and glucose fluxes in subjects with C-peptide–negative type 1 diabetes while controlling for meal macronutrient composition and caloric content and levels of physical activity. We also examined differences in the diurnal patterns of postprandial insulin action between healthy (1) and type 1 diabetic subjects while applying an identical study design in both groups. To the best of our knowledge, this study is the first to report on the application of the triple tracer method (9) in type 1 diabetes to determine postprandial glucose metabolism after mixed-meal consumption.
RESEARCH DESIGN AND METHODS
After approval from the Mayo Institutional Review Board and obtaining signed informed consent, subjects with C-peptide–negative type 1 diabetes on insulin pump therapy were enrolled. Thirteen subjects were taking insulin aspart, whereas the remaining six were taking on insulin lispro. Inclusion criteria were 18–60 years of age, body mass index of <40 kg/m2, HbA1c ≤8.5%, creatinine level ≤1.5 mg/dL, and normal gastric emptying for solids and liquids. Exclusion criteria were significant gastrointestinal symptoms by questionnaire, hypoglycemia unawareness by Clarke questionnaire, documented recent upper gastrointestinal disorder, medications affecting gastric motility (e.g., erythromycin), pregnancy or breast-feeding, or other comorbidities (e.g., nephropathy, neuropathy, macrovascular disease, hypertension) that precluded participation. Those with stable background diabetic retinopathy were included. Medications (except stable thyroid hormone or hormone replacement therapy) that could influence glucose tolerance were exclusionary. Subjects did not engage in vigorous physical activities for 72 h before the screening and study visits. Each subject underwent two screening visits.
Both healthy and type 1 diabetic subjects were studied concomitantly. The healthy subjects were studied between March 2010 and June 2011, and the diabetic subjects were studied between June 2010 and March 2012. As anticipated, it took us longer to recruit subjects with type 1 diabetes because of our stringent inclusion criteria to enroll as homogenous a cohort as possible.
Screening visit 1.
After an overnight fast, subjects reported in the morning to the Clinical Research Unit (CRU) of the Mayo Center for Translational Science Activities (CTSA) for a history, physical examination, screening laboratory tests, standard urinalysis, and resting electrocardiogram. All women of childbearing potential had a negative pregnancy test within 24 h of the study visit. A dietary history was taken to ensure adherence to a weight maintaining diet consisting of at least 200 g of carbohydrates per day and that the diet met American Diabetes Association guidelines for protein, fat, and carbohydrates. Body composition was also measured with dual-energy X-ray absorptiometry (10).
Screening visit 2.
With the use of scintigraphic techniques (11), we assessed gastric emptying for solids and liquids in all subjects who were eligible after the first screening visit, and only subjects who had normal gastric emptying proceeded to the in-patient study visit within 3 weeks of the second screening visit.
In-patient study visit.
The study interventions are as described recently (1,12). Briefly, all subjects spent 3 days and 4 nights in the CRU. Subjects reported at ∼1600 h on the evening before the first study day, when a continuous glucose sensor and physical activity monitoring system (PAMS) consisting of duplicate triaxial accelerometers were placed. They consumed a standard 10 kcal/kg meal (55% carbohydrate, 15% protein, and 30% fat) between 1700 and 1730 h. No additional food was eaten until the next morning. All subjects were provided with breakfast at 0700 h, lunch at 1300 h, and dinner at 1900 h for 3 consecutive days. Subject administered a premeal insulin bolus with their pump according to their customary insulin:carbohydrate ratio and sensitivity factor and continued with their basal insulin infusion patterns.
Study meals.
All meals were provided by the CRU metabolic kitchen. Subjects received 3 days of weighed meals (three meals each day), with each comprising 33% of the total estimated calorie intake based on Harris Benedict calorie requirements, including a low level of physical activity, with ∼50 g of carbohydrate in each meal. The meal consisted of Jell-O with dextrose, eggs (scrambled or omelet), and ham slices. A few subjects preferred steak slices to ham. The macronutrient contents for the three labeled meals and the six unlabeled meals that each subject consumed were identical. No snacks or calorie-containing drinks were permitted between meals unless otherwise required to treat hypoglycemia (point-of-care glucose ≤60 mg/dL) as per institutional guidelines. Unfinished food was weighed and excluded from calculated caloric intake. One meal daily was randomly selected per Latin square design to include 50 g glucose labeled with [1-13C] glucose in the Jell-O as the carbohydrate component. As detailed recently (1), we applied the Latin square design to maximize the time between tracer meals (i.e., minimize carryover effects). This design was specifically chosen to remove confounding effects of unequal glycogen labeling and carryover effects of residual tracer glucose concentrations in plasma on postprandial glucose fluxes that would have occurred if all three successive meals were labeled during 1 day.
Triple tracer mixed meal.
A primed continuous infusion of [6,6 2H2] glucose (11.84 mg/kg fat-free mass [FFM] prime, 0.1184 mg/kg FFM/min continuous; Masstrace, Woburn, MA) was started 3 h (−180 min) before the first bite of the mixed meal used to estimate postprandial glucose kinetics (9). Jell-O containing [1-13C] glucose was consumed along with the rest of the mixed meal of eggs and ham or steak slices. An infusion of [6-3H] glucose was started at time 0, and the rate varied to mimic the anticipated rate of appearance of the [1-13C] glucose contained within the meal. Simultaneously, the rate of infusion of [6,6-2H2] glucose was altered to approximate the anticipated pattern of change in endogenous glucose production (EGP) (10). Blood was sampled periodically for measurement of tracer:tracee ratios and glucose, insulin, and glucagon concentrations.
Physical activity protocol.
As described in detail (12), we used PAMS to capture data on body posture and movement in duplicate every 0.5 s. The subjects performed a carefully planned physical activity protocol, adherence to which was captured with the PAMS. Each labeled meal was preceded by at least 3 h and followed by 6 h of inactivity when the subjects were resting in bed to enable periodic blood draws.
Analytical techniques
Hormone analyses.
C-peptide concentration was measured on the Cobas e411 (Roche Diagnostics, Indianapolis, IN) with a two-site electrochemiluminescence immunometric assay. Insulin levels were measured by a two-site immunoenzymatic assay performed on the DxI automated immunoassay system (Beckman Coulter, Inc., Chaska, MN), and glucagon concentration was measured by a direct, double-antibody radioimmunoassay (RIA) (Linco Research, St. Charles, MO) (10). The DxI method also reliably detects both insulin aspart and insulin lispro analogs (13) that have been tested in the Mayo clinical laboratory and cross-checked in the Mayo CTSA immunochemical laboratory. Cortisol concentration was measured by a competitive binding immunoenzymatic assay on the DxI automated immunoassay system. Intraassay coefficients of variation (CVs) were 13.1, 9.4, and 6.6% at 1.56, 2.85, and 30.2 μg/dL, respectively, and interassay CVs were 9.0, 8.1, and 9.3% at 2.47, 17.3, and 27.5 μg/dL, respectively. Cortisol binding globulin (CBG) concentration was measured by competitive RIA (DIAsource ImmunoAssays S.A., Louvain-la-Neuve, Belgium). Intraassay CVs were 8.6 and 3.9% at 23 and 83 μg/mL, and interassay CVs were 10.8 and 4.8% at 25 and 114 μg/mL. Melatonin concentration was measured by competitive RIA (IBL-America, Minneapolis, MN). Intraassay CVs were 9.8, 9.7, and 13.4% at 19, 42, and 126 pg/mL, and interassay CVs were 8.0, 10.9, and 13.3% at 29, 74, and 154 pg/mL.
Glucose tracers.
Plasma samples were placed on ice, centrifuged at 4°C, separated, and stored at –80°C until assay. Plasma glucose concentration was measured by a glucose oxidase method (YSI Inc., Yellow Springs, OH). Plasma [6-3H] glucose–specific activity was measured by liquid scintillation counting as described (10). Plasma enrichment of [1-13C] glucose and [6,6-2H2] glucose were measured by gas chromatography–mass spectrometry (Thermoquest, San Jose, CA) to simultaneously quantitate C1,2 and C3–6 fragments (9).
Glucose kinetics.
Fasting and postprandial rates of glucose turnover were calculated as described (9). The systemically infused [6-3H] glucose was used to trace the systemic rate of appearance of [1-13C] glucose contained in the meal, whereas [6,6-2H2] glucose was used to trace the rate of appearance of endogenously produced glucose. The plasma concentration ratio of [6-3H] glucose to [1-13C] glucose was used to calculate the rate of appearance of ingested [1-13C] glucose, and the plasma concentration ratio of [6,6-2H2] glucose to endogenously produced glucose was used to calculate EGP. The plasma concentration of endogenously produced glucose was calculated by subtracting the concentration of exogenously derived (ingested) glucose (i.e., plasma [1-13C] glucose concentration multiplied by meal [1-13C] glucose enrichment) from total plasma glucose concentration (9).
Meal indices.
The oral glucose minimal model (14,15) was used to interpret plasma glucose and insulin concentrations measured during the meal test. The model assumes that insulin action on glucose production and disposal emanates from a compartment remote from plasma, which is usually identified with the interstitium. The most important parameter of the model, estimated from data, is net SI, which measures the overall effect of insulin to stimulate whole-body (liver and periphery) glucose disposal and inhibit glucose production. All these models have been described in detail in the online supplementary data linked to our recent report (1). However, at variance with the healthy subjects in whom data started in steady-state conditions before each meal, this was not always the case in the type 1 diabetic subjects. Thus, glucose derivative at the beginning of each meal was taken into account in the model. In addition, given that the premeal data were not used to assess basal condition, the model was also identified by these premeal data. Finally, as already observed in type 2 diabetes (10), estimating SI with precision can be difficult. Thus, to improve the numerical identifiability of the model, one must link SI to glucose effectiveness through parameter GEZI (glucose effectiveness at zero insulin). At variance with type 2 diabetes, here GEZI is not fixed to a population value in all subjects but is optimally chosen in each individual to improve model fitting.
Statistical analyses.
The experimental design translated statistically into a three treatment (meals), three period (study days) crossover study. SAS PROC MIXED (SAS Institute Inc., Cary, NC) was used to test for meal differences averaged over study periods with the methodology of Brown and Prescott (16). The likelihood of carryover effects were presumed to be negligible given the short half-life of glucose and glucose tracers and the restricted randomization process. This assumption was supported by the statistical modeling for all indices except for insulin incremental area under the curve (iAUC), where the statistical test for carryover had a P value of 0.04. However, following the recommendations of Senn (17), no further adjustment to the model (i.e., adjustment for carryover effects) was implemented. Model-based estimates (otherwise known as least squares means) were calculated to provide an average effect over the study period. Distributional assumptions for the mixed model were assessed by graphical displays of residuals and numerical summaries by meal study day. Longitudinal summary statistics (18) were used to synthesize the serial measurements into a single index. Area under the curve (AUC) and iAUC were calculated by the trapezoidal rule. Post hoc comparison of postprandial states after meals (breakfast, lunch, and dinner) were tested at the 0.05 level of significance by the Tukey-Kramer correction factor. The overall effect of meal (i.e., a type III analysis) was conducted by the Kenward-Rogers approach for determining degrees of freedom (19).
To test whether there were differences in insulin action in type 1 diabetic subjects relative to healthy subjects (1), the data were pooled so that a meal by diabetes status interaction term could be fit to the data. This interaction term, which tested for differences in profiles across meals, was the main parameter of interest. Like the models with type 1 diabetes described here, model-based estimates were computed to test for differences at each particular meal while controlling for study period. On the realization of differences in the profiles of SI between the subject cohorts, an exploratory study was conducted to examine the role of three hormones (cortisol, CBG, and melatonin) on SI at breakfast. Multiple linear regression models were used to test whether the relationship of the individual hormones were associated differently with SI between type 1 diabetic and healthy subjects. All analyses were conducted with SAS version 9.3 statistical software.
RESULTS
Subject characteristics.
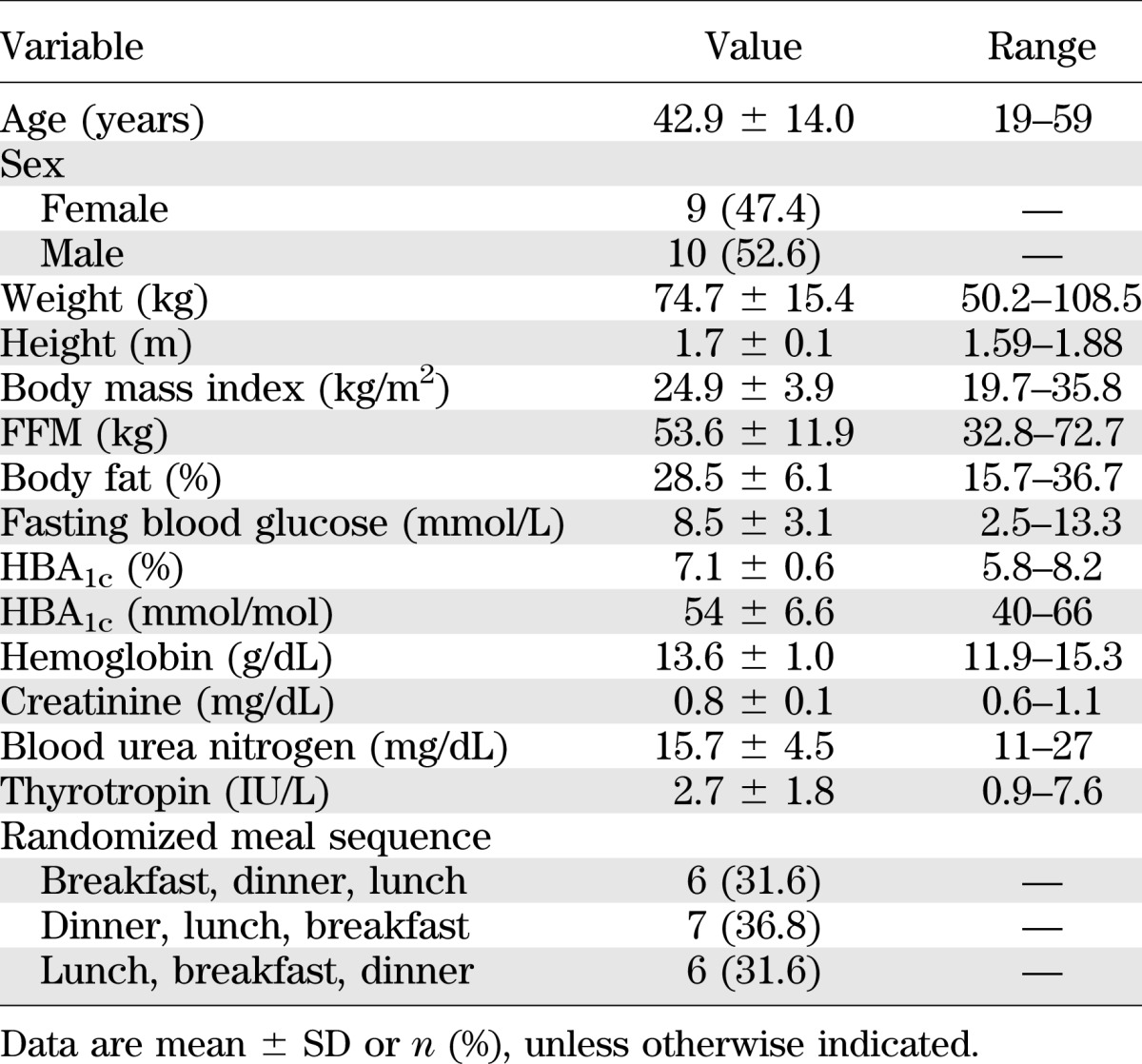
Twenty-eight subjects were screened for the study. There were five screen failures (two for abnormal gastric emptying, two for residual C-peptide secretion, and one for poor venous access) (Table 1). Three additional subjects were withdrawn after successful screening because of the inability to obtain adequate venous access during the study. The remaining 20 subjects completed all study procedures; however, one subject’s laboratory values indicated C-peptide secretion (an exclusion criterion protocol deviation) and was removed from the analysis set. Therefore, data are presented for 19 subjects. All had undetectable fasting C-peptide concentrations (<0.33 pmol/L) at the screening visit. Physical activity levels measured in accelerometer units did not differ among the 3 study days. Gastric emptying rates for solid (T½: mean ± SE 111.4 ± 4.9 min) and liquid (T½: 23 ± 3.0 min) were normal in all subjects. The meal compositions are provided in Supplementary Table 1. Only one subject did not completely ingest one meal at breakfast. The unconsumed amount (0.5 g carbohydrate, 6.0 g protein, 4 g fat, and ∼68 calories) was taken into account during the meal composition and glucose flux calculations.
TABLE 1.
Baseline characteristics of the subjects completing the three-meal study (n = 19)
Glucose, insulin, and glucagon concentrations.
Preprandial plasma glucose concentrations did not differ between meals (P = 0.083) (Fig. 1A–C and Table 2). Postprandial peak plasma glucose concentration differed by meal (P = 0.010), with it being lower at dinner than at breakfast (P < 0.01). The incremental area above baseline (0–360 min) of glucose excursions, however, did not differ between meals (P = 0.15). There were no episodes of hypoglycemia for 3 h before and 6 h after the start of the labeled meals. The plasma glucose concentrations during the labeled meals in type 1 diabetic and healthy subjects are shown together in Supplementary Fig. 1.
FIG. 1.

A: Plasma glucose concentrations obtained at breakfast, lunch, and dinner in type 1 diabetic (T1DM) subjects. B: Plasma insulin concentrations obtained at breakfast, lunch, and dinner in type 1 diabetic subjects. C: Plasma glucagon concentrations obtained at breakfast, lunch, and dinner in type 1 diabetic subjects.
TABLE 2.
Outcome measures of 3-day meal sequence
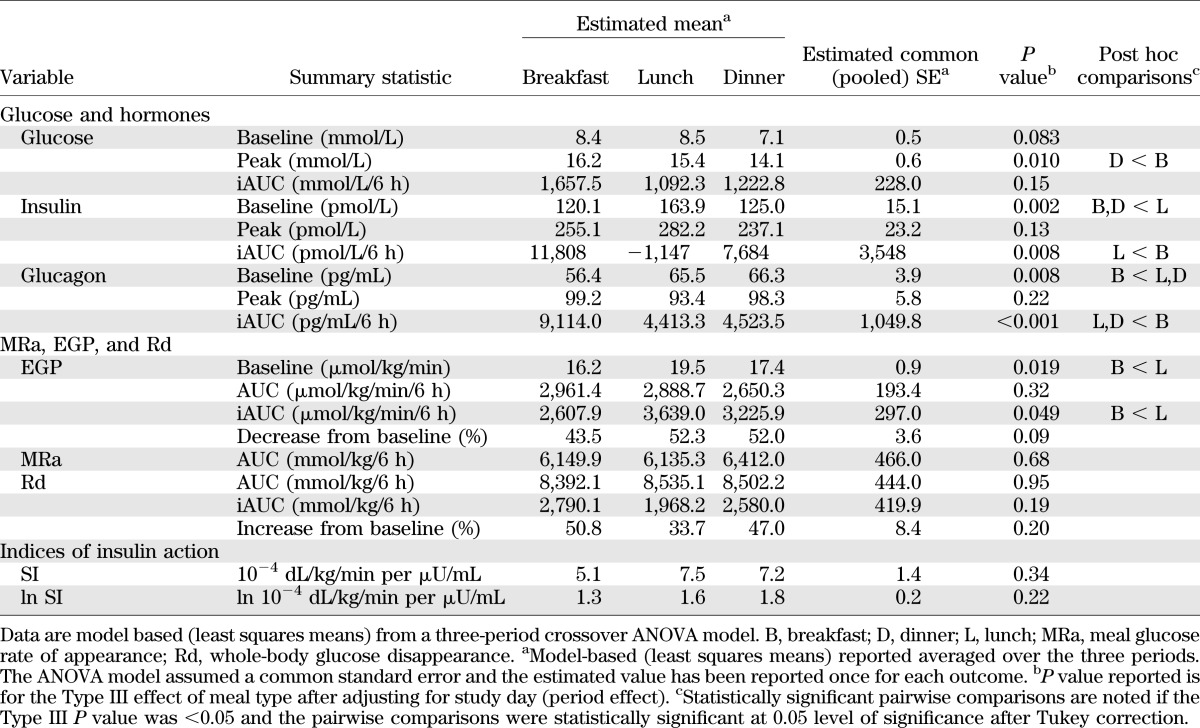
Preprandial baseline plasma insulin concentrations differed between meals (P < 0.01) with lunch being higher than either breakfast or dinner. Peak postprandial insulin concentrations, however, did not differ between meals (P = 0.13). In contrast, the incremental area above baseline (0–360 min) of insulin concentration was different by meal (P = 0.008), with it being higher at breakfast than at lunch (P < 0.01), but it did not differ between other meals.
Preprandial baseline glucagon concentrations was lower at breakfast than at lunch (P = 0.023) or dinner (P = 0.013). Although peak postprandial glucagon concentrations did not differ between meals, the incremental area above baseline (0–360 min) was higher (P < 0.01) at breakfast than at lunch or dinner.
Meal appearance, EGP, and glucose disappearance.
Meal glucose appearance did not differ among the three labeled meals (Fig. 2A–C and Table 2). Likewise, there were no detectable differences in glucose disappearance among the three meals. The integrated rates of EGP were lower at breakfast than at lunch at baseline (P = 0.02) and with the iAUC (P = 0.049). The percentage suppression (P = 0.09) and absolute AUC (P = 0.32) were not different across the meals.
FIG. 2.

A: Meal glucose appearance obtained at breakfast, lunch, and dinner in type 1 diabetic (T1DM) subjects. B: EGP obtained at breakfast, lunch, and dinner in type 1 diabetic subjects. C: Glucose disappearance obtained at breakfast, lunch, and dinner in type 1 diabetic subjects. MRa, meal glucose rate of appearance; Rd, whole-body glucose disappearance.
Insulin action.
SI, either on its natural scale or on its natural log (ln)-transformed scale, did not differ among the three labeled meals (Fig. 3A–C and Table 2). Further analyses of the pattern of SI over meals revealed a different response pattern between type 1 diabetic and healthy subjects (P = 0.016). This difference was amplified by the differences in SI at breakfast. SI was lower at breakfast in type 1 diabetic than in healthy subjects (5.1 vs. 11.2 × 10−4 dL/kg/min per μU/mL, P = 0.013), but at other meals, there was no statistically significant difference. Thus, the estimated mean response profiles (11.2–7.9–8.1 vs. 5.1–7.6–7.2 × 10−4 dL/kg/min per μU/mL for breakfast, lunch, and dinner, respectively) gave the impression that SI trended to rise from breakfast through dinner in type 1 diabetic subjects, whereas the opposite was noted in the healthy subjects, with SI falling from breakfast to dinner (1).
FIG. 3.

A: SI obtained at breakfast, lunch, and dinner in type 1 diabetic (T1DM) subjects. B: Model-based SI for all subjects for each meal shown individually. The darker line indicates the average estimates for each meal. C: Diurnal SI pattern at breakfast, lunch, and dinner obtained in healthy subjects and type 1 diabetic subjects. (A high-quality color representation of this figure is available in the online issue.)
Cortisol, CBG, and melatonin.
An exploratory analysis was conducted to describe the association of prebreakfast hormone levels with SI. This analysis was unplanned at the time of the original study, so measurements of these hormones were in subjects who had permitted the use of their stored samples for further testing (15 type 1 diabetic and 14 healthy subjects). Plasma concentrations of each hormone were assessed with the 0400 h sample (waking time before experimental procedures). The mean hormone levels were not found to differ by diabetes classification (cortisol 12.0 ± 3.1 vs. 9.2 ± 6.0 μg/dL, P = 0.25; CBG 48.6 ± 6.4 vs. 49.9 ± 6.1 μg/mL, P = 0.84; melatonin 67.5 ± 36.4 vs. 41.1 ± 35.4 pg/mL, P = 0.06 for healthy and type 1 diabetic subjects, respectively), and the association of the hormones with SI were not found to differ by diabetes classification (all diabetes by hormone interaction terms P > 0.50). Thus, although the SI diurnal pattern was different between type 1 diabetic and healthy subjects, prebreakfast hormone levels do not help to explain this difference. The numerically lower levels of cortisol and melatonin in type 1 diabetic subjects may warrant further investigation.
DISCUSSION
We show under experimental conditions controlling for meal composition and physical activity that individuals with type 1 diabetes do not demonstrate a uniformly identifiable diurnal pattern of postprandial SI over meals that could be generalized to the disease population as a whole. This finding is contrary to our recent report in a similar cohort of healthy individuals (1) and could be the result of the greater variability of SI in type 1 diabetic individuals. However, the diurnal pattern of SI over meals notably showed differences between healthy and type 1 diabetic subject groups such that in the healthy subjects, SI fell from breakfast to lunch, whereas it rose from breakfast to lunch in the type 1 diabetic subjects.
Closed-loop control algorithms currently being developed for glucose control in type 1 diabetes need to be refined and optimized on the basis of multiple considerations, including physiological parameters and characteristics that relate to natural perturbations (e.g., meals, activity) to the glucose-insulin relationship in such individuals. One such fundamental parameter that needs to be investigated is whether a diurnal pattern to postprandial glucose tolerance exists in type 1 diabetes. If indeed a diurnal pattern is clearly demonstrable in type 1 diabetes, then algorithms would need to be so informed for optimization. Few studies have attempted to investigate the existence of a diurnal pattern in type 1 diabetes. With use of the Biostator, Mathiesen et al. (20) demonstrated that insulin requirements per kilojoule consumed was higher at breakfast than at lunch, thereby implying a lower insulin action at breakfast than at lunch, but Service et al. (21) did not observe a definitive diurnal pattern of postprandial insulin requirements in type 1 diabetic subjects. However, multiple investigators have reported correlations among the total amount of carbohydrate ingested, postprandial glucose excursions, and insulin administered when using the Biostator (21–23). Furthermore and not surprisingly, other reports confirmed that glycemic excursions, both fasting and postprandial, were tighter with the Biostator, where insulin was administered intravenously, than with subcutaneous delivery of insulin (24–26).
Potential common factors that influence postprandial glucose tolerance include physical activity and meal size, calorie content, and composition. Although some (20,24,27) but not all (21–23,26) studies that used the Biostator controlled for physical activity levels, meal sizes, composition, and calorie content varied from meal to meal in all but one (27). Furthermore, all these studies used the Biostator system with intravenous insulin delivery and did not apply specific methods to measure postprandial SI.
In an effort to determine the existence of a diurnal pattern to postprandial glucose tolerance in type 1 diabetic subjects with subcutaneous open loop insulin delivery, we carefully accounted for confounding factors by controlling for size, calorie content, and composition of all meals as well as for physical activity levels by applying an identical study design as that reported recently (1). Additionally, we applied a state-of-the-art triple tracer modeling technique (9) to accurately estimate postprandial glucose fluxes and SI. Furthermore, to minimize the possibility of meal-induced endogenous insulin secretion that could dampen postprandial glucose excursions (28) and confound assessment and interpretation of postprandial glucose fluxes and SI, we enrolled a homogenous cohort of individuals with C-peptide–negative diabetes on insulin pump therapy and provided a single-wave prandial insulin bolus for every meal according to their clinically optimized insulin:carbohydrate ratio. Because there were no episodes of hypoglycemia after the labeled meals, there was no need to provide additional calories during the study.
It is noteworthy that although the numerical value of SI was lower at breakfast than at the other two meals, increased variability reduced precision within the type 1 diabetic cohort. Specifically, the residual variance for healthy subjects (1) was 11.5 × 10−4 dL/kg/min per μU/mL, and in the type 1 diabetic subjects, this variation was nearly three times larger (30.1 × 10−4 dL/kg/min per μU/mL). Additionally, there were no detectable differences in systemic appearance of meal glucose and glucose disappearance among meals. However, there was less suppression of postprandial EGP at breakfast than at lunch (despite higher postprandial insulin excursion), implying greater hepatic insulin resistance at breakfast than at lunch. A higher iAUC of postprandial glucagon concentrations at breakfast than at lunch could, at least in part, contribute to this observation. Taken together, the data suggest lower hepatic but unchanged peripheral SI contributing to the lower numerical value for SI at breakfast than at lunch or dinner.
Although the diurnal pattern of SI in type 1 diabetic subjects in the present study concords to the albeit indirect observations of Mathiesen et al. (20), important differences in study designs must be noted, including the route of insulin delivery (subcutaneously versus intravenously) and meal composition (identical versus variable). Physical activity levels, precisely measured in accelerometer units in the present study, did not differ from day to day. In Mathiesen et al., although the standardized timed physical activity was performed on a bicycle ergometer three times a day, direct estimation of glucose fluxes or SI was not done.
The difference in the diurnal trends of SI between the healthy and type 1 diabetic subjects, who were similar in their anthropometric characteristics, including percent body fat and FFM, is intriguing. As shown in Fig. 3C, the major factor that contributes to this dichotomy is the difference in SI at breakfast between the groups rather than at lunch or dinner. We do not have a ready explanation for this difference. Measurements of cortisol, CBG, and melatonin concentrations before breakfast did not reveal significant differences between the healthy and the type 1 diabetic subjects, nor were there associations of any of these hormone concentrations with SI at breakfast. Therefore, although associations of cortisol and melatonin concentrations to insulin action have been reported in the literature (29,30), we did not find such a link in the present study. Furthermore, modulation of diurnal patterns of SI by sleep architecture (31,32) could be a player and deserves further investigation. It is also noteworthy that the pattern of postprandial glucagon excursions in both healthy and type 1 diabetic subjects were similar in that the iAUC of glucagon concentration was greater at breakfast than at lunch or dinner. Although this could, at least in part, explain the higher rates of EGP in both groups at breakfast than at lunch, it does not explain per se the difference in SI patterns between healthy and type 1 diabetic subjects.
There are limitations to the study that restrict the generalizability of the findings and interpretations of the results. The highly structured protocol was used to minimize variability in levels of physical activity and meal content among subjects. Furthermore, the identical experimental design was used in healthy subjects to enable direct comparison of SI between groups. This study is the first to our knowledge where multiple tracer meals were given to type 1 diabetic subjects. The variability within and between type 1 diabetic subjects was unknown at the start of the study. The original sample size calculations anticipated an SD for ln SI of 0.36 and a difference of 0.6 between meals. The observed differences were on par with this clinically relevant difference (Table 2), but the observed SDs in type 1 diabetic subjects were much larger than anticipated. In particular, the observed SDs were 0.89, 0.59, and 1.04 for breakfast, lunch, and dinner, respectively. Therefore, a priori power was decreased, and the risk for type II error (false-negative) rate was higher than planned for. For example, a larger sample size could have resulted in statistically significant differences between meals in some of the parameters, namely EGP or SI. This limitation, however, is attenuated by the notation that the observation of larger between-subject variations suggests the need for more individualization in the artificial pancreas algorithm.
Taken together, these data in a group of individuals with C-peptide–negative type 1 diabetes demonstrate that large intersubject variability precludes a definitive diurnal pattern in SI that could be extrapolated to the type 1 diabetic population as a whole. Hence, any diurnal pattern of SI in type 1 diabetes is individual specific, suggesting that artificial pancreas algorithms need to be personalized. This observation that SI varies in a subject-specific fashion is an important finding and will be incorporated into the type 1 diabetes simulator (33,34) to allow more reliable testing of closed-loop control algorithms.
Supplementary Material
ACKNOWLEDGMENTS
This work was supported by National Institutes of Health Grants DK-R01-085561 and DK-DP3-094331 and by Grant UL1-R000135 from the National Center for Advancing Translational Sciences, a component of the National Institutes of Health. C.D.M. and C.C. are partially funded by Italian Ministero dell’Istruzione, dell’Università e della Ricerca (Progetto FIRB 2009).
No potential conflicts of interest relevant to this article were reported.
L.H., D.K.N., and A.S. contributed to the study conduct and data handling. C.D.M., A.E.B., J.A.L., R.B., R.E.C., and C.C. contributed to the study design, data analyses, and manuscript review and editing. R.A.R. contributed to the study design and manuscript review and editing. Y.C.K. and A.B. contributed to the study design, data research, and manuscript writing and editing. A.B. is the guarantor of this work and, as such, had full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
Parts of this study were presented at the 48th Annual Meeting of the European Association for the Study of Diabetes, Berlin, Germany, 1–5 October 2012.
The authors are deeply indebted to the research participants and thank the staff of the Mayo CTSA CRU; the GI Motility Core; the CRU Mass Spectroscopy Laboratory; CRU Immunochemical Core Laboratory; and Pamela Reich (research assistant), Betty Dicke, Chad Clark (laboratory technicians), Brent McConahey (research assistant), and Shelly McCrady Spitzer (research assistant) of the Endocrine Research Unit, Mayo Clinic, Rochester, Minnesota.
Footnotes
This article contains Supplementary Data online at http://diabetes.diabetesjournals.org/lookup/suppl/doi:10.2337/db12-1759/-/DC1.
See accompanying commentary, p. 2173.
REFERENCES
- 1.Saad A, Dalla Man C, Nandy DK, et al. Diurnal pattern to insulin secretion and insulin action in healthy individuals. Diabetes 2012;61:2691–2700 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Malherbe C, De Gasparo M, De Hertogh R, Hoet JJ. Circadian variations of blood sugar and plasma insulin levels in man. Diabetologia 1969;5:397–404 [DOI] [PubMed] [Google Scholar]
- 3.Van Cauter E, Shapiro ET, Tillil H, Polonsky KS. Circadian modulation of glucose and insulin responses to meals: relationship to cortisol rhythm. Am J Physiol 1992;262:E467–E475 [DOI] [PubMed] [Google Scholar]
- 4.Jarrett RJ, Baker IA, Keen H, Oakley NW. Diurnal variation in oral glucose tolerance: blood sugar and plasma insulin levels morning, afternoon, and evening. BMJ 1972;1:199–201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Morgan LM, Aspostolakou F, Wright J, Gama R. Diurnal variations in peripheral insulin resistance and plasma non-esterified fatty acid concentrations: a possible link? Ann Clin Biochem 1999;36:447–450 [DOI] [PubMed] [Google Scholar]
- 6.Peter R, Dunseath G, Luzio SD, Chudleigh R, Roy Choudhury S, Owens DR. Daytime variability of postprandial glucose tolerance and pancreatic B-cell function using 12-h profiles in persons with type 2 diabetes. Diabet Med 2010;27:266–273 [DOI] [PubMed]
- 7.Lee A, Ader M, Bray GA, Bergman RN. Diurnal variation in glucose tolerance. Cyclic suppression of insulin action and insulin secretion in normal-weight, but not obese, subjects. Diabetes 1992;41:750–759 [DOI] [PubMed] [Google Scholar]
- 8.Waldhäusl W. Circadian rhythms of insulin needs and actions. Diabetes Res Clin Pract 1989;6:S17–S24 [DOI] [PubMed] [Google Scholar]
- 9.Basu R, Di Camillo B, Toffolo G, et al. Use of a novel triple-tracer approach to assess postprandial glucose metabolism. Am J Physiol Endocrinol Metab 2003;284:E55–E69 [DOI] [PubMed] [Google Scholar]
- 10.Basu A, Dalla Man C, Basu R, Toffolo G, Cobelli C, Rizza RA. Effects of type 2 diabetes on insulin secretion, insulin action, glucose effectiveness, and postprandial glucose metabolism. Diabetes Care 2009;32:866–872 [DOI] [PMC free article] [PubMed]
- 11.Bharucha AE, Camilleri M, Forstrom LA, Zinsmeister AR. Relationship between clinical features and gastric emptying disturbances in diabetes mellitus. Clin Endocrinol (Oxf) 2009;70:415–420 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Manohar C, Levine JA, Nandy DK, et al. The effect of walking on postprandial glycemic excursion in patients with type 1 diabetes and healthy people. Diabetes Care 2012;35:2493–2499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Owen WE, Roberts WL. Cross-reactivity of three recombinant insulin analogs with five commercial insulin immunoassays. Clin Chem 2004;50:257–259 [DOI] [PubMed] [Google Scholar]
- 14.Dalla Man C, Caumo A, Cobelli C. The oral glucose minimal model: estimation of insulin sensitivity from a meal test. IEEE Trans Biomed Eng 2002;49:419–429 [DOI] [PubMed] [Google Scholar]
- 15.Dalla Man C, Caumo A, Basu R, Rizza R, Toffolo G, Cobelli C. Minimal model estimation of glucose absorption and insulin sensitivity from oral test: validation with a tracer method. Am J Physiol Endocrinol Metab 2004;287:E637–E643 [DOI] [PubMed] [Google Scholar]
- 16.Brown H, Prescott R. Applied Mixed Models in Medicine 2nd ed. West Sussex, U.K., John Wiley & Sons, 2006 [Google Scholar]
- 17.Senn SS. Cross-over Trials in Clinical Research 2nd ed. West Sussex, U.K., John Wiley & Sons, 2002 [Google Scholar]
- 18.Der G, Everitt BS. Statistical Analysis of Medical Data using SAS. Boca Raton, FL, Chapman and Hall/CRC, 2006 [Google Scholar]
- 19.Littell RC, Milliken GA, Stroup WW, Wolfinger RD, Scabenberger O. SAS for Mixed Models. Cary, NC, The SAS Institute, 2006 [Google Scholar]
- 20.Mathiesen ER, Rubin P, Sandahl Christiansen J, Aaby Svendsen P, Lauritzen T, Deckert T. Diurnal pattern of insulin requirements in insulin-dependent diabetics. Scand J Clin Lab Invest 1982;42:63–68 [PubMed] [Google Scholar]
- 21.Service FJ, Rizza RA, Hall LD, et al. Prandial insulin requirements in insulin-dependent diabetics: effects of size, time of day, and sequence of meals. J Clin Endocrinol Metab 1983;57:931–936 [DOI] [PubMed] [Google Scholar]
- 22.Slama G, Klein JC, Delage A, et al. Correlation between the nature and amount of carbohydrate in meal intake and insulin delivery by the artificial pancreas in 24 insulin-dependent diabetics. Diabetes 1981;30:101–105 [DOI] [PubMed] [Google Scholar]
- 23.Levy-Marchal C, Perlman K, Zinman B, Albisser AM. Preprogrammed intravenous insulin infusion in diabetic humans: metabolic consequences of altering meal size. Diabetes Care 1986;9:283–290 [DOI] [PubMed] [Google Scholar]
- 24.Zinman B, Stokes EF, Albisser AM, et al. The metabolic response to glycemic control by the artificial pancreas in diabetic man. Metabolism 1979;28:511–518 [DOI] [PubMed] [Google Scholar]
- 25.Albisser AM, Leibel BS, Ewart TG, et al. Clinical control of diabetes by the artificial pancreas. Diabetes 1974;23:397–404 [DOI] [PubMed] [Google Scholar]
- 26.Nosadini R, Noy GA, Nattrass M, et al. The metabolic and hormonal response to acute normoglycaemia in type 1 (insulin-dependent) diabetes: studies with a glucose controlled insulin infusion system (artificial endocrine pancreas). Diabetologia 1982;23:220–228 [DOI] [PubMed] [Google Scholar]
- 27.Service FJ. Normalization of plasma glucose of unstable diabetes: studies under ambulatory, fed conditions with pumped intravenous insulin. J Lab Clin Med 1978;91:480–489 [PubMed] [Google Scholar]
- 28.Werther GA, Turner RC, Jenkins PA, Baum JD. Twenty-four hour profiles of plasma C-peptide in type 1 (insulin-dependent) diabetic children. Diabetologia 1982;22:245–249 [DOI] [PubMed] [Google Scholar]
- 29.Bouatia-Naji N, Bonnefond A, Cavalcanti-Proença C, et al. A variant near MTNR1B is associated with increased fasting plasma glucose levels and type 2 diabetes risk. Nat Genet 2009;41:89–94 [DOI] [PubMed] [Google Scholar]
- 30.Agil A, Rosado I, Ruiz R, Figueroa A, Zen N, Fernández-Vázquez G. Melatonin improves glucose homeostasis in young Zucker diabetic fatty rats. J Pineal Res 2012;52:203–210 [DOI] [PubMed] [Google Scholar]
- 31.Jauch-Chara K, Schmid SM, Hallschmid M, Born J, Schultes B. Altered neuroendocrine sleep architecture in patients with type 1 diabetes. Diabetes Care 2008;31:1183–1188 [DOI] [PubMed] [Google Scholar]
- 32.Tasali E, Leproult R, Ehrmann DA, Van Cauter E. Slow-wave sleep and the risk of type 2 diabetes in humans. Proc Natl Acad Sci U S A 2008;105:1044–1049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kovatchev BP, Breton M, Man CD, Cobelli C. In silico preclinical trials: a proof of concept in closed-loop control of type 1 diabetes. J Diabetes Sci Tech 2009;3:44–55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kovatchev B, Patek S, Dassau E, et al. Juvenile Diabetes Research Foundation Artificial Pancreas Consortium Control to range for diabetes: functionality and modular architecture. J Diabetes Sci Tech 2009;3:1058–1065 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.