

Tropomyosin is in many ways the archetypal coiled-coil fibrous protein and has played a central role in developing an understanding of the function of this class of proteins in structural terms. Analysis of its sequence (1) provided insight into both the general principles underlying coiled-coil formation as well as identifying interaction interfaces between tropomyosin and other proteins (2–8). Structural studies on tropomyosin have been frustrated by its flexibility and the high solvent content of crystals and, although several low- and intermediate-resolution structures have been obtained (9–11), these were only able to identify general outlines, such as coiled-coil pitch. Determination of a 2-Å-resolution crystal structure of a 81-residue fragment comprising almost a third of tropomyosin (12) represents the culmination of over 30 years' determined effort by Cohen's laboratory and verifies many sequence analysis predictions at atomic resolution, as well as giving insight into how tropomyosin might bend around actin in muscle thin filaments.
Tropomyosin is a central component of the regulatory machinery of skeletal muscle contraction and is constructed from two α-helical chains that wrap around one another to form a coiled-coil rod, approximately 20 Å in diameter and 400 Å long, that lies in the grooves of the actin helix in thin filaments. The widely accepted steric blocking mechanism of vertebrate skeletal muscle regulation (13, 14) proposes that changes in the structure of the troponin complex induced by changes in free Ca2+ concentration are transmitted to tropomyosin and cause it to change position in the actin groove. The simplest formulation of this model proposes that in high Ca2+ (“on” position), tropomyosin is located deep in the actin groove, allowing myosin to bind to actin and generate force, whereas in low Ca2+ (“off” position), tropomyosin is located at higher radius and obstructs the myosin-actin interaction. Although tropomyosin movement also may introduce structural changes in actin and there also may be three states rather than two (15), there is persuasive evidence for the tropomyosin movement itself (16, 17).
Crick (18) identified the key structural basis for the way the two helices in a coiled-coil interact. Each α-helix contains seven residues (designated a, b, c, d, e, f, and g) in approximately two turns. At the interface between the helices, the side chains of residues in core positions a and d that protrude from one α-helix pack into the spaces between the a and d side chains in the other (Fig. 1). Crick called this “knobs in holes” packing and noted that small changes in the number of residues per turn in the α-helix (which need not be integral) would cause the helices to coil around one another (reviewed in ref. 19). He also proposed that the a and d residues would be mainly hydrophobic and packing them in the core would make a major contribution to the formation of the coiled-coil. Coiled-coils are a widespread motif in proteins where they often generate dimers as well as having structural roles (reviewed in refs. 20 and 21).
Figure 1.

Interaction between the two α-helices in the tropomyosin coiled-coil. Each α-helix is shown with seven residues (a–g) in two turns. (A) End-on view looking from N terminus. The interface between the α-helices derives primarily from hydrophobic residues in core positions a and d, although there are also some salt bridges formed between residues e and g. (B) The core interface viewed parallel to the coiled-coil axis shows how residues from one chain occupy the spaces between the corresponding residues from the second chain to give “knobs in holes” packing (18). In tropomyosin there are slightly more than 3.6 residues per turn, which produces a left-handed supercoil.
Because of the structural regularity of tropomyosin, a great deal has been deduced from its amino acid sequence (1). In addition to a 7-fold gene duplication repeat (4), there is a characteristic pattern of hydrophobic residues in positions a and d (1–3) consistent with the knobs in holes hypothesis. Model building indicated that salt bridges between positions e and g were also important and favored the chains being in register (3). The high-resolution structure of the tropomyosin fragment (12) shows both the knobs in holes packing and the e-g salt bridges and in this respect is similar to the structures of other coiled-coils such as the GCN4 leucine zipper (22) and cortexillin (23). There is also a significant variation of radius of coiled-coil associated with different sizes of core side chains.
In thin filaments, each tropomyosin binds to seven actins along the groove of the actin helix. Fourier analysis (5–7) detected 14 quasi-equivalent negatively charged zones associated with actin binding. Seven zones are thought to correspond to binding for the on position and the other seven to the off position and, on the basis of subtle differences in sequence, were classified into α- and β-bands, respectively (7). There was also a pattern of hydrophobic residues on the exterior of the coiled-coil (in positions b, c, and f), which may be associated with a third position of the tropomyosin relative to actin (24). Irregularities in the 14-fold repeat pattern (8) together with electron microscopy data (25) indicated residues possibly involved in binding to troponin. Although most tropomyosin sequence repeats are based on 14 zones, there is a weaker 7-fold periodicity associated with small core resides (5). However, this repeat is not as regular as the zones of negative charge, and so is unlikely to be involved directly in actin binding.
Tropomyosin has to bend to wrap around the actin helix and this bending can produce two types of local coiled-coil deformation (Fig. 2). A bend about the narrow edge of the coiled-coil simply bends both α-helices in the same way, but a bend about the broad face involves a shearing motion so that one chain will move relative to the other. Theoretical analysis indicated that both types of distortion were likely to be present and also proposed that the second type, involving a relative sliding of the chains, might be associated with the 7-fold periodicity in the distribution of small amino acids in the coiled-coil core (5). Brown et al. (12) observe that there is indeed an axial sliding of the two chains in the coiled-coil where there are clusters of alanine in position d. There are seven of these alanine clusters located near the C terminus of each gene-duplication repeat. There will be a larger hole between the small side chains of alanines in the core in this region while the knob that fits into it will be correspondingly smaller, and this enables one chain to move a small distance (of the order of 1 Å) relative to the other (12).
Figure 2.
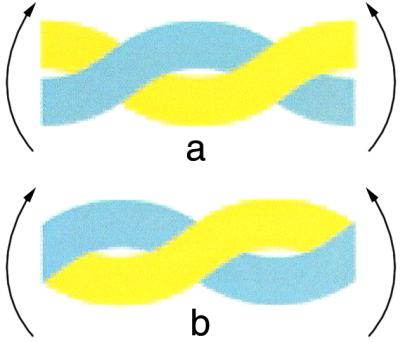
Bending coiled-coils about an axis perpendicular to the plane of the page. Bending about the broad face (a) involves a shearing motion so that one chain (yellow) follows a longer path and so the two chains need to slide relative to one another elsewhere in the coiled-coil to compensate. However, a bend about the narrow edge of the supercoil (b) simply bends both α-helices in the same way. Both types of bending are likely to be present when tropomyosin wraps around actin in muscle thin filaments.
Although tropomyosin generally exhibits an undulating conformation in crystals and also in many thin filament reconstructions, this bending probably derives from crystal contacts rather than indicating an intrinsically undulating shape for the molecule itself, consistent with the core alanine clusters helping to accommodate bending stress rather than themselves introducing distortions. Although other coiled-coil proteins have analogous clusters of core alanines, tropomyosin appears to have a much higher concentration (12) and many coiled-coil proteins, such as myosin, paramyosin, and intermediate filament proteins, instead introduce kinks using skip residues (26). One explanation for the large number of core alanine clusters in tropomyosin might be that the molecule needs to be more flexible to accommodate changes in its position in thin filaments. It has been proposed, for example, that tropomyosin may rotate by a quarter turn when it moves between on and off positions (5, 6). Such a rotation would interchange broad and narrow faces of the supercoil and so require a redistribution of bending stresses, which would naturally be facilitated by small relative movements of the two α-helices. Consistent with this hypothesis, comparison of tropomyosin sequences from different species indicates that the conservation of core alanine clusters is greater in those that bind troponin.
Although it is still necessary to establish precisely how tropomyosin binds to actin as well as the basis of the end-to-end overlap between successive molecules, the structure of the 81-residue fragment provides a context for understanding its function in regulating muscle contraction at the atomic level.
Footnotes
See companion article on page 8496.
References
- 1.Stone D, Sodek J, Johnson P, Smillie L B. Proc IX FEBS Meeting. 1975;31:125–136. [Google Scholar]
- 2.McLachlan A D, Stewart M. J Mol Biol. 1975;98:293–304. doi: 10.1016/s0022-2836(75)80119-7. [DOI] [PubMed] [Google Scholar]
- 3.Parry D A D. J Mol Biol. 1975;98:519–535. doi: 10.1016/s0022-2836(75)80084-2. [DOI] [PubMed] [Google Scholar]
- 4.McLachlan A D, Stewart M, Smillie L B. J Mol Biol. 1975;98:281–291. doi: 10.1016/s0022-2836(75)80118-5. [DOI] [PubMed] [Google Scholar]
- 5.McLachlan A D, Stewart M. J Mol Biol. 1976;103:271–298. doi: 10.1016/0022-2836(76)90313-2. [DOI] [PubMed] [Google Scholar]
- 6.Parry D A D. Biochem Biophys Res Commun. 1976;68:323–328. doi: 10.1016/0006-291x(76)91146-3. [DOI] [PubMed] [Google Scholar]
- 7.Stewart M, McLachlan A D. Nature (London) 1975;257:331–333. doi: 10.1038/257331a0. [DOI] [PubMed] [Google Scholar]
- 8.McLachlan A D, Stewart M. J Mol Biol. 1976;106:1017–1022. doi: 10.1016/0022-2836(76)90349-1. [DOI] [PubMed] [Google Scholar]
- 9.Phillips G N, Fillers J P, Cohen C. J Mol Biol. 1986;192:111–131. doi: 10.1016/0022-2836(86)90468-7. [DOI] [PubMed] [Google Scholar]
- 10.Whitby F G, Kent H, Stewart F, Stewart M, Xie X, Hatch V, Cohen C, Phillips G N. J Mol Biol. 1992;227:441–452. doi: 10.1016/0022-2836(92)90899-u. [DOI] [PubMed] [Google Scholar]
- 11.Whitby F G, Philips G N. Proteins Struct Funct Genet. 2000;38:49–59. [PubMed] [Google Scholar]
- 12.Brown J H, Kim K-H, Jun G, Greenfield N J, Dominguez R, Volkmann N, Hitchcock-DeGregori S E, Cohen C. Proc Natl Acad Sci USA. 2001;98:8496–8501. doi: 10.1073/pnas.131219198. . (First Published July 3, 2001; 10.1073/pnas.131219198) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Huxley H E. Cold Spring Harbor Symp Quant Biol. 1972;37:361–376. [Google Scholar]
- 14.Parry D A D, Squire J M. J Mol Biol. 1973;75:33–55. doi: 10.1016/0022-2836(73)90527-5. [DOI] [PubMed] [Google Scholar]
- 15.Vibert P, Craig R, Lehman W. J Mol Biol. 1997;266:8–14. doi: 10.1006/jmbi.1996.0800. [DOI] [PubMed] [Google Scholar]
- 16.Xu C, Craig R, Tobacman L, Horowitz R, Lehman W. Biophys J. 1999;77:985–992. doi: 10.1016/S0006-3495(99)76949-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Narita A, Yasunaga T, Ishikawa T, Mayanagi K, Wakabayashi T. J Mol Biol. 2001;308:241–261. doi: 10.1006/jmbi.2001.4598. [DOI] [PubMed] [Google Scholar]
- 18.Crick F H C. Acta Crystallogr. 1953;6:689–697. [Google Scholar]
- 19.Fraser R D B, Macrae T P. Conformation in Fibrous Proteins and Related Synthetic Polypeptides. New York: Academic; 1973. [Google Scholar]
- 20.Cohen C, Parry D A D. Proteins Struct Funct Genet. 2000;7:1–15. [Google Scholar]
- 21.Burkhard P, Strelkov S V, Stetefeld J. Trends Cell Biol. 2001;11:82–88. doi: 10.1016/s0962-8924(00)01898-5. [DOI] [PubMed] [Google Scholar]
- 22.O'Shea E K, Klemm J D, Kim P S, Alber T. Science. 1991;254:539–544. doi: 10.1126/science.1948029. [DOI] [PubMed] [Google Scholar]
- 23.Burkhard P, Kammerer R A, Steinmetz M O, Bourkenkov G P, Aebi U. Structure (London) 2000;8:223–230. doi: 10.1016/s0969-2126(00)00100-3. [DOI] [PubMed] [Google Scholar]
- 24.Matsuura Y, Stewart M, Kawamoto M, Kamiya N, Saeki K, Yasunaga T, Wakabayashi T. J Mol Biol. 2000;296:579–595. doi: 10.1006/jmbi.1999.3467. [DOI] [PubMed] [Google Scholar]
- 25.Stewart M. Proc R Soc London B. 1975;190:257–266. doi: 10.1098/rspb.1975.0091. [DOI] [PubMed] [Google Scholar]
- 26.Brown J H, Cohen C, Parry D A. Proteins Struct Funct Genet. 1996;26:134–145. doi: 10.1002/(SICI)1097-0134(199610)26:2<134::AID-PROT3>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]