

Abstract
Drugs that selectively activate estrogen receptor β (ERβ) are potentially safer than the nonselective estrogens currently used in hormonal replacement treatments that activate both ERβ and ERα. The selective ERβ agonist AC-186 was evaluated in a rat model of Parkinson’s disease induced through bilateral 6-hydroxydopamine lesions of the substantia nigra. In this model, AC-186 prevented motor, cognitive, and sensorimotor gating deficits and mitigated the loss of dopamine neurons in the substantia nigra, in males, but not in females. Furthermore, in male rats, 17β-estradiol, which activates ERβ and ERα with equal potency, did not show the same neuroprotective benefits as AC-186. Hence, in addition to a beneficial safety profile for use in both males and females, a selective ERβ agonist has a differentiated pharmacological profile compared to 17β-estradiol in males.
Keywords: Parkinson’s disease, neuroprotection, AC-186, gender difference, buccal/sublingual administration, selective estrogen receptor beta agonist
Subtype selective estrogen receptor modulation may have a role in the treatment of neurodegenerative disorders, including Parkinson’s disease (PD), Alzheimer’s disease (AD), and multiple sclerosis (MS).1−4 In most of these diseases, aging is the major risk factor, and they generally occur around the age when endogenous sex steroid levels in plasma and brain are declining.5 Mechanistically, the protective effects of estrogens have been linked to stabilization of mitochondria by protection against oxidative stress.6 In neurons, estrogen can prevent the initiation of programmed cell death by mitochondria via regulation of several members of the B-cell lymphoma 2 (Bcl-2) family, including enhancing expression of antiapoptotic Bcl-xL and Bcl-w genes and down-regulating expression of the pro-apoptotic gene Bim.7,8 Alternatively, estrogens may inhibit apoptosis by activation of the antioxidant defense system, Nrf2-Keap1.9 In addition, estrogens may regulate mitochondrial bioenergetic capacity by increasing expression of mitochondrial complex 1 and cytochrome c oxidase.10 More intriguingly, estrogens regulate enzymes that catabolize proteins that aggregate in AD and PD, that is, neprilysin, insulin-degrading enzymes, and cathepsin D, which degrade the Aβ-peptide and α-synuclein.11−13 Furthermore, estrogen signaling plays an essential role in modulating neuroplasticity and cognition.14 The therapeutic use of the endogenous estrogen 17β-estradiol (E2), which activates both ERα and ERβ equally, is limited due to its feminizing effects and an increased risk of cancer. However, these side effects are believed to be associated with activation of the ERα.15,16 Thus, ERβ selective compounds could potentially be safer drugs for chronic treatment in both men and women.
Most studies with estrogens have been conducted in women or using female animals.4 For example, a study of a selective ERβ agonist, ERB-041, in 291 patients with rheumatoid arthritis (RA) showed no significant effect compared with placebo; however, over 93% of patients in the placebo group and around 80% in the treatment groups were women.17 Nonetheless, there are indications that point to estrogens and in particular ERβ agonists as potential therapies in males via the selective ERβ agonist 5α-androstane-3β,17β-diol, a metabolite of dihydrotestosterone (DHT).16,18,19 Hence, in men, the pronounced age dependent decrease of androgen levels seen in the brain of patients with neurodegenerative diseases compared with nondemented patients5 could affect ERβ activation. Recently we found that chronic activation of ERβ using AC-186 in 3xTg-AD mice modulated the expression of androgen receptor levels.20 Further, looking at gender differences in gene expression and vulnerability to rotenone in ventral mesencaphalic cultures of dopamine neurons, ERβ was significantly differentially expressed in males versus females.21
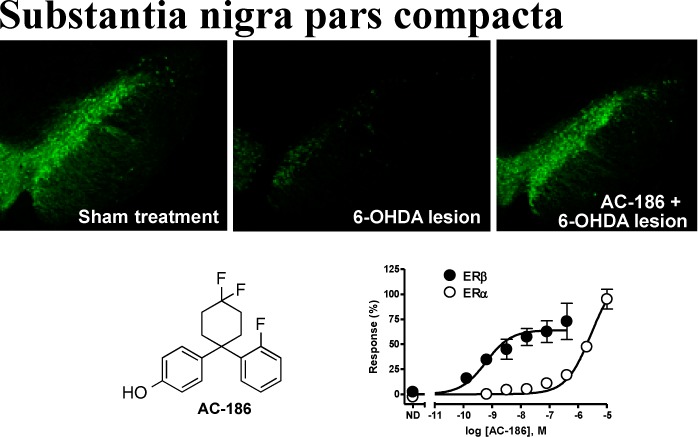
In 2008, we reported that a nonsteroidal selective ERβ agonist, AC-131 (Figure 1), was active in several pain animal models involving nerve injury and chronic inflammatory pain.16,23 From a lead discovery program, three diverse compounds from this class that demonstrated stability in human liver microsomes, AC-186, AC-623, and AC-957, were further evaluated as potential drug development candidates. Because oral dosing can be difficult in elderly patients that experience difficulty swallowing, we considered alternative routes to oral administration. In addition to ease of use, buccal administration offers certain advantages over oral administration, namely, rapid drug delivery into the bloodstream, and avoidance of first pass metabolism, resulting in a lower overall exposure burden, for example, liver exposure. The latter is clearly exemplified in hormone replacement therapy using patches which have a better safety profile compared to oral administration.24 The three selective ERβ agonists were assessed for their permeability across human buccal membranes. Based on the apparent permeability coefficients (Papp) of the test compounds assessed in MatTek EpiOral buccal tissue, although the structural similarity, AC-186 (12.5 × 10–6cm/s) was classified as a highly permeable compound, while AC-623 (3.1 × 10–6 cm/s) and AC-957 (3.9 × 10–6 cm/s) were not. The high permeability of AC-186 was confirmed in Caco-2 cells, where it had apical-basolateral Papp = 27.9 × 10–6 cm/s and basolateral-apical Papp = 40.4 × 10–6 cm/s, for an efflux ratio of 1.45. An efflux ratio < 2 indicates that transport of AC-186 across membranes is not subject to efflux proteins. AC-186 was therefore selected as a candidate for further PK studies via buccal/sublingual administration. In rats, AC-186 demonstrated around 8% oral bioavailability but 79% bioavailability following sublingual administration; the latter correlates with the exposures when administered s.c. in rats. Therefore, s.c. administrations were used in the in vivo experiments. The oral cavity of dogs is considered a more accurate model for predicting buccal absorption in humans because of the histological similarities between canine and human buccal mucosa,25 and in a follow-up study in beagle dogs AC-186 showed 44% bioavailability with buccal administration using a standard formulation vehicle, polyethylene glycol.
Figure 1.

Structures and EC50 values in a functional assays (Receptor Selection and Amplification Technology, R-SAT)22 for ERα and ERβ of 17β-estradiol, AC-131, AC-186, AC-623, and AC-957.
To assess in vivo neuroprotective efficacy, the compounds were evaluated for neuroprotective efficacy in rats receiving bilateral 6-OHDA lesions of the substantia nigra pars compacta (SNpc). Prior to surgeries, male rats received baseline assessment of weight, motor function (spontaneous locomotor behavior, rotorod performance), and sensorimotor gating (prepulse inhibition of acoustic response). Subjects were assigned to treatment conditions to equalize performance on all these measures. Subjects (n = 9–15) were then treated once daily for 7 days with either vehicle (sesame oil), estradiol (100 μg/kg sc), AC-131 (10 mg/kg sc), AC-186 (10 mg/kg sc), AC-623 (10 mg/kg sc), or AC-957 (10 mg/kg sc). On the second day of treatment, subjects received bilateral sham or 6-hydroxydopamine (6-OHDA) lesions (8 μg in 4 μL) of the SNpc (A/P −5.0 mm, M/L ±1.6 mm, D/V −8.2 mm) following the rat stereotaxic atlas of Paxinos and Watson (1998).26 Behavioral assessments of motor function (spontaneous locomotion, rotorod, and traversing a challenging beam), cognition (novel object recognition), and sensorimotor gating (prepulse inhibition) began after the final day of treatment. Following the behavioral assessments, tissue sections at the levels of SNpc and striatum (STR) were collected in order to evaluate the survival of nigro-striatal dopamine neurons.
In all cases, sham animals treated with vehicle, estradiol, AC-131, AC-186, AC-623, or AC-957 displayed no behavioral differences. Thus, for clarity, data from sham animals of all treatment conditions were pooled. Relative to sham-lesioned controls, lesioned animals (6-OHDA/vehicle) displayed impaired motor performance (Figure 2) as measured by spontaneous locomotor activity, the ability to remain on a rotorod that gradually increased speed of rotation, or the ability to traverse a challenging beam. Lesioned animals also showed impaired cognition (Figure 3) and sensorimotor gating (Figure 4), deficits that are commonly observed in Parkinson’s patients. Treatment with AC-131, AC-186, AC-623, or AC-957 beginning 1 day prior to 6-OHDA infusion prevented the expression of all of these behavioral deficits, with the exception that AC-957 did not prevent the cognitive deficit associated with SNpc lesions. Notably, treatment with estradiol prevented deficits in prepulse inhibition, but did not reliably improve any additional motor or cognitive performance measures. Based on the ADMET and initial in vivo data, AC-186 was selected as the lead molecule for further development, AC-186 together with our previous published tool compound, AC-131, were compared with estradiol for their ability to protect nigrostriatal dopamine neurons as measured by tyrosine hydroxylase (TH) immunohistochemistry in the SNpc and STR. 6-OHDA lesions in vehicle treated rats produced decreases in all TH immunofluorescence measures including cell counts, cell size, mean pixel intensity in the field of view, and percent of field immunopositive compared to vehicle treated sham-lesioned rats (Figure 5). Consistent with an improved behavioral profile, animals that received AC-131 or AC-186 had normal levels of tyrosine hydroxylase immunofluorescence (TH+) in the SNpc (Figure 5) and STR (Figure 6) compared to vehicle-treated 6-OHDA-lesioned animals. Estradiol, AC-131, or AC-186 treatment of 6-OHDA-lesioned animals resulted in a larger number of TH+ cells in the SN compared with vehicle-treated 6-OHDA animals. Further, in 6-OHDA-lesioned animals treated with the selective ERβ agonists AC-131 or AC-186, but not estradiol, the size of TH+ cells (pixels/cell), the percent of the image that was immunopositive (probably reflecting both cell size and the presence of large, visible fibers), and the intensity of immunofluorescence (reflecting TH density) was improved relative to vehicle-treated 6-OHDA-lesioned animals. Similarly, selective activation of ERβ receptors with AC-131 or AC-186, but not nonselective activation with estradiol, was able to protect dopaminergic terminals in the STR as reflected by percent of the image that was TH+ and the intensity of TH immunofluorescence. Thus, the selective agonists AC-131 and AC-186 were better able to protect the viability or functionality of dopamine cells in the SN and STR compared with estradiol, and this was reflected in a normalized behavioral profile following exposure to 6-OHDA. Additional studies showed that AC-186 conducted in ovariectomized female rats following the same protocols used for motor performance and tyrosine hydroxylase analyses in the male rats, did not provide any evidence of behavioral normalization or neuroprotection (see Figures S1 and S2 in the Supporting Information).
Figure 2.

Motor performance evaluations of sham (all treatments combined) and 6-hydroxydopamine-lesioned animals treated with sesame oil vehicle (Veh), estradiol (E2), AC-131, AC-186, AC-623, or AC-957. (A) Distance traveled during a 15 min spontaneous locomotor session. (B) Time animals remained on the rotorod on the test trials. (C and D) Start latency and time required to traverse the challenging beam, respectively. For each of these measures of motoric ability, 6-OHDA-lesions impaired performance (*p < 0.05 vs sham-lesioned animals), and treatment with an ERβ agonist (AC-131, AC-186, AC-623, or AC-957) prevented the impairments (+p < 0.05 vs vehicle-treated 6-OHDA-lesioned animals). The data for AC-623 in spontaneous locomotor activity are not shown because the behavioral measure did not QC. Data were analyzed using one-way ANOVA, followed by Bonferroni’s multiple comparison post hoc analyses. Data are presented as the group means ± standard error of the mean (SEM), and represent 9–15 animals per group.
Figure 3.

Novel object recognition (NOR) task performance of sham (Sham-All Tx) and 6-OHDA (Lesion) animals. Sham-treated animals spent more time exploring the novel versus familiar object (>50%), indicating a normal preference for the novel object during the test condition. In contrast, vehicle-treated 6-OHDA animals did display a preference for either the novel or familiar object (=50%), and their performance was significantly different from sham-lesioned control animals (*p < 0.05). 6-OHDA animals treated with AC-131, AC-186, or AC-623, but not AC-957 or estradiol, showed normalized preference for the novel object compared to lesioned controls (+p < 0.05). There were no statistically significant difference between lesioned animals and sham animals in exploration time during sample. Thus, treatment with an ERβ agonist protected against 6-OHDA lesion-induced deficits in NOR cognitive performance. Data were analyzed using one-way ANOVA followed by Bonferonni’s multiple comparison post hoc analyses.
Figure 4.

Prepulse inhibition (PPI) performance in sham and 6-OHDA-treated animals. Sham-lesioned animals demonstrated a normal inhibition of the startle response with increased prepulse intensity (i.e., increased % prepulse inhibition). Two-way ANOVA revealed significant main effects of treatment condition and prepulse intensity. Bonferroni post hoc analyses for repeated measures showed that vehicle-6-OHDA animals showed impaired PPI at all prepulse intensities compared with sham controls (*p < 0.05). Pretreatment with estradiol, AC-131, AC-186, AC-623, or AC-957 prevented the 6-OHDA-induced deficits in PPI (+p < 0.05 vs vehicle-treated 6-OHDA lesioned animals). Data were analyzed using repeated measures two-way ANOVA followed by Bonferonni’s multiple comparison post hoc analyses.
Figure 5.

Tyrosine hydroxylase immunofluorescence (TH+) in the SN following sham- or 6-OHDA-lesion. 6-OHDA lesions produced (A) reduced TH+ cell counts in the SN, (B) reduced percentage of the image that was TH+, (C) reduced mean cell size, and (D) reduced mean pixel intensity of immunofluorescent pixels (all *p < 0.05 vs sham control subjects). Treatment with estradiol, AC-131, AC-186, AC-623, or AC-957 ameliorated 6-OHDA lesion-induced reductions in cell number (*p < 0.05 vs vehicle/6-OHDA group), while only the selective ERβ agonists AC-131, AC-186, AC-623, or AC-957 improved percent TH+, mean cell size, and mean pixel intensity (*p < 0.05 vs vehicle/6-OHDA group). Data were analyzed with one-way ANOVA followed by Bonferroni’s post hoc comparisons.
Figure 6.

Tyrosine hydroxylase immunofluorescence (TH+) in the STR following sham- or 6-OHDA-treatment. 6-OHDA lesions reduced (A) the percentage of the image that was TH+ and (B) mean pixel intensity of immunofluorescent pixels (*p < 0.05 vs sham control animals). Treatment with the ERβ-selective agonist AC-131 or AC-186 resulted in an increase in percentage of image TH+ and increased mean pixel intensity compared with lesioned controls (+p < 0.05), while estradiol treatment did not prevent 6-OHDA-induced damage in the striatum.
Recent theories suggest that death of nigrostriatal dopamine neurons in PD might be associated with inflammatory mechanisms.27 Thus, AC-186 was assessed to determine whether ERβ agonist treatment might alter the inflammatory marker profile following bilateral 6-OHDA lesions of the SNpc. ELISAs for tumor necrosis factor α (TNFα), monocyte chemotactic protein 1 (MCP-1 aka CCL2), interleukin-6 (IL-6), and interleukin-1β were conducted in peripheral blood mononuclear cells (PBMCs) and brain tissue homogenates from sham or 6-OHDA lesioned animals. Preliminary studies showed that IL-6 and IL-1β were not detectable in PBMCs and were not elevated in brain homogenates (data not shown). Both TNFα and MCP-1 were elevated in PBMCs from 6-OHDA animals (Figure 7). Thus, these two markers were chosen for further examination. In additional experiments, 6-OHDA treatment resulted in elevated levels of TNFα in both brain and PBMCs. Treatment with AC-186 prevented the increase in TNFα levels in both brain and PBMCs (Figure 7). MCP-1 was elevated in PBMCs following 6-OHDA when blood was taken on day 7 postsurgery, but not in brain homogenates taken on day 14 (following behavioral testing). The increase in MCP-1 in PBMCs was not prevented by AC-186 treatment, even though the behavioral profile was normalized.
Figure 7.

ELISA evaluations of TNFα and MCP-1 in PBMCs (A and B) or brain homogenates (C and D). 6-OHDA lesions increased TNFα in both PBMCs and brain homogenates (+p < 0.05 vs vehicle/sham) and treatment with AC-186 blocked this increase (*p < 0.05 vs vehicle/6-OHDA lesion). MCP-1 was increased by 6-OHDA in (B) PBMCs but not (D) brain (+p < 0.05 vs vehicle/sham), and the increase in PBMCs was not prevented by AC-186.
Taken together, these studies have demonstrated that ERβ selective agonists produce a number of beneficial effects in male rats with bilateral 6-OHDA lesions. Bilateral lesions were associated with motoric, cognitive, and sensorimotor gating deficits, and all of these deficits were ameliorated by treatment with the selective ERβ agonists AC-957, AC-623, AC-131, or AC-186. Although 17β-estradiol showed effect in some experiments, for example, PPI, it did not display the same full activity as the selective ERβ agonists. The partial effects of 17β-estradiol is consistent with previous reports that administration of exogenous 17β-estradiol in males has negligible or even harmful effect.28,29 The differential activity between 17β-estradiol and a selective ERβ agonist (WAY-200070) has previously been reported, where the selective ERβ agonist showed activity, but not 17β-estradiol in a model of photoaging.30 Using experimental autoimmune encephalomyelitis (EAE), a model of MS, selective ERβ agonists promotes remyelination, but not anti-inflammatory effects in female mice.3,31 Interestingly, AC-186 did not provide neuroprotection in female rats, indicating gender, hormonal influences, or both, may influence the neuroprotective actions of selective ERβ agonists.
This study has been done using a well-established toxin model of PD which may represent an inflammatory phase of the disease, prior to an extensive α-synuclein pathology.27 Hence, treatment with ERβ agonists might represent an early or prophylactic intervention in men. Interesting, a study looking at habitual intake of dietary flavonoids and risk of PD demonstrated a 40% lower risk for PD in men who regularly ingest flavonoids in their diet, while finding no significant effect of dietary flavonoids on PD risk in women.32
Based on these findings, AC-186 has been selected for further development. Buccal/sublingual administration may be advantageous for PD patients, many of whom suffer from dysphagia. We are therefore exploring the development of AC-186 for use as a buccal/sublingually administered medicine because of its high permeability, bioavailability, safety, and CNS penetration.
Methods
Subjects
The subjects for these experiments were male Sprague–Dawley rats purchased from Charles Rivers Laboratories (Hollister, CA) weighing 225–250 g upon arrival. Rats were housed in pairs in polypropylene cages within a temperature controlled vivarium maintained on a 12 h light/dark cycle (lights on 7 a.m.). For the duration of the experiments, animals received free access to food and water. All procedures were conducted in accordance with the NIH Guidelines for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee (IACUC) at ACADIA Pharmaceuticals. Animals were acclimated to vivarium conditions and handling for a minimum of 1 week prior to surgery.
Lesion Surgery
In order to protect norepinephrine terminals, each animal received an injection of desipramine (10 mg/kg) about 15 min prior to being anesthetized using isofluorane. Animals were placed into a stereotaxic apparatus and bilateral infusions of 6-OHDA (8 μg/4 μL) or 0.2% ascorbic acid vehicle were aimed at the SNpc (A/P −5.0 mm, M/L ±1.6 mm, D/V −8.2 mm relative to bregma).
Rotorod
Rotorod testing was conducted on a rotating cylinder (70 mm diameter) with knurled tread to aid in gripping. Animals were placed on the cylinder and it was set to rotate at 1 rpm for 15 s. If animals fell or jumped from the cylinder within 30 s., they were replaced and the acclimation period restarted. Once animals successfully remained on the cylinder for the acclimation period, the speed of rotation was increased 1 rpm every 15 s to a maximum of 10 rpm. The time in seconds that animals remained on the cylinder after the acclimation period and the maximum rpm achieved were recorded. A second trial was conducted after a 2 min intertrial interval using the same procedure, but the acclimation period was decreased such that animals were only required to step with all four feet before the speed of rotation was increased.
Challenging Beam Test
The challenging beam test was conducted on a 102 cm long bilevel beam made from ABS plastic. The top, narrower beam gradually tapered from 3.5 to 0.7 cm, while the bottom, wider beam gradually tapered from 5 to 1.8 cm along the length of the beam. The beam was elevated 23 cm above the table. Animals received five training trials. On the first training trial, animals were placed at the end of the beam and were required to jump into a holding tub. On successive trials, animals were placed 25, 50, 75, and 100 cm from the end of the beam and were required to traverse the beam and jump into the holding tub at the end. Following training, a single test trial was conducted where each animal was placed at the beginning of the beam and the start latency (time required to move all four feet from their starting locations) and run time (time required to traverse the beam after starting) were recorded. Animals were allowed a maximum of 300 s to traverse the beam, at which point they were removed from the beam and a run time of 300 s was recorded.
Novel Object Recognition
Novel object recognition (NOR) testing was conducted in two phases: sample and test. For sample, subjects were placed into the NOR chamber, where two identical objects were placed. Each rat was allowed to explore for 3 min, and the time spent exploring at each position was recorded. After 3 min, each rat was removed from the arena and placed back into its cage. The test phase was conducted 4 h after the sample phase. During test, one familiar object (seen during sample) and one novel object were placed into the chamber, and each rat was allowed 3 min to explore. The test sessions were recorded on video and scored by an observer blind to each subject’s treatment condition. For test data, the percent of exploration time spent at the novel object was determined.
Prepulse Inhibition (PPI)
Prepulse inhibition testing was conducted using a previously described method.33 Briefly, testing consisted of a habituation session followed 24 h later by a test session. During the habituation session animals were exposed to the background noise (65 dB) and a number of acoustic startling stimuli (120 dB) in order to assess baseline startle reactivity. For the test session, rats were weighed and injected with test compound or vehicle and returned to their home cages. After 60 min, animals were placed in a Plexiglas chamber within a sound attenuated enclosure (Kinder Scientific, Poway, CA).The test session began with a 5 min habituation to the chamber where a continuous low level white noise was presented (65 dB). The test session then proceeded with mixed trials of pulse alone, no stimulus (to assess background movement), prepulse alone, and prepulse–pulse pairings. The startle pulse was 120 dB, while prepulses were 4, 8, or 16 dB above background. Prepulses preceded the startle pulse by 100 ms. Startle reactions to these various trial stimuli were transduced and collected via a computer controlled interface. Prepulse inhibition was measured as the reduction in startle response on prepulse–pulse trials compared to pulse alone trials. The test included 40 trials and lasted about 16 min. For each treatment, the startle reactivity of animals (i.e., magnitude of responding to 120 dB pulses) was compared to vehicle treatment.
Tyrosine Hydroxylase Fluorescent Immunohistochemistry
Animals were anesthetized and perfused transcardially with PBS followed by 4% paraformaldehyde. Fixed tissue was placed in 30% sucrose overnight and then sectioned (50 μm) through the subtantia nigra and striatum. Sections were rinsed in blocking buffer (0.8 PBS, 3% normal serum, 0.1% Triton) for 1 h. The sections were incubated with rabbit anti-tyrosine hydroxylase polyclonal antibody for 2 h at room temperature, followed by a donkey anti-rabbit fluorescent secondary antibody for 1 h. Sections were rinsed, mounted, coverslipped, and then viewed with a fluorescence microscope using the FITC channel. For each subject analyzed, three consecutive images were taken bilaterally at A/P −5.2 mm according to the atlas of Paxinos and Watson (1998). Images were analyzed using ImageJ software.
Isolation of PBMCs (Peripheral Blood Mononuclear Cells) from Whole Blood
One milliliter of whole blood was collected into sodium citrate to prevent coagulation. Within 2 h, the whole blood was mixed with an equal part balanced salt solution. Diluted blood was layered onto Ficoll-Paque PLUS without mixing in a ratio of 4:3 and centrifuged at 1600 rpm for 40 min at 18 °C. The lymphocyte layer was collected, plated at 1 × 106, and cultured overnight in cell incubators.
Brain Tissue Homogenate
Rats were anesthetized and decapitated, and brains were rapidly removed. Brains were placed on ice, and the SN and VTA dissected. Dissected tissue was frozen on dry ice and stored at −80 °C until use. The tissue was then thawed in 500 μL of 1× cell lysis buffer + 1× protease/phosphotase inhibitor. Tissue was homogenized and spun at maximum speed for 20 min at 4 °C. The supernatant was collected, and protein content assayed and frozen overnight at −80 °C.
ELISA
For PBMCs, the supernatant was collected and spun down at 2000 rpm for 4 min. ELISAs for both PBMCs and brain homogenate were run on 50 μL of supernatant in duplicate using rat MCP-1 ELISA kit (Pierce cat# ERMCP1) and Rat TNFα ELISA kit (Pierce cat# ER3TNFA).
Acknowledgments
We thank Robert W. Johnson and Tammi Howard for their technical assistance.
Supporting Information Available
Brain exposure and B:P ratios, selectivity and safety evaluations, in vivo 6-OHDA data for female rats (Figures S1 and S2), and in vitro pharmacology (R-SAT). This material is available free of charge via the Internet at http://pubs.acs.org.
Author Present Address
∥ C.N.D.: EMD Serono Research and Development Institute, Billerica, MA 01821.
Author Contributions
K.M. conceived, designed, managed, performed the in vivo experiments, and analyzed data; D.P. conceived, designed, performed the PPI in vivo experiments, and analyzed data; C.N.D. performed ELISA experiments; J.-N.M. performed in vitro experiments; D.W.B. oversaw the behavioral in vivo experiments; E.S.B. conceived and designed in vitro and in vivo experiments; R.O. conceived the initiation and managed the project, conceived in vitro and in vivo experiments, led the medicinal chemistry effort, analyzed data, and wrote the paper.
This work was supported by grants from the Michael J. Fox Foundation for Parkinson’s research.
The authors declare the following competing financial interest(s): All authors are either past or present employees of ACADIA Pharmaceuticals Inc. and hold stock in the company.
Supplementary Material
References
- Cui J.; Shen Y.; Li R. (2013) Estrogen synthesis and signaling pathways during aging: from periphery to brain. Trends Mol. Med. 19, 197–209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu W.-F.; Tan X.-J.; Dai Y.-B.; Krishnan V.; Warner M.; Gustafsson J.-Å. (2013) Targeting estrogen receptor β in microglia and T cells to treat experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. U.S.A. 110, 3543–3548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wisdom A. J.; Cao Y.; Itoh N.; Spence R. D.; Voskuhl R. R. (2013) Estrogen receptor-β ligand treatment after disease onset is neuroprotective in the multiple sclerosis model. J. Neurosci. Res. 91, 901–908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baraka A. M.; Korish A. A.; Soliman G. A.; Kamal H. (2011) The possible role of estrogen and selective estrogen receptor modulators in a rat model of Parkinson’s disease. Life Sci. 88, 879–885. [DOI] [PubMed] [Google Scholar]
- Rosario E. R.; Chang L.; Head E. H.; Stanczyk F. Z.; Pike C. J. (2011) Brain levels of sex steroid hormones in men and women during normal aging and in Alzheimer’s disease. Neurobiol. Aging 32, 604–613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Razmara A.; Duckles S. P.; Krause D. N.; Procaccio V. (2007) Estrogen suppresses brain mitochondrial oxidative stress in female and male rats. Brain Res. 1176, 71–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pike C. J.; Carroll J. C.; Rosario E. R.; Barron A. M. (2009) Protective actions of sex steroid hormones in Alzheimer’s disease. Front. Neuroendocrinol. 30, 239–258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpkins J. W.; Yi K. D.; Yang S.-H.; Dykens J. A. (2010) Mitochondrial mechanisms of estrogen neuroprotection. Biochim. Biophys. Acta 1800, 1113–1120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C.-S.; Tseng Y.-T.; Hsu Y.-Y.; Lo Y.-C. (2013) Nrf2-Keap1 Antioxidant Defense and Cell Survival Signaling Are Upregulated by 17β-Estradiol in Homocysteine-Treated Dopaminergic SH-SY5Y Cells. Neuroendocrinology 97, 232–241. [DOI] [PubMed] [Google Scholar]
- Grimm A.; Lim Y.-A.; Mensah-Nyagan A. G.; Götz J.; Eckert A. (2012) Alzheimer’s disease, oestrogen and mitochondria: an ambiguous relationship. Mol. Neurobiol. 46, 151–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saido T.; Leissring M. A. (2012) Proteolytic Degradation of Amyloid β-Protein. Cold Spring Harbor Perspect. Med. 10.1101/cshperspect.a006379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang K.; Yang L.; Yin C.; Xiao Z.; Zhang J.; Liu Y.; Huang J. (2010) Estrogen stimulates degradation of beta-amyloid peptide by up-regulating neprilysin. J. Biol. Chem. 285, 935–942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao M.; Nguyen T.-V. V.; Rosario E. R.; Ramsden M.; Pike C. J. (2008) Androgens regulate neprilysin expression: role in reducing beta-amyloid levels. J. Neurochem. 105, 2477–2488. [DOI] [PubMed] [Google Scholar]
- Liu F.; Day M.; Muñiz L. C.; Bitran D.; Arias R.; Revilla-Sanchez R.; Grauer S.; Zhang G.; Kelley C.; Pulito V.; Sung A.; Mervis R. F.; Navarra R.; Hirst W. D.; Reinhart P. H.; Marquis K. L.; Moss S. J.; Pangalos M. N.; Brandon N. J. (2008) Activation of estrogen receptor-beta regulates hippocampal synaptic plasticity and improves memory. Nat. Neurosci. 11, 334–343. [DOI] [PubMed] [Google Scholar]
- Yi K. D.; Perez E.; Yang S.; Liu R.; Covey D. F.; Simpkins J. W. (2011) The assessment of non-feminizing estrogens for use in neuroprotection. Brain Res. 1379, 61–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piu F.; Cheevers C.; Hyldtoft L.; Gardell L. R.; Del Tredici A. L.; Andersen C. B.; Fairbairn L. C.; Lund B. W.; Gustafsson M.; Schiffer H. H.; Donello J. E.; Olsson R.; Gil D. W.; Brann M. R. (2008) Broad modulation of neuropathic pain states by a selective estrogen receptor beta agonist. Eur. J. Pharmacol. 590, 423–429. [DOI] [PubMed] [Google Scholar]
- Roman-Blas J. A.; Castañeda S.; Cutolo M.; Herrero-Beaumont G. (2010) Efficacy and safety of a selective estrogen receptor β agonist, ERB-041, in patients with rheumatoid arthritis: a 12-week, randomized, placebo-controlled, phase II study. Arthritis Care Res. 62, 1588–1593. [DOI] [PubMed] [Google Scholar]
- Lund T. D.; Hinds L. R.; Handa R. J. (2006) The androgen 5alpha-dihydrotestosterone and its metabolite 5alpha-androstan-3beta, 17beta-diol inhibit the hypothalamo-pituitary-adrenal response to stress by acting through estrogen receptor beta-expressing neurons in the hypothalamus. J. Neurosci. 26, 1448–1456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weihua Z.; Lathe R.; Warner M.; Gustafsson J.-Å. (2002) An endocrine pathway in the prostate, ERbeta, AR, 5alpha-androstane-3beta,17beta-diol, and CYP7B1, regulates prostate growth. Proc. Natl. Acad. Sci. U.S.A. 99, 13589–13594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- George S., Petit G. H., Gouras G. K., Brundin P., and Olsson R. (2013) Selective androgen and estrogen receptor modulating compounds moderate cognitive deficits and amyloid-β levels in a mouse model of Alzheimer’s disease. Manuscript submitted. [DOI] [PMC free article] [PubMed]
- Tao Q.; Fan X.; Li T.; Tang Y.; Yang D.; Le W. (2012) Gender segregation in gene expression and vulnerability to oxidative stress induced injury in ventral mesencephalic cultures of dopamine neurons. J. Neurosci. Res. 90, 167–178. [DOI] [PubMed] [Google Scholar]
- Burstein E. S.; Piu F.; Ma J.-N.; Weissman J. T.; Currier E. A.; Nash N. R.; Weiner D. M.; Spalding T. A.; Schiffer H. H.; Del Tredici A. L.; Brann M. R. (2006) Integrative functional assays, chemical genomics and high throughput screening: harnessing signal transduction pathways to a common HTS readout. Curr. Pharm. Des. 12, 1717–1729. [DOI] [PubMed] [Google Scholar]
- Gardell L. R.; Hyldtoft L.; Del Tredici A. L.; Andersen C. B.; Fairbairn L. C.; Lund B. W.; Gustafsson M.; Brann M. R.; Olsson R.; Piu F. (2008) Differential modulation of inflammatory pain by a selective estrogen receptor beta agonist. Eur. J. Pharmacol. 592, 158–159. [DOI] [PubMed] [Google Scholar]
- Schaudig K.; Thomssen C. (2007) Hormonal therapy with patch or pill: how much does it matter?. Thromb. Haemostasis 97, 503–504. [PubMed] [Google Scholar]
- Ebert C. D., John V. A., Beall P. T., and Rosenzweig K. A. (1987) Transbuccal Absorption of Diclofenac Sodium in a Dog Model. In Controlled-Release Technology (Lee P. I., and Good W. R., Eds.), pp 310–321, American Chemical Society, Washington, DC. [Google Scholar]
- McFarland K.; Price D. L.; Bonhaus D. W. (2011) Pimavanserin, a 5-HT2A inverse agonist, reverses psychosis-like behaviors in a rodent model of Parkinson’s disease. Behav. Pharmacol. 22, 681–692. [DOI] [PubMed] [Google Scholar]
- Lema Tomé C. M.; Tyson T.; Rey N. L.; Grathwohl S.; Britschgi M.; Brundin P. (2012) Inflammation and α-Synuclein’s Prion-like Behavior in Parkinson’s Disease—Is There a Link?. Mol. Neurobiol. 47, 561–574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillies G. E.; McArthur S. (2010) Estrogen actions in the brain and the basis for differential action in men and women: a case for sex-specific medicines. Pharmacol. Rev. 62, 155–198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McAllister C.; Long J.; Bowers A.; Walker A.; Cao P.; Honda S.-I.; Harada N.; Staufenbiel M.; Shen Y.; Li R. (2010) Genetic targeting aromatase in male amyloid precursor protein transgenic mice down-regulates beta-secretase (BACE1) and prevents Alzheimer-like pathology and cognitive impairment. J. Neurosci. 30, 7326–7334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang K. C. N.; Wang Y.; Oh I. G.; Jenkins S.; Freedman L. P.; Thompson C. C.; Chung J. H.; Nagpal S. (2010) Estrogen receptor beta is a novel therapeutic target for photoaging. Mol. Pharmacol. 77, 744–750. [DOI] [PubMed] [Google Scholar]
- Tiwari-Woodruff S.; Morales L. B. J.; Lee R.; Voskuhl R. R. (2007) Differential neuroprotective and antiinflammatory effects of estrogen receptor (ER)alpha and ERbeta ligand treatment. Proc. Natl. Acad. Sci. U.S.A. 104, 14813–14818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao X.; Cassidy A.; Schwarzschild M. A.; Rimm E. B.; Ascherio A. (2012) Habitual intake of dietary flavonoids and risk of Parkinson disease. Neurology 78, 1138–1145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geyer M. A., and Swerdlow N. R. (2001) Measurement of Startle Response, Prepulse Inhibition, and Habituation. In Current Protocols in Neuroscience, John Wiley & Sons, Inc., Hoboken, NJ. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.