

Abstract
The effects of storage and deep desiccation on structural changes of DNA in orthodox seeds are poorly characterized. In this study we analyzed the 5-methylcytosine (m5C) global content of DNA isolated from seeds of common pear (Pyrus communis L.) that had been subjected to extreme desiccation, and the seedlings derived from these seeds. Germination and seedling emergence tests were applied to determine seed viability after their desiccation. In parallel, analysis of the global content of m5C in dried seeds and DNA of seedlings obtained from such seeds was performed with a 2D TLC method. Desiccation of fresh seeds to 5.3% moisture content (mc) resulted in a slight reduction of DNA methylation, whereas severe desiccation down to 2–3% mc increased DNA methylation. Strong desiccation of seeds resulted in the subsequent generation of seedlings of shorter height. A 1-year period of seed storage induced a significant increase in the level of DNA methylation in seeds. It is possible that alterations in the m5C content of DNA in strongly desiccated pear seeds reflect a reaction of desiccation-tolerant (orthodox) seeds to severe desiccation. Epigenetic changes were observed not only in severely desiccated seeds but also in 3-month old seedlings obtained from these seeds. With regard to seed storage practices, epigenetic assessment could be used by gene banks for early detection of structural changes in the DNA of stored seeds.
Introduction
Plants modulate their physiology and development through genome-wide changes in gene expression in response to environmental conditions [1]. The mechanisms of desiccation tolerance of orthodox (desiccation tolerant) seeds are well known. These mechanisms include the accumulation of late embryogenesis abundant proteins (LEA), dehydrins [2], non-reducing sugars, sucrose [3], selenium [4], activation of antioxidants [5], [6], intracellular dedifferentiation [7], metabolic ‘switching off’ [8], and the operation of repair systems [9], [3]. Although a number of phenomena, as discussed above, have been implicated in the acquisition and maintenance of desiccation tolerance, a coherent view integrating control of the acquisition of desiccation tolerance still remains to be elucidated [10].
It has been recently shown that epigenetic regulation of gene expression is involved in plants’ response to environmental stresses (i.e. drought stress) [1]. The most extensively studied and characterized epigenetic modification of DNA is the methylation of cytosine, which represents the addition of a methyl group to carbon 5 (C5) of the pyrimidine ring [11]. DNA methylation in plants predominantly occurs at CG dinucleotides, but also occurs at CHG (where H is A, C or T) and asymmetric CHH (where H is A, C or T) sites [12].
At the present time, the relationship of this epigenetic modification to the adaptation of seeds to environmental stress is not entirely understood. Nevertheless, it is apparent that plants modulate their physiology and development through epigenetic modifications of their DNA [1]. Specifically, DNA methylation and histone modification, play a crucial role in the regulation of gene expression during the responses of mature plants to environmental stresses [13], [14], [15]. Furthermore, these modifications can be inherited by subsequent generations [16], [17]. In plants, the methylation of symmetric (CG, CHG) and asymmetric (CHH) sequences is maintained by DNA methyltransferases: Methyltransferase 1 (MET 1) [18], [19], [20], Chromomethylase3 (CMT3) [21] and DNMT3a/3b homologs: Domains Rearranged Methylase 1 and 2 (DRM1/2), which require an active guiding by small interfering RNAs (siRNAs) to target DNA [22], [23].
Orthodox seeds are dried to specific moisture content before they are stored in gene banks that have been established to preserve the seeds under conditions that ensure genetic stability [24]. We hypothesize that seed moisture content and temperature affect the course of epigenetic changes, including the global methylation of genomic DNA. Understanding the stresses that affect the overall level of DNA methylation in seeds and during seedling development is crucial for effective storage of genetic resources in ex situ gene banks.
Most of the stress-induced alterations in epigenetic modification of DNA are reset upon relief from the stress. However, in non-Mendelian inheritance, some of the modifications are stable and may be carried forward in the form of a ‘stress memory’ that is inherited across mitotic or even meiotic cell divisions. It is thought that epigenetic stress memory may help plants to cope more effectively with subsequent stresses [25]. However, the effects of stresses on the level of DNA methylation in seeds have not been previously analyzed, and it is not known whether epigenetic changes are induced. We hypothesized that DNA methylation levels are altered at the whole-genome level, as well as at the level of individual genes, in response to environmental stresses. It is likely that alterations in the global level of DNA methylation could play an important role in the response to desiccation of orthodox seeds. It may be relevant because they typically undergo deep desiccation in gene banks to inhibit their aging and prolong storage.
In the present study, we analyzed the effects of desiccation and storage on the global level of DNA methylation of common pear (Pyrus communis L.) seeds. In addition, the abundance of m5C in seedlings obtained from these seeds was also analyzed. Influence of desiccation on orthodox seed germination, seedling emergence and growth was assessed as well.
Materials and Methods
Ethics Statement
No specific permits were required for the described experimental field studies: a) no specific permissions were required for these locations b) locations are not privately-owned or protected; c) the field studies did not involve endangered or protected species.
Plant Material
Seeds of common pear (Pyrus communis L.) were collected when ripe after abscission (green turning to yellow) in 2007 from Łopuchówko (52.6151°N, 17.0896°E; 30 km north of Poznań) and in 2009 from Borkowice (52.2167°N, 16.8000°E; 30 km south of Poznań), Poland. Seeds were extracted manually from fruits. Collections from Borkowice were represented by fresh seeds (non-stored) and seeds stored for 1 year. For collections from Łopuchówko, only seeds stored for 2 years were used in the experiment. For both provenances, partially dried seeds were stored at 3°C.
Seed Moisture Content
After harvest, seeds were dried on a laboratory bench at 20°C to a moisture content (mc) of 8.2–8.5% (safe mc, used later as a control for severely desiccated seeds of mc 2.2–2.8% from both provenances). Desiccation to the lowest levels of seed mc, i.e. 2.2% and 2.8%, was performed over activated silica gel in a tightly closed vessel. The required seed mass, corresponding to the measured mc, was calculated using the following formula [26]:
where X = final mass of the seeds
M = fresh mass of the seeds
Wp = moisture content of seeds before drying
Wd = desired moisture content of seeds.
Seeds were placed in a drying box on blotting paper (in a layer with a thickness not exceeding two times the seed height) and desiccated for 14 d. The mc of seeds was assessed with three replications of 10 seeds each by drying at 103°C±2°C for 18 h. Moisture content was expressed as a percentage on a fresh-weight basis. Stratification of the dormant seeds was required before subsequent germination and seedling emergence tests.
Stratification
The seeds were placed in plastic boxes in a moist substrate mixture (1∶1, v/v) of quartz sand (<1 mm fraction) and sieved peat with pH 3.5–4.5 [27]. Seeds were mixed with the substrate (1∶3, v/v) and placed in 0.25 L plastic bottles. Seeds were monitored for fungal infections and the first germinated seeds were counted. Specifically, seeds were visually assessed for the emergence of a radicle (2–3 mm long), which is a visible indicator of release from dormancy. The substrate was watered weekly if required and seeds were stratified for 16 weeks at 3°C, until no more than 5% of seeds had developed a visible radicle. Subsequently, the seeds were transferred to a cyclically alternating temperature regime and subjected to a germination test [26].
Germination, Seedling Emergence Test, and Seedling Growth
Stratified seeds were germinated in darkness, in an identical substrate mixture that used for stratification. Optimum thermal conditions for seed germination were ensured by cyclically alternating temperatures of 3°C/20°C (16 h/8 h per day, respectively). Under these conditions, a seed with a 3 mm long radicle was considered as “germinated”.
After the germination test, a final evaluation of seed viability was performed and seeds were dissected for visually assessment. The seeds were cut with a scalpel along the longitudinal axis, across the cotyledons and the embryonic axis. All seeds that failed to germinate by the end of the testing period were classified as “decayed seeds”.
Seedling emergence was also evaluated with an identical substrate that was used for stratification and the germination tests. Stratified seeds were sown in plastic boxes in the substrate at a depth of 1 cm, and covered with a layer of sand. The boxes were covered with a transparent lid to enable the penetration of light to emerging seedlings and to maintain a suitable moisture level. The lid was subsequently removed when seedlings were approximately 2–3 cm high. Thermal conditions were identical to the germination test (3°C/20°C, 16 h/8 h per day) until the emergent seedlings were approximately 2–3 cm high. Boxes with seedlings were then moved into a controlled growth chamber at 25°C under light conditions (16 h/8 h photoperiod at 60 µmolm−2s−1) and maintained for 4–5 weeks.
For epigenetic analysis (determination of the m5C content in leaf DNA), seedlings were grown for 3 months under a 16 h/8 h photoperiod at 77 µmol m−2 s−1at 20°C. Each experiment was comprised of four replicates, containing 50 seeds each.
The height of 3-month-old seedlings was measured for seeds that were harvested from the Borkowice provenance. The seeds were stored for 1 year and had an 8.5% mc. After storage, they were desiccated to 2.2% mc and used for subsequent seedling growth assays. Shoot height (mm) was measured from the root collar to the shoot apex of germinated seedlings. Each experiment was replicated 4 times and contained 25 seedlings per replicate experiment.
DNA Isolation and Assessment of Global DNA Methylation Levels
Total genomic DNA was extracted from embryos (after removal of the seed coat) and leaves of seedlings with the Qiagen DNAeasy Plant Mini Kit (Qiagen, Hilden, Germany). One variant of the experiment (DNA extraction) consisted of 3 replicates. Each replicate consisted of five embryos. For DNA extraction from leaves, for one replicate one-quarter of an apical part of three leaves from one seedling were used. The level of m5C was measured five times for each biological replicate.
Analysis of the global content of m5C in DNA was carried out as previously described [28]. This method has been already successfully applied for the assessment of DNA methylation in plants [29]. Although it is currently possible to assess global methylation status with HPLC analyses, there are some serious technical limitations to this approach. HPLC requires a substantial amount of the starting material and DNA preparations must be RNA-free since it is a technical challenge to use column chromatography to differentiate the ribo- and deoxyribonucleoside of 5-methylcytosine. This consideration is especially challenging in case of DNA samples that are derived from seeds that are rich in storage material. A thin-layer chromatography (TLC) based method seems to be the most appropriate since it requires a small amount of DNA and ribo- and deoxyribonucleotides are easily distinguishable under this experimental testing condition. Moreover, even small changes in 5-methylcytosine amount are capable of being detected [28].
To begin our evaluation of global DNA methylation level, we first dried DNA (1 µg) and then digested with 0.001 U spleen II nuclease and 0.02 U micrococcal nuclease in 20 mM succinate buffer containing 10 mM CaCl2 for 6 h at 37°C. The hydrolysate (0.3 µg) was then labelled with 1.6 µCi [γ-32p]ATP (6,000 Ci/mmol; Hartmann Analytic, Germany) and 1.5 U T4 polynucleotide kinase in 10 mM bicine-NaOH buffer (pH 9.7) containing 10 mM MgCl2, 10 mM dithiothreitol, and 1 mM spermidine. After incubation for 30 min at 37°C, 0.03 U apyrase in 10 mM bicine-NaOH buffer was added and incubated for 30 min. Next, 0.2 µg RNase P1 in 500 mM ammonium acetate buffer (pH 4.5) was used for 3′-phosphate cleavage. The [γ-32p] m5C content was analyzed by two-dimensional thin layer chromatography on cellulose plates (Merck) in isobutyric acid: NH4OH:H2O (66∶1∶17) and 0.1 M sodium phosphate (pH 6.8)–ammonium sulfate–n-propanol (100 mL/60 g/1.5 mL, v/w/v). Radioactivity was subsequently measured using a Fluoro Image Analyzer FLA-5100 with Multi Gauge 3.0 software (FujiFilm). Each analysis was repeated for a total of four times.
For quantitative evaluation of m5C content, fluoroscopic image analyses accounted for the contents of C, T and m5C. The global level of m5C (R) was calculated with the following formula:
where I is the intensity of individual spots corresponding to the nucleotide analyzed (Figure1).
Figure 1. Assessment of global DNA methylation of seeds from Borkowice, stored for 1 year.
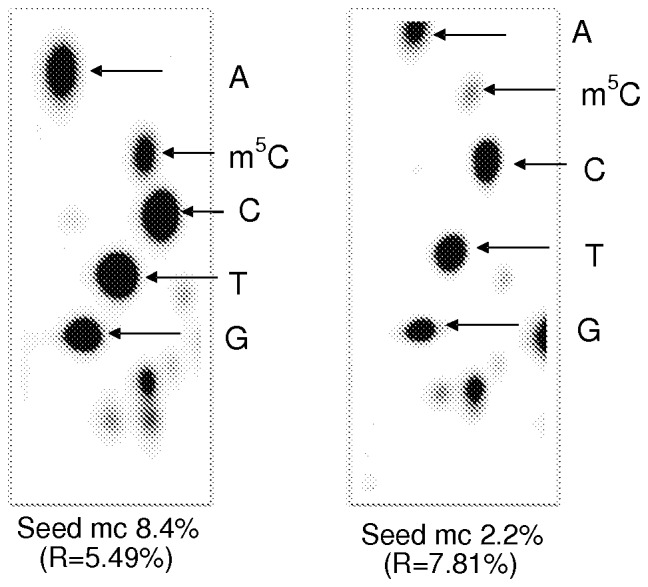
2′deoxynucleotides derived from DNA hydrolysis (labelled spots) and RNA contamination (unlabelled spots) are clearly separated. A- adenine, m5C -5-methylcytosine, C-cytosine, T-thymine, G- guanine.
Statistical Analysis
STATISTICA software, 1998 edition (StatSoft Polska, 1995–2005) was used for the statistical analyses of all data. Analysis of variance (ANOVA) was used to determine the significance of differences between means, and a Tukey’s test was used for pair-wise comparisons. The Tukey’s test was performed after arc-sine transformation and was always run at a significance level of P≤0.05. Separate ANOVAs and Tukey’s tests were performed for analysis of germination, seedling emergence, global methylation, and seedling height. Standard deviations were indicated as errors bars on graphs.
Results
The germination of control seeds, which were not deeply desiccated and had a moisture content of 8.2–8.8%, was nearly 100% (Figure 2). Additional desiccation of freshly collected seeds over silica gel to a 5.3% mc (Borkowice) or 4.8% mc (Łopuchówko) did not significantly affect seed germinability. Seeds from both provenances that were desiccated to 2.2–2.8% mc showed a 90–98% germination rate (Figure 2).
Figure 2. Effects of seed desiccation and storage on the germination of pear seeds.
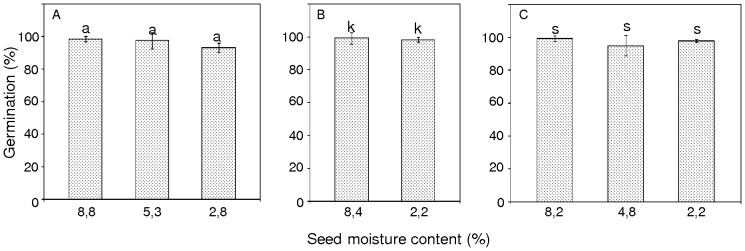
(A) Unstored seeds from Borkowice. (B) Seeds from Borkowice that had been stored for 1 year. (C) Seeds from Łopuchówko that had been stored for 2 years. Values marked with the same letter are not significantly different at P<0.05, ANOVA, Tukey`s test, error bars indicate standard deviations.
Control seeds from the two provenances with 8.2, 8.4, or 8.8% mc showed 100% seedling emergence (Figure 3). Emergence was categorized as emergent plants containing several leaves and an average height approximately 3 cm. Desiccation of seeds from the two provenances to 4.8% (2-years stored seeds) or 5.3% mc (fresh seeds) did not significantly reduce the level of seedling emergence, which remained high at 98–100% (Figure 3). Similarly, desiccation of 1-year stored seeds from Borkowice to 2.2 or 2.8% mc did not induce significant changes and a similar percentage of seedling emergence was observed (98–100%). However, desiccation of seeds from Łopuchówko to 2.2% mc resulted in a significant reduction of seedling emergence from 100 to 96% in comparison to control seeds (8.2% mc) (Figure 3). For all sets of seeds that were studied, all were considered equal in both quality and viability.
Figure 3. Effects of pear seed desiccation and storage on seedling emergence.
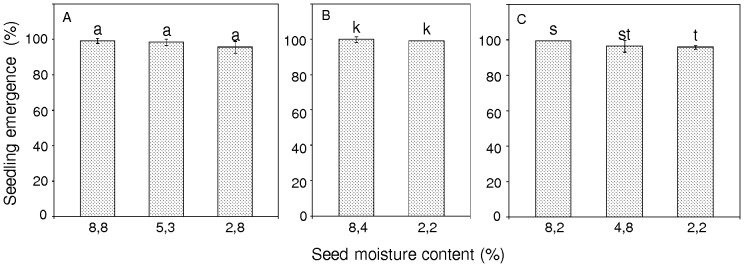
(A) Unstored seeds from Borkowice. (B) Seeds from Borkowice that had been stored for 1 year. (C) Seeds from Łopuchówko that had been stored for 2 years. Values marked with the same letter are not significantly different at P<0.05, ANOVA, Tukey`s test, error bars indicate standard deviations.
Methylation of DNA after Seed Desiccation
The global level of DNA methylation (R-percentage of methylated cytosines) was R = 3.08% for seeds that were desiccated to 8.8% mc immediately after harvest. Further drying of the seeds to 5.3% mc resulted in a slight reduction in the level of methylation to R = 2.73% (Figure 4A). Deep drying of seeds over silica gel to 2.8% mc significantly increased the level of m5C to R = 4.42% in comparison to seeds with 5.3 or 8.8% mc (Figure 4A).
Figure 4. Changes in the percentage of DNA in pear seeds that was modified to carry 5-methylcytosine after desiccation and storage.
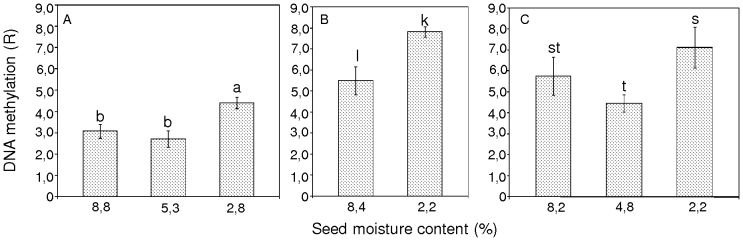
(A) Unstored seeds from Borkowice. (B) Seeds from Borkowice that had been stored for 1 year. (C) Seeds from Łopuchówko that had been stored for 2 years. Values marked with the same letter are not significantly different at P<0.05, ANOVA, Tukey`s test, error bars indicate standard deviations.
Control seeds (8.4% mc) that were stored for 1 year prior to deep desiccation contained a high level of m5C (R = 5.49%) (Figure 4B), which was higher than that of DNA in fresh seeds (8.8% mc; R = 3.08%), (Figure 4A). The drying of seeds after 1-year storage to 2.2% mc resulted in an increase in the global DNA methylation level from R = 5.49 to 7.81% in comparison to seeds that were not as deeply desiccated, 8.4% mc (Figure 4B).
Seeds from the Łopuchówko provenance (stored for 2 years at 8.2% mc) were characterized by R = 5.75% immediately after storage (Figure 4C). Drying of the seeds to 4.8% mc slightly reduced the m5C content from R = 5.75 to 4.45%, whereas further drying to 2.2% mc resulted in a significant increase in m5C content to R = 7.11% (Figure 4C). For stored seeds, the global level of m5C (R = 4.5–7.5%) (Figure 4B, C) was higher than that of non-stored seeds (R = 2.8–4.5%), (Figure 4A).
No significant differences were observed for the germination and emergence of seeds that were analyzed freshly after harvest from Borkowice (8.8% mc) or after a 1 year duration of storage (8.4% mc), (Figure 5). However, a comparison of the percentage of DNA methylation revealed a relevant time-dependent increase in m5C. After 1 year in storage, the amount of 5-methylcytosine increased from 3.08 to 5.49% (Figure 5).
Figure 5. Changes in germination (A), seedlings emergence (B) and DNA methylation level (C) in seeds from Borkowice (8.8% mc) after 12 months of storage.
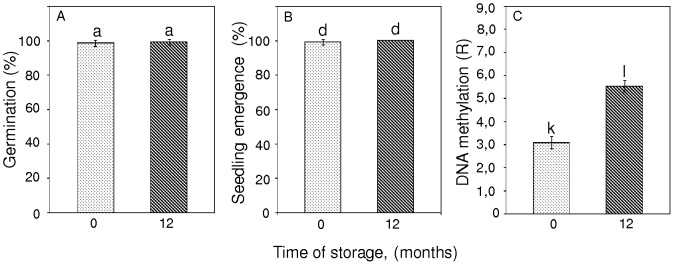
Values marked with the same letter are not significantly different at P<0.05, ANOVA, Tukey`s test, error bars indicate standard deviations.
Seedlings Growth
Three-month-old seedlings that were derived from seeds dried to 2.8% mc were significantly shorter (mean height 61.5 mm) than similar aged seedlings that were generated from control seeds (8.8% mc; mean height 66.6 mm), (Figure 6).
Figure 6. Height of seedlings (mm) derived from desiccated pear seeds.
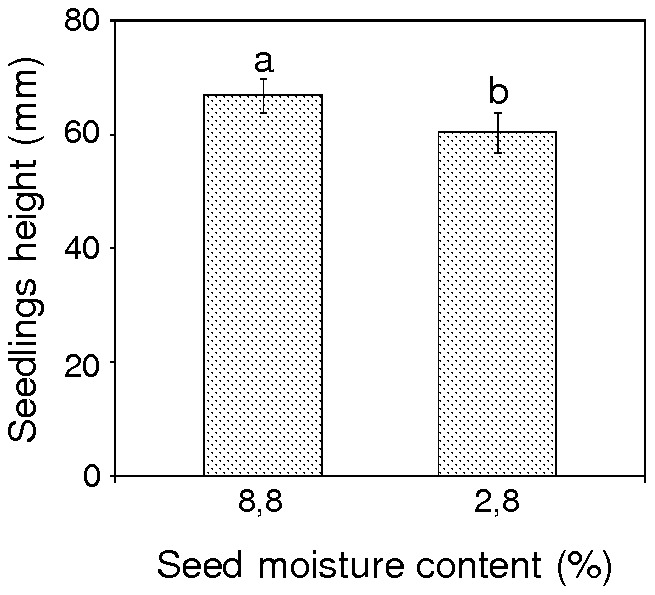
Values marked with the same letter are not significantly different at P<0.05, ANOVA, Tukey`s test. Seeds collected from Borkowice were stored for 1 year at 3°C before desiccation, error bars indicate standard deviations.
DNA Methylation in Seedlings Derived from Desiccated Seeds
The global DNA methylation level in seedlings obtained from seeds desiccated to different levels of mc (2.8–8.8%) ranged from R = 6.49 to 12.95% (Figure 7). The highest level of m5C (R = 12.95%) was observed in seedlings that were grown from seeds with 8.8% mc (stored, control seed). Seedlings that were derived from seeds desiccated to 5.3% mc were characterized by a significantly lower level of global DNA methylation (10.40%). A stronger reduction in the m5C content of genomic DNA (down-methylation) to R = 6.49% was observed in seedlings that were grown from severely desiccated seeds (2.8% mc), (Figure 7).
Figure 7. Percentage 5-methylcytosine methylation of DNA in seedlings derived from desiccated pear seeds.
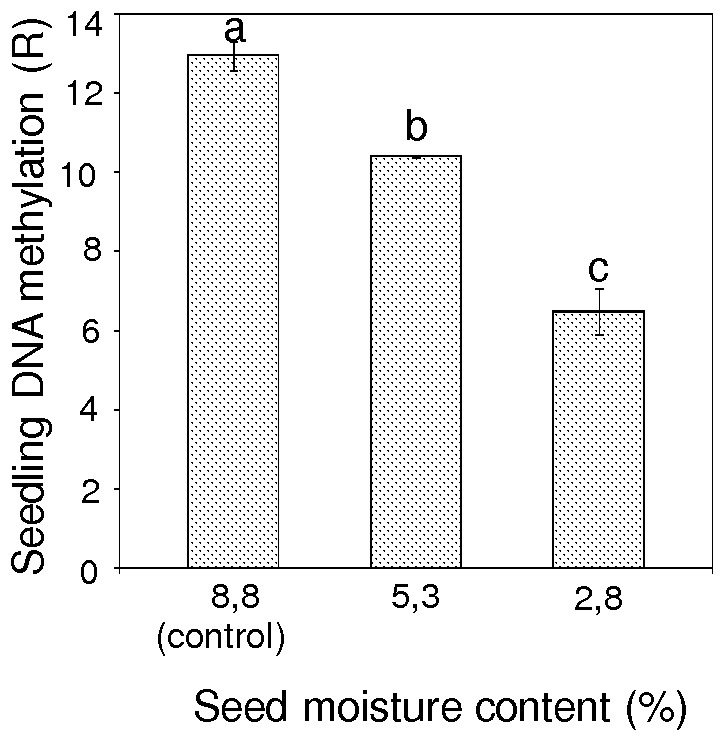
Values marked with the same letter are not significantly different at P<0.05, ANOVA, Tukey`s test. Seeds were collected from Borkowice and not stored before desiccation, error bars indicate standard deviations.
Discussion
Changes in DNA Methylation during the Desiccation of Orthodox Seeds
Cytosine methylation in DNA plays a key role in the regulation of gene expression, and therefore affects cellular protein profiles [30], [31], [32]. Such changes control the growth and development of plants [11], [33], [34], [35], because DNA methylation is not only maintained through cell division but can also be modified. The level of 5-methylcytosine ranges from 6 to 30% in plants [36] and is actively carried out by specific DNA methyltransferases [37].
The mechanism by which stresses affect the epigenome is not well understood. Increases in global DNA methylation are known to inhibit gene expression and a reduction of methylation enhances gene expression [38]. Epigenetic alterations have been identified to allow plants to adapt to new conditions without affecting the DNA sequence [39]. There are many examples of genes whose expression is regulated by methylation status. Also, genes that are responsible for the reaction to stress signals have been shown to be regulated by epigenetic mechanisms [14]. So far, it is known that DNA methylation is substantially and rapidly reduced in the coding region of a glycerophosphodiesterase-like gene one hour after oxidative stress in tobacco. The reduced DNA methylation in the coding region correlates with stress induction of the glycerophosphodiesterase-like gene [15]. In addition, drought stress can reduce the level of DNA methylation in rice and this induced epigenetic change in its genome is considered as a very important regulatory mechanism which enables rice to adapt to drought [40].
At the present time, it is not well understood if changes in global DNA methylation in seeds play an important role in the regulation of genome-wide gene expression in response to environmental stress, especially desiccation. The ability of tissues to survive desiccation is commonly found in orthodox seeds. It is thought that metabolism shuts down as plant tissues dehydrate and enter into a state of suspended animation [41]. Contrary to this statement, our results showed that genome-wide changes of DNA methylation were induced in seeds after periods of severe desiccation. This study is the first report which shows that seeds can react at an epigenetic level to stress signals. Such changes may be a result of physiological-induced processes and unspecific DNA damages that are not the effects of programmed events. Unfortunately, at the present time, we cannot link this epigenetic reaction with any specific physiological response. However, it seems to be feasible that observed alterations in DNA methylation of dried seeds induce transcriptional changes during or before seed imbibition [42]. Nevertheless, we showed that the desiccation of common pear seeds from 8.2–8.8% to 4.8–5.3% resulted in a slight decline of global DNA methylation levels. However, no changes were observed for seed germination or the emergence of seedlings.
Secondly, our unique results clearly showed that the level of global DNA methylation increased significantly in highly desiccated seeds (desiccation above silica gel to 2.2–2.8% mc) compared with that of seeds desiccated to approximately 8% mc. It is known that an increase in the DNA methylation level represses gene expression [37]. Thus, it seems reasonable to consider that seeds may attempt to survive extremely unfavorable environmental conditions by using hypermethylation of DNA to reduce all vital cellular functions to a minimum.
In the present study, we revealed an important connection between global DNA methylation changes and the adjustment of orthodox seeds to severe desiccation conditions. Other authors have shown that histone methylation and deacetylation are involved in the induction of gene expression that inhibits seed germination under unfavorable environmental conditions [43], [44], [45]. In our study, we further showed that orthodox pear seeds use different epigenetic strategies when desiccated to different levels, with little demethylation occurring in response to moderate desiccation (Figure 4C), and hypermethylation occurring in response to extreme desiccation (Figure 4). Wang and co-workers [40] reported that drought stress tends to reduce the overall level of DNA methylation in the leaves and roots of rice plants. Other reports also indicate that environmental factors tend to cause demethylation of genomic DNA [15]. An opposite trend in the global level of m5C was observed for severely desiccated pear seeds in the present study.
Storage at 3°C of pear seeds from both provenances caused an increase in their global DNA methylation. Stored and non-stored seeds responded in a similar manner after severe desiccation to 2–3% mc (hypermethylation). This finding confirms our earlier assumption that this strategy for the modification of DNA methylation is characteristic of orthodox seeds and does not depend on the initial level of DNA methylation of the stored or non-stored seeds.
Analyses of time-dependent changes in seed viability and global m5C levels confirmed that a period of 1-year storage had no effect on the germination or emergence of seedlings compared to fresh, non-stored seeds. However, a global increase in DNA methylation from 3.08 to 5.49% was observed in response to storage at 3°C (Figure 5).
Effect of Seed Desiccation on DNA Methylation of Seedlings derived from Desiccated Seeds
We showed for the first time that the level of DNA methylation is generally lower in seeds than in seedlings. Previously, it was shown that the endosperm of Arabidopsis seeds had lower levels of DNA methylation than the embryo [46], [47]. Moreover, DNA methylation was higher in mature leaves (needles) of Pinus radiata than in seedlings [48]. We demonstrated that changes in the global level of m5C in seeds are detectable in 3-month-old seedlings. These alterations that were observed in seedlings were reversed in comparison to those observed in seeds. A desiccation stress signal which had affected seeds reveal in seedlings as significant decline in amount of m5C. Seedlings derived from seeds that were dried to 2.8 and 5.3% mc contained a lower level of DNA methylation than those obtained from seeds with 8.8% mc. Moreover, severe desiccation of seeds resulted in a subsequent reduction of height in germinated seedlings.
It was previously observed that DNA methylation was lowered in two heterochromatic loci in tobacco cell suspensions that were treated with osmotic and salinity stress and this trend was reversed after cessation of the stress [49]. However, our results indicate that a desiccation stress signal affecting seeds alters DNA methylation, both in seeds and seedlings derived from these seeds, even they were not subjected to desiccation stress. The global m5C levels in pear seedlings that are older than 3 months still remain to be determined.
Conclusions
We investigated the influence of desiccation on Pyrus communis seeds and seedlings. Our comparative studies confirmed that severe desiccation did not negatively impact seed germination rates, slightly reduced seedling emergence and resulted in a minor reduction of seedling height. However, we observed that severe desiccation of seeds caused an opposite effect in the DNA methylation of these seeds (slight increase) and in 3-month old seedlings (decrease). We concluded that orthodox seeds are capable of reacting to severe desiccation by means of epigenetic phenomena.
Acknowledgments
The authors would like to thank Elżbieta Drzewiecka-Pieniężna and Magdalena Sobczak for providing technical assistance.
Funding Statement
This work was partly supported by a grant from the National Science Centre, Poland (no. N309 072036) and General Directorate of the State Forests in Warsaw (OR-2717-2/12). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Zhou J, Wang X, Jiao Y, Qin Y, Liu X, et al. (2007) Global genome expression analysis of rice in response to drought and high-salinity in shoot, flag leaf, and panicle. Plant Mol Biol 63: 591–608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Kalemba EM, Pukacka S (2008) Changes in late embryogenesis abundant proteins and a small heat shock protein during storage of beech (Fagus sylvatica L.) seeds. Environ Exp Bot 63: 274–280. [Google Scholar]
- 3. Berjak P (2006) Unifying perspectives of some mechanisms basic to desiccation tolerance across life forms. Seed Sci Res 16: 1–15. [Google Scholar]
- 4. Pukacka S, Ratajczak E, Kalemba E (2011) The protective role of selenium in recalcitrant Acer saccharium L. seeds subjected to desiccation. J Plant Physiol 168: 220–225. [DOI] [PubMed] [Google Scholar]
- 5. McDonald MB (1999) Seed deterioration physiology, repair and assessment. Seed Sci Technol 27: 177–237. [Google Scholar]
- 6. Pukacka S, Ratajczak E (2007) Ascorbate and glutathione metabolism during development and desiccation of orthodox and recalcitrant seeds of the genus Acer . Funct Plant Biol 34: 601–613. [DOI] [PubMed] [Google Scholar]
- 7.Kermode AR, Finch-Savage BE (2002) Desiccation sensitivity in orthodox and recalcitrant seeds in relation to development. In: Black M, Pritchard HW, editors Desiccation and Survival in Plants, Drying Without Dying. CAB International, Wallingford. 149–184.
- 8.Vertucci CW, Farrant JM (1995) Acquisition and loss of desiccation tolerance. In: Kigel J, Galili G, editors. Seed Development and Germination. Marcel Dekker, New York. 237–271.
- 9. Pammenter NW, Berjak P (1999) A review of recalcitrant seed physiology in relation to desiccation-tolerance mechanism. Seed Sci Res 9: 3–37. [Google Scholar]
- 10. Berjak P, Pammenter NW (2008) From Avicennia to Zizania: Seed recalcitrance in perspective. Ann Bot-London 101: 213–228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Law JA, Jacobsen SE (2009) Dynamic DNA methylation. Science 323: 1568–1569. [DOI] [PubMed] [Google Scholar]
- 12. Chan SW, Henderson IR, Jacobsen SE (2005) Gardening the genome: DNA methylation in Arabidopsis thaliana . Nat Rev Genet 6: 351–360. [DOI] [PubMed] [Google Scholar]
- 13. Steward N, Ito M, Yamaguchi Y, Koizumi N, Sano H (2002) Periodic DNA methylation in maize nucleosomes and demethylation by environmental stress. J Biol Chem 277: 37741–37746. [DOI] [PubMed] [Google Scholar]
- 14. Boyko A, Kathiria P, Zemp FJ, Yao Y, Pogribny I, et al. (2007) Transgenerational changes in the genome stability and methylation in pathogen-infected plants. Nucleic Acids Res 35: 1714–1725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Choi CS, Sano H (2007) Abiotic-stress induces demethylation and transcriptional activation of gene encoding a glycerophosphodiesterase-like protein. Mol Genet Genomics 277: 589–600. [DOI] [PubMed] [Google Scholar]
- 16. Moliner J, Ries G, Zipfel C, Hohn B (2006) Transgeneration memory of stress plants. Nature 442: 1046–1049. [DOI] [PubMed] [Google Scholar]
- 17. Verhoeven KJ, Jansen JJ, van Dijk PJ, Bire A (2010) Stress-induced DNA methylation changes and their heritability in asexual dandelions. New Phytol 185: 1108–1118. [DOI] [PubMed] [Google Scholar]
- 18. Finnegan E, Dennis E (1993) Isolation and identification by sequence homology of putative cytosine methyltransferase from Arabidopsis thaliana . Nucleic Acids Res 21: 2383–2388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Kankel MW, Ramsey DE, Stokes TL, Flowers SK, Haag JR, et al. (2003) Arabidopsis MET1 cytosine methyltransferase mutants. Genetics 163: 1109–1122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Saze H, Scheid O, Paszkowski J (2003) Maintenance of CpG methylation is essential for epigenetic inheritance during plant gametogenesis. Nat Genet 34: 65–69. [DOI] [PubMed] [Google Scholar]
- 21. Jackson J, Lindroth A, Cao X, Jacobsen S (2002) Control pf CpNpG DNA methylation by the KRYPTONITE histone H3 methyltransferase. Nature 416: 556–560. [DOI] [PubMed] [Google Scholar]
- 22. Cao X, Jacobsen SE (2002) Role of Arabidopsis DRM methyltransferases in de novo DNA methylation and gene silencing. Curr Biol 12: 1138–1144. [DOI] [PubMed] [Google Scholar]
- 23. Chan SW, Zilberman D, Xie Z, Johansen LK, Carrington JC, et al. (2004) RNA Silencing Genes Control de Novo DNA Methylation. Science 303: 1336. [DOI] [PubMed] [Google Scholar]
- 24. Harding K (2004) Genetic integrity of cryopreserved plant cells: a review. CryoLetters 25: 3–22. [PubMed] [Google Scholar]
- 25. Chinnusamy V, Zhu JK (2009) Epigenetic regulation of stress responses in plants. Curr Opin Plant Biol 12: 133–139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Suszka B, Muller C, Bonnet-Masimbert M (1996) Seeds of forest broadleaves, from harvest to sowing. Paris: Institut National de la Recherche Agronomique, 294 p. [Google Scholar]
- 27.Gordon AG, Rowe DCF (1982) Seed Manual for Ornamental Trees and Shrubs. Forestry Commission Bulletin 59. London: Her Majesty’s Stationery Office.
- 28. Barciszewska MZ, Barciszewska AM, Rattan SIS (2007) TLC-based detection of methylated cytosine: application to aging epigenetics. Biogerontology 8: 673–678. [DOI] [PubMed] [Google Scholar]
- 29. Guzy-Wróblewska J, Filek M, Kaliciak A, Szarejko I, Machackova I, et al. (2013) Vernalization and photoperiod-related changes in the DNA methylation state in winter and spring rapeseed. Acta Physiol Plant 35: 817–827. [Google Scholar]
- 30. Bird A (2002) DNA methylation. Genes Dev 16: 6–21. [DOI] [PubMed] [Google Scholar]
- 31. Morgan HD, Santos F, Green K, Wendy D, Wolf R (2005) Epigenetic reprogramming in mammals. Hum Mol Genet 14: 47–58. [DOI] [PubMed] [Google Scholar]
- 32. Costa S, Shaw P (2007) “Open minded” cells: how cells can change fate. Trends Cell Biol 173: 101–106. [DOI] [PubMed] [Google Scholar]
- 33. Lister R, O’Malley RC, Tonti-Filippini J, Gregory BD, Berry CC, et al. (2008) Highly integrated single-base resolution maps of the epigenome in Arabidopsis . Cell 133: 523–536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Zhang X (2008) The epigenetic landscape of plants. Science 320: 489–492. [DOI] [PubMed] [Google Scholar]
- 35. Zhang M, Kimatu JN, Xu K, Liu B (2010) DNA cytosine methylation in plant development. J Genet Genomics 36: 1–12. [DOI] [PubMed] [Google Scholar]
- 36. Chen T, Li E (2004) Structure and function of eukaryotic DNA methyltransferases. Curr Top Dev Biol 60: 55–89. [DOI] [PubMed] [Google Scholar]
- 37. Law JA, Jacobsen SE (2010) Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat Rev Genet 11: 204–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Zemach A, McDaniel IE, Silva P, Zilberman D (2010) Genome-wide evolutionary analysis of eukaryotic DNA methylation. Science 328: 916–919. [DOI] [PubMed] [Google Scholar]
- 39. Causevic A, Delaunay A, Ounnar S, Righezza M, Delmotte F, et al. (2005) DNA methylating and demethylating treatments modify phenotype and cell wall differentiation state in sugarbeet cell lines. Plant Physiol Bioch 43: 681–691. [DOI] [PubMed] [Google Scholar]
- 40. Wang WS, Pan YJ, Zhao XQ, Dwivedi D, Zhu LH, et al. (2011) Drought-induced site-specific DNA methylation and its association with drought tolerance in rice (Oryza sativa L.). J Exp Bot 66: 1951–1960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Vicre M, Farrant JM, Driouich A (2004) Insights into the cellular mechanisms of desiccation tolerance among angiosperm resurrection plant species. Plant Cell Environ 27: 1329–1340. [Google Scholar]
- 42. Holdsworth MJ, Bentsink L, Soppe WJ (2008) Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytol 179: 33–54. [DOI] [PubMed] [Google Scholar]
- 43. Henderson JT, Li HC, Rider SD, Mordhorst AP, Romero-Severson J, et al. (2004) PICKLE acts throughout the plant to repress expression of embryonic traits and may play a role in gibberellins dependent responses. Plant Physiol 134: 995–1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Li HC, Chuang K, Henderson JT, Rider SD Jr, Bai Y, et al. (2005) PICKLE acts during germination to repress expression of embryonic traits. Plant J 44: 1010–1022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Chinnusamy V, Gong Z, Zhu JK (2008) Abscisic acid-mediated epigenetic processes in plant development and stress responses. J Integr Plant Biol 10: 1187–1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Gehring M, Bubb KL, Henikoff S (2009) Extensive demethylation of repetitive elements during seed development underlines gene imprinting. Science 324: 1447–1451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Hsieh TF, Ibarra CI, Silva P, Zemach A, Eshed-Williams L, et al. (2009) Genome-wide demethylation of Arabidopsis endosperm. Science 324: 1451–1454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Fraga MF, Jesús Cañal M, Rodriguez R (2002) Phase-change related epigenetic and physiological changes in Pinus radiata D. Don. Planta 215: 672–678. [DOI] [PubMed] [Google Scholar]
- 49. Kovarik A, Koukalová B, Bezdek M, Opatrn Z (1997) Hypermethylation of tobacco heterochromatic loci in response to osmotic stress. Theor Appl Genet 95: 301–306. [Google Scholar]