

Abstract
There is growing evidence that terrestrial ecosystems are exporting more dissolved organic carbon (DOC) to aquatic ecosystems than they did just a few decades ago. This “browning” phenomenon will alter the chemistry, physics, and biology of inland water bodies in complex and difficult-to-predict ways. Experiments provide an opportunity to elucidate how browning will affect the stability and functioning of aquatic ecosystems. However, it is challenging to obtain sources of DOC that can be used for manipulations at ecologically relevant scales. In this study, we evaluated a commercially available source of humic substances (“Super Hume”) as an analog for natural sources of terrestrial DOC. Based on chemical characterizations, comparative surveys, and whole-ecosystem manipulations, we found that the physical and chemical properties of Super Hume are similar to those of natural DOC in aquatic and terrestrial ecosystems. For example, Super Hume attenuated solar radiation in ways that will not only influence the physiology of aquatic taxa but also the metabolism of entire ecosystems. Based on its chemical properties (high lignin content, high quinone content, and low C:N and C:P ratios), Super Hume is a fairly recalcitrant, low-quality resource for aquatic consumers. Nevertheless, we demonstrate that Super Hume can subsidize aquatic food webs through 1) the uptake of dissolved organic constituents by microorganisms, and 2) the consumption of particulate fractions by larger organisms (i.e., Daphnia). After discussing some of the caveats of Super Hume, we conclude that commercial sources of humic substances can be used to help address pressing ecological questions concerning the increased export of terrestrial DOC to aquatic ecosystems.
Introduction
Aquatic ecosystems are connected to the surrounding landscape through inputs of material and energy from terrestrial ecosystems. It is estimated that inland water bodies receive nearly three petagrams of terrestrial carbon on an annual basis [1], [2], [3]. The majority of this organic matter, comprised of humic substances derived from vascular plants, is delivered to streams, lakes, and estuaries in the form of dissolved organic carbon (DOC). Terrestrial DOC influences the physical, chemical, and biological features of recipient aquatic ecosystems in many ways. For example, the chromophoric properties of terrestrial DOC can affect the physiology and behavior of aquatic organisms by reducing UV stress [4], and at the same time, reduce rates of primary productivity via the attenuation of photosynthetically active radiation [5]. In addition, terrestrial DOC forms complexes with trace elements and nutrients, which influences the turnover and bioavailability of these resources for aquatic biota [6], [7]. Last, terrestrial DOC is made up of allochthonous compounds that can subsidize aquatic food webs [8] and regulate net ecosystem production [9].
Growing evidence suggests that DOC concentrations in aquatic ecosystems are rising in certain parts of the world [10], [11]. It has been hypothesized that this “browning” trend may be associated with drivers of global change, including altered land-use [12], precipitation patterns [13], temperatures [14], and atmospheric deposition [15]. Regardless of the controlling factors, there is considerable uncertainty about how aquatic ecosystems will respond to increasing inputs of terrestrial DOC [16]. Insight about the effects of browning can be gained from comparative studies [17], but these space-for-time substitutions are correlative and may not capture the full range of interactions that aquatic ecosystems will experience in the future.
Experimental approaches provide an opportunity to address uncertainty about global change scenarios, including the effects of browning. However, it is challenging for aquatic scientists to obtain enough terrestrial DOC to conduct manipulative studies at ecologically relevant scales. For small-scale experiments (∼1L), it is possible to extract milligram to gram quantities of natural humic substances to address questions related to microbial activity over short periods of time [18]. But larger experimental units (and more DOC) are required if one is interested in elucidating the interactive effects that terrestrial DOC has on food webs, physical processes, and ecosystem functioning. Terrestrial DOC has been manipulated in aquatic mesocosms (∼10–1,000 L) through the addition of leachates from soils [19], deciduous leaves [20], or peat moss [21]. The downside of this approach, however, is that leaching sometimes involves artificial processing of the organic material [22], [23]. In addition, the chemical composition of freshly leached DOC may be distinct from the older humic substances that typically dominate DOC in aquatic ecosystems [24]. Browning can also be simulated by adding water from humic-rich systems to mesocosms containing low DOC water from another system [25], [26], but this mixing approach may be confounded by the transfer of organisms and unintended effects on mesocosm water chemistry. Last, terrestrial DOC can be manipulated at the ecosystem scale. For example, concentrations of terrestrial DOC have been indirectly elevated by altering hydrological flows through the installation of curtains in relatively small lakes [27], [28]. And in one rather unique case, 23,000 L of humic water originating from the Suwanee River (Georgia, USA) was transported nearly 3,000 km in a tanker truck before being released into two high elevation rivers (Colorado, USA) [29]. Unfortunately, these types of large-scale manipulation are not always practical or easy to control, nor are they amenable to replication [30].
As an alternative to the experimental approaches described above, DOC manipulations could potentially be performed at a range of scales using concentrated solutions of humic substances, which can readily be obtained in large quantities from commercial suppliers. Humic substances are commonly used as an amendment in agricultural ecosystems to increase soil water-holding capacity, reduce erosion, improve pH buffering capacity, and stimulate the activity of beneficial microorganisms [31]. Many of the commercially available sources of humic substances are derived from Leonardite, a highly oxidized mineral-like substance that is extracted from geologic deposits in North America and other parts of the world [32]. Previous studies have investigated the chemical and physical properties of Leonardite-derived materials to understand how humic substances affect the behavior of metals and the solubility of organic pollutants [33], [34], [35]. It is yet to be determined whether or not commercially available humic substances might serve as an adequate analog for addressing ecological questions about the importance of terrestrial DOC in aquatic ecosystems. Therefore, in this study, we used a combination of analytical procedures, laboratory assays, and comparative surveys to describe the chemical and physical characteristics of a commercially available source of humic substances. In the process, we identify a putative set of heterotrophic bacteria that are capable of growing on humic substances and report results from a series of whole-pond manipulations that explore the relationship between DOC loading and microbial metabolism. Last, we discuss the opportunities and caveats of using commercially available sources of humic substances for addressing ecological questions about terrestrial DOC in aquatic ecosystems.
Materials and Methods
Background
We compared the properties of a commercially available source of humic substances to DOC in natural ecosystems using various approaches, including whole-ecosystem manipulations. Specifically, we altered the supply rate of humic substances to 11 ponds at the Kellogg Biological Station (KBS) Experimental Pond Facility at Michigan State University. Each of the vinyl-lined earthen ponds measures 30 m in diameter, has a maximum depth of 2 m, and an operating volume of approximately 106 L. Based on concentrations of total phosphorus, the ponds are considered mesotrophic (0.48–0.65 µmol P L−1) and contain plankton assemblages that are typical of lentic habitats in the region [36]. Prior to initiating our experiment, we filled each of the ponds with water from a groundwater well and allowed approximately one month for acclimation. We then established a gradient of terrestrial carbon supply (0 to 177 mmol C m−2 d−1) by adding different quantities of a commercially available source of humic substances to each pond. The humic substances used in our study came from a product called “Super Hume”, which is marketed by CropMaster, United Agricultural Services of America, Inc. (Lake Panasoffkee, Florida, USA). Super Hume is extracted from Leonardite shale, a naturally occurring oxidized form of lignite coal. The extraction involves treatment with potassium hydroxide, which yields a basic (pH = 11.5) and concentrated (15% fulvic acid and 4% humic acid, dry mass) solution of humic substances. Super Hume was shipped to the Experimental Pond Facility in 1,000 L carboys and added to each pond on a weekly basis using a gas-powered pump. We sampled water from the center of each non-aerated pond in a jon boat using a 1-m depth-integrated tube sampler three times per week for approximately 14 weeks from early June to mid September 2009. We monitored various limnological variables during our sampling. Some of these methods and results are discussed below, while other findings are the focus of another manuscript [37].
Physical Characteristics: Size Distribution
The size distribution of humic substances is important for determining the physical, chemical, and biological behavior of organic matter in aquatic ecosystems. To assess the size distribution of Super Hume at a relatively coarse resolution, we first quantified the total organic carbon concentration of Super Hume solutions (n = 3) diluted in Milli-Q filtered water (target concentration = 2 mmol L−1) via oxidation and subsequent nondispersive infrared (NDIR) detection using a Shimadzu TOC-V carbon analyzer. We then measured the amount of organic carbon in different size classes based on the concentration of organic carbon that passed through Whatman (Kent, UK) glass microfiber filters (GF/D = 2.7 µm and GF/F = 0.7 µm) and Supor (Pall Corporation, Port Washington, NY, USA) polyethersulfone (PES) membrane filters (0.2 µm). This filtration process allowed us to quantify the relative amounts of Super Hume in four size classes: <0.2 µm; 0.2–0.7 µm; 0.7–2.7 µm; >2.7 µm.
Physical Characteristics: Flocculation
At the beginning of our field experiment, we observed that DOC concentrations in the experimental ponds receiving Super Hume declined at a faster rate than we had anticipated. This pattern suggested that Super Hume was undergoing flocculation, a process whereby DOC aggregates into particulate organic carbon (POC), which is subject to sinking. To test our hypothesis, we estimated flocculation rates for Super Hume under laboratory conditions. We added different amounts of Super Hume to nine 1-L polycarbonate bottles containing unfiltered water from a KBS reference pond. Flocculation was estimated as the loss of DOC (<0.7 µm) in non-agitated bottles that were incubated at room temperature for four days. We did not account for biological production or consumption of DOC during the incubations. The flocculation rate constant was quantified by regressing the DOC flocculation rate (µmol L−1 d−1) against the initial DOC concentration (µmol L−1). To assess the potential for background water chemistry to affect sedimentation, we also estimated flocculation with water collected from six lakes (North Gate Pond, Morris Lake, Roach Lake, Kickapoo Lake, Peter Lake, Long Lake) at the University of Notre Dame Environmental Research Center (UNDERC) in northern Wisconsin. For these assays, flocculation rates were only measured at a single Super Hume concentration (667 µmol DOC L−1) as opposed to the full gradient (0–3,333 µmol DOC L−1) for water from the KBS reference pond.
Physical Characteristics: Optical Properties
We assessed the light-absorbing properties of Super Hume by comparing color and Secchi depth along DOC gradients in the KBS experimental ponds and natural lakes. Color was quantified on surface water samples spectrophotometrically using a 10 cm quartz cuvette and expressed as a440, where a440 = 2.303 (absorbance at 440 nm/0.1 m) [38] while Secchi depth was measured following standard procedures [39]. For natural systems, data for color and Secchi depth were obtained from lakes in New England and upstate New York [17], Wisconsin [40], and Michigan (Lennon unpublished data). For the experimental ponds, we used time-averaged estimates for color (n = 36) and Secchi depth (n = 12). We then compared the color-DOC and Secchi-DOC relationships using indicator variables multiple regression [41] where DOC was treated as a continuous predictor variable and ecosystem type (experimental pond vs. natural lake) was treated as a categorical predictor variable. We log10-transformed our data when necessary to help meet the assumptions of normality and equal variance. Differences in the slopes or intercepts between ecosystem types would suggest that Super Hume differentially affected the optical properties of water when compared to natural sources of DOC.
Chemical Characteristics: Elemental Ratios
The elemental composition of terrestrial organic matter is an important feature that can influence resource quality for aquatic consumers. Therefore, first, we measured total nitrogen (TN) and total phosphorus (TP) after persulfate digestion using spectrophotometric analysis [42]. Second, we analyzed the concentrations of major cations and anions on a diluted Super Hume sample (800 µmol DOC L−1) using a Dionex membrane-suppression ion chromatograph. Last, we estimated total alkalinity by acid titration, as well as by charge balance of the major cations and anions. We then compared the elemental chemistry of Super Hume to observations from a database of streams and rivers (n = 78), groundwater samples (n = 58), lakes (n = 111), wetlands (n = 134), and soil-water samples (n = 142) that were collected in southwestern Michigan between 1996–2009 using methods described elsewhere [43], [44], [45]. In order to compare the chemistry of Super Hume to environmental samples, we standardized the molar concentration (or equivalents, in the case of alkalinity) by the molar DOC concentration of each sample.
Chemical Characteristics: Organic Properties
We characterized the organic chemical properties of Super Hume using two methods. First, we compared Super Hume to aquatic and terrestrial reference materials using pyrolysis-gas chromatography/mass spectrometry (py-GC/MS). The seven reference materials consisted of four soil samples, an algal sample, a DOC sample from a eutrophic lake (Wintergreen Lake, MI), and a DOC sample from a dystrophic lake (Brandywine Lake, MI). The soil reference samples were fine-loamy, mixed, mesic typic Hapludalfs obtained from surface soils of agricultural sites located near the KBS Experimental Pond Facility. The algal sample was prepared from a laboratory culture of Ankistrodesmus sp., which was dried at 60°C before pyrolysis, while carbon from the lake DOC samples was obtained by collecting organic matter after evaporating 1 L of 0.7 µm-filtered water samples. A detailed description of the py-GC/MS methods can be found elsewhere [46]. Briefly, after sample pyrolysis at 600°C, compounds were separated and identified using gas chromatography and ion trap mass spectrometry. Peaks were identified using the National Institute of Standards and Technology (NIST) compound library and were subsequently binned into six primary chemical classes (lignin, lipids, phenols, nitrogen-bearing compounds, polysaccharides, and compounds of unknown origin). Data are reported as relative abundances and represent proportions of the total ion signal characterized during analysis. We visualized the multivariate data for the different samples using Principal Coordinates Analysis (PCoA) with a Bray-Curtis distance matrix. We used the envfit function in the vegan package of R [47] to project vectors onto our ordination to visualize correlations between chemical attributes and our reference samples.
Second, we characterized Super Hume using fluorescence spectroscopy and parallel factor analysis (PARAFAC). Using a Perkin Elmer LS50B fluorometer, we generated excitation-emission matrix spectra (EEM) from a 3D fluorescence scan (excitation: 240–450 every 10 nm; emission 350–550 every 2 nm) on a diluted Super Hume sample (883 µmol C L−1). We corrected the EEM by implementing manufacturer-supplied correction files and by subtracting the resulting values from Milli-Q water blanks. Prior to analysis, we normalized the data to the area under the water Raman peak (excitation 350 nm) [48]. We then fit the Super Hume fluorescence data to a preexisting and validated PARAFAC model, which statistically decomposed the EEM spectra of the sample into loading components related to the organic matter constituents. This model yields 13 loading components based on 379 samples from diverse aquatic habitats [49]. We used the model fit, residuals, and loadings of the 13 model components to characterize the organic matter properties of Super Hume relative to other surface waters.
Chemical Characteristics: Isotopic Composition
Because they can be used to help track the fate of carbon in food webs and ecosystems, we quantified the stable and radioactive carbon isotope ratios of Super Hume. Dried Super Hume was analyzed for δ13C at the University of California Davis Stable Isotope Facility with a PDZ Europa trace gas analyzer and a continuous-flow Europa 20/20 isotope ratio mass spectrometer (IRMS). The Δ14C of Super Hume was estimated from graphite targets at the Center for Accelerator Mass Spectrometry at Lawrence Livermore National Laboratory after subtracting background measurements of 14C-free coal [50].
Biological Responses: Zooplankton Life History
We observed what appeared to be humic substances in the guts of zooplankton from a Super Hume-enriched pond. To evaluate the potential effects that Super Hume might have on zooplankton fitness, we conducted a life table experiment using a clone of Daphnia pulex x pulicaria isolated from a KBS pond that had received Super Hume over the course of the growing season (177 mmol C m−2 d−1). Prior to initiating the life table experiment, we propagated the Daphnia clone for multiple generations at room temperature in COMBO medium [51] and fed the animals with a culture of green algae (Ankistrodesmus sp.).
To initiate the life table experiment, we randomly assigned 15 neonates to a control treatment without Super Hume (−SH) and 15 neonates to a Super Hume treatment (+SH) containing 1,666 µmol DOC L−1. We then followed the methods for life table analysis described in detail elsewhere [52]. Briefly, individual Daphnia were grown in 70 mL tissue-culture flasks containing COMBO medium and 20,000 cells mL−1 of Ankistrodesmus in an environmental chamber (25°C, 16∶8 light-dark cycle) for 21 days. Each day, we recorded whether an individual was alive or dead (survivorship) and the number of neonates that were produced (reproduction). At this time, we also transferred the focal daphnid to a new flask containing fresh medium, food, and Super Hume (depending on the treatment).
We used Euler’s method to calculate the intrinsic rate of increase, r (d −1): 1 = Σ e−r x l(x) b(x), where b(x) is number of offspring produced per individual on day x, and l(x) is the proportion of individuals in a treatment surviving to the next day. We estimated the standard error for the intrinsic rate of increase using jackknifing procedures [53]. In addition, we calculated the net reproductive rate (R o) as the sum of offspring produced for individuals and generation time as Σx l(x) b(x)/Σ l(x) b(x).
Last, we estimated the ingestion rates and assimilation rates of Daphnia in the −SH and +SH treatments by feeding animals suspensions of Ankistrodesmus (50,000 cells mL−1) that had been labeled with H14CO3 − [54]. We measured ingestion rates based on the counts per minute (CPM) of individual Daphnia after two hours of feeding. Assimilation rates were estimated by quantifying the CPM of labeled Daphnia after they had incubated in COMBO medium without Ankistrodesmus for two hours. We then estimated assimilation efficiency as the ratio of assimilation rates to ingestion rates. We used a Kaplan-Meier test to determine the effects of Super Hume on Daphnia survivorship; for all other life history traits, we performed statistical comparisons using Student’s t-tests.
Biological Responses: Humic Oxidizing Bacteria
We isolated bacteria from the experimental pond that received the highest supply rate (177 mmol C m−2 d−1) of Super Hume. Water was collected from the surface (0.5 m) of the pond in early August 2009 and immediately transported back to the lab where dilutions of the samples were spread onto washed agar plates (1.5%) containing a modified version of WC medium [55] (we substituted NH4Cl for KNO3 as the nitrogen source and did not include Na2SiO3 or H2O3Se). Super Hume was added to the plates at a final concentration of 3,333 µmol DOC L−1. We then watched for colony formation while incubating the plates in an aerobic chamber at 25°C in the dark for up to three weeks. To purify isolates, we picked single colonies and restruck them onto Super Hume plates before cryopreservation in 20% glycerol at −80°C. We identified the bacterial isolates by direct sequencing of the 16S rRNA gene. Briefly, DNA was extracted using the UltraClean® Microbial DNA Isolation Kit (Mo-Bio, Carlsbad, CA). We then used 5 ng of this DNA as a template in a polymerase chain reaction (PCR) using 8F and 1492R primers with thermal cycler conditions outlined elsewhere [56]. The resulting PCR products were sequenced at the Research Technology Support Facility (RTSF) at Michigan State University (East Lansing, Michigan, USA). We used the classifier tool in the Ribosomal Database Project [57] to identify each of the bacterial isolates. For visual purposes, we aligned our sequences in mothur [58] version 1.25.1 against the Silva reference database and generated a phylogenetic tree using the neighbor-joining algorithm in ClustalX (www.clustal.org). We deposited the 16S rRNA gene sequences in GenBank with the accession numbers JX312319– JX312328.
Biological Responses: Microbial Metabolism
We examined the relationships between microbial metabolism and DOC supply in the experimental ponds that were enriched with Super Hume. We estimated bacterial productivity (BP) as the uptake and incorporation of 3H-leucine (50 nmol L−1 final concentration) into bacterial protein [59], bacterial respiration (BR) as the rate of dissolved oxygen depletion in GF/D-filtered (2.7 µm) samples using a Presens SensorDish Reader [60] and bacterial growth efficiency (BGE) as BP/(BP+BR). Microbial metabolism and DOC concentration were measured in each of the 11 ponds three times per week for 12 weeks. We evaluated the relationship between microbial metabolism and DOC using simple linear regression on time-averaged values.
Results
Physical Characteristics: Size Distribution
Super Hume contained a mixture of dissolved and fine particulate organic matter. Unfiltered Super Hume samples had a total organic carbon concentration of approximately 3.3 moles C L−1. The vast majority (80%) of this material could operationally be defined as DOC, with 40±6.0% in the <0.2 µm size class and 20±10.0% in the 0.2–0.7 µm size class. The remaining fraction of Super Hume (20%) could be operationally defined as particulate organic carbon (POC), with 11±1.0% in the 0.7–2.7 µm size class and 9±7.0% in the >2.7 µm size class. All values represent mean ± SEM.
Physical Characteristics: Flocculation
Our laboratory assays supported the hypothesis that Super Hume was undergoing flocculation in the experimental ponds. We observed that Super Hume was sedimenting to the bottom of the bottles during our incubations. This was confirmed by a significant loss of DOC over time from incubated pond water samples, which increased with Super Hume concentration (r2 = 0.89, F1,10 = 55.1, P<0.0001, Fig. 1):
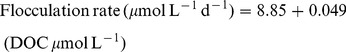 |
(1) |
Figure 1. Flocculation rates of Super Hume.

We estimated flocculation rates in laboratory assays as the loss of dissolved organic carbon (DOC) at different Super Hume concentrations added to water from a reference pond. Regression lines represent predicted values and 95% confidence intervals.
Our results indicate that 4.7–12.2% of the Super Hume standing stock was lost each day to sedimentation. However, the flocculation rates measured in northern Wisconsin lakes (4.5±0.92 µmol DOC L−1 d−1) at a given Super Hume concentration (667 µmol DOC L−1) were nine-fold lower than what was observed in the water from the experimental ponds (one sample t-test, t5 = −16.3, P<0.0001).
Physical Characteristics: Optical Properties
In general, Super Hume and natural DOC had similar effects on the optical properties of aquatic ecosystems. The multiple regression model for color was highly significant (R2 = 0.84, F3,110 = 188, P<0.0001), but the slope and intercepts were affected by ecosystem type, which resulted in the following equations:
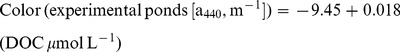 |
(2) |
and
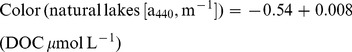 |
(3) |
Despite the differences in the regression parameters, most of the color observations from the experimental ponds fell within the 95% prediction intervals associated with the regression model for natural lakes in equation 3 (Fig. 2a).
Figure 2. Super Hume effects on the aquatic light-environment.
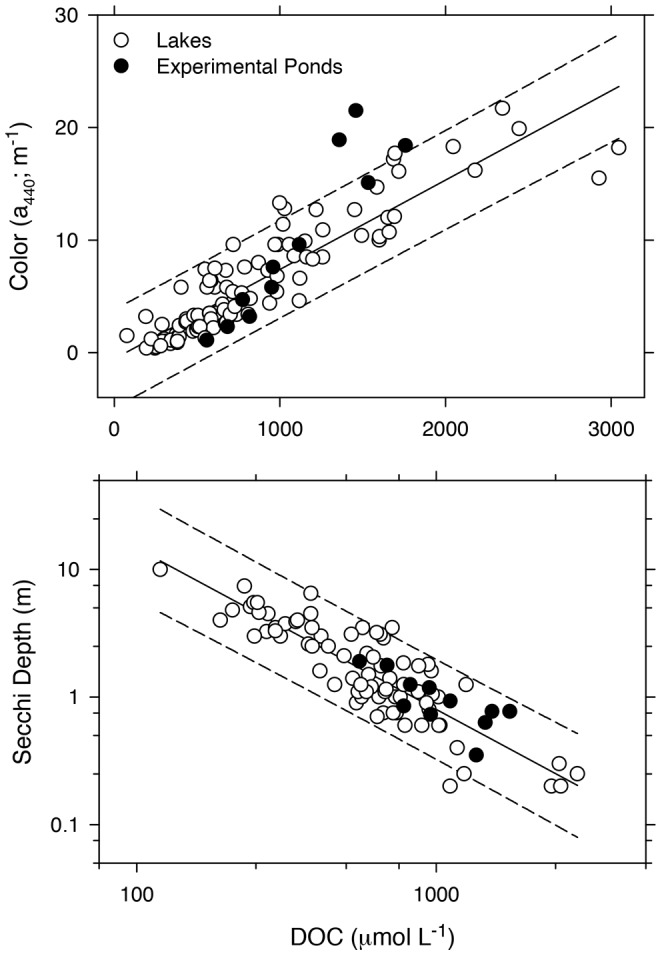
Comparison of color (A) and Secchi depth (B) as a function of DOC in natural lakes and experimental ponds. Variation in DOC concentration in the experimental ponds was achieved by manipulating the Super Hume supply rate. Regression lines represent predicted values and 95% prediction intervals.
Super Hume and natural DOC also had similar effects on Secchi depth, with no differences between natural lakes and experimental ponds (R2 = 0.76, F3,84 = 88.2, P<0.0001). Neither the slope nor the intercept was affected by ecosystem type, so a single model was used to fit the two data sets (Fig. 2b).
| (4) |
Chemical Characteristics: Elemental Ratios
Super Hume is carbon-rich material with low nitrogen (N) and phosphorus (P) content (C:N = 440, C:P = 10,128 by moles). In addition, the N:P ratio of Super Hume was relatively high (276). With the exception of potassium (K), our results suggest that Super Hume amendments would only marginally affect the concentrations of major cations and anions found in streams, groundwater, lakes, wetlands, and soils in southwestern Michigan (Fig. 3). The K:DOC ratio of Super Hume was relatively high (0.247) compared to the median K:DOC ratios observed in wetlands (0.019), lakes (0.057), streams (0.087), groundwater (0.108), and soils (0.190). The alkalinity of Super Hume was 0.35±0.061 meq L−1 (mean ± SEM), which is relatively low compared to the alkalinity of natural systems (Fig. 3).
Figure 3. Super Hume stoichiometry.

Molar ratios of major cation and anions with respect to DOC. We express alkalinity as equivalents per mole DOC. Observations for natural ecosystems (stream, groundwater, lakes, wetlands, and soil water) come from a survey of sites distributed throughout southwestern Michigan. Super Hume values are represented by the horizontal dashed line.
Chemical Characteristics: Organic Properties
The py-GC/MS data suggest that the chemical composition of Super Hume is similar to that of soil organic matter (Fig. 4). The first two axes of the principal coordinates analysis (PCoA) explained >80% of the variation in the py-GC/MS data. The first axis clearly separated soil samples from aquatic samples. Along this axis, Super Hume grouped closely with the soil samples, which had high relative abundances of lignin, protein, and polysaccharides. In contrast, algae and DOC from Wintergreen Lake (eutrophic) had a high lipid content, while the DOC from Brandywine Lake (dystrophic) was enriched with fatty acid methyl esters. The second axis indicated that the organic chemistries of DOC from the eutrophic and dystrophic lakes were distinct, while Super Hume was intermediate to these aquatic endmember samples.
Figure 4. Multivariate ordination of Super Hume chemistry.

We performed a Principal Coordinates Analysis (PCoA) based on the chemical characteristics of Super Hume and other reference organic materials as determined by pyrolysis-gas chromatography/mass spectrometry (py-GC/MS). Reference samples are represented by symbols, while vectors reflect correlations between chemical attributes and the samples.
Most of the variation (99%) in the fluorescent properties of the Super Hume EEM could be explained using the PARAFAC model (Fig. 5) [49]. The residual EEM values (differences between the raw EEM and model fit EEM) were low (<0.2%), indicating that there were no anomalous fluorophores in Super Hume. Oxidized quinone-like compounds (components 11, 2, 12) comprised 27% of the sample (7, 16, 4% respectively), while reduced quinone-like compounds (components 7, 9, 5, 4) comprised 49% of the sample. Component 4, which was a major component of the Super Hume sample (35%), is prevalent in EEMs of isolated humic substances derived from soils [61]. Only three of the 13 model components were not detected in Super Hume. Two of these (components 8 and 13), are indicative of amino acid-like molecules, while the other component (component 3) has been characterized as “unknown” and only recovered from Antarctic samples.
Figure 5. Heat map of Super Hume chemistry.

The heat map represents the excitation-emission matrix spectra (EEMs) from a 3D fluorescence scan of Super Hume that was used in parallel factor analysis (PARAFAC).
Chemical Characteristics: Isotopic Composition
The δ13C of Super Hume was −23.6±0.48‰ (mean ± SEM, n = 4). The Δ14C was −984.8‰, which confirms that the carbon in Super Hume is ancient, as expected based on its origin from lignite.
Biological Responses: Zooplankton Life History
We did not find evidence from our life table experiment that Daphnia pulex x pulicaria was negatively affected by Super Hume, even at fairly high concentrations. Rather, some of our results suggest that Super Hume could potentially increase the fitness of D. pulex x pulicaria. For example, generation times were marginally reduced in the presence (15.5±0.455) vs. absence (16.6±0.129) of Super Hume (t9.31 = 2.18, P = 0.056, Fig. 6D), which contributed in part to an overall higher intrinsic rate of increase (−SH = 0.150±0.0012, +SH = 0.164±0.0012; t28 = −8.20, P<0.0001, Fig. 6E). These effects of Super Hume on population growth could not be attributed to differences in D. pulex x pulicaria ingestion rates (t5.51 = −0.252, P = 0.810) or assimilation efficiencies of algal carbon (t5.95 = −1.19, P = 0.281, Fig. 6F). Super Hume had no effect on survivorship (Chi-square = 1.30, P = 0.246, Fig. 6B) or net reproductive rate (t13.5 = −0.61, P = 0.550, Fig. 6C).
Figure 6. Effects of Super Hume on Daphnia.

A) We observed the accumulation of humic substances in gravid individuals of Daphnia pulex x pulicaria collected from a pond that had been experimentally enriched with Super Hume. Subsequently, we measured the effects of Super Hume (1666 µmol L−1) on the life history (B–D), algal ingestion rates (E), and algal assimilation efficiency (F) where D. pulex x pulicaria was fed algae in the presence (+SH) or absence (−SH) of Super Hume. Values represent means ± SEM.
Biological Responses: Humic Oxidizing Bacteria
We isolated and sequenced ten distinct bacterial isolates that were capable of growing on Super Hume as a sole carbon source. After our initial plating, colonies regrew when struck onto new agar plates containing Super Hume. However, we were not able to grow visibly turbid cultures in the liquid WC medium amended with Super Hume. The phylogenetic identities of the bacteria were not dissimilar to other bacteria that have been cultivated from freshwater ecosystems (Fig. 7). Three of the isolates belonged to the Bacterioidetes phylum, four belonged to the Proteobacteria pylum (two in the β-Proteobacteria class and two in the γ-Proteobacteria class), and three to the Actinobacteria phylum (Fig. 7).
Figure 7. Heterotrophic bacteria grew on Super Hume.

Phylogenetic relationship of bacteria cultivated on Super Hume as a sole carbon source (bold) along with reference sequences (non-bold). Trees were constructed from aligned 16S rRNA sequences using neighbor-joining methods. γ = γ-Proteobacteria and β = β-Proteobacteria. The scale bar represents the sequence dissimilarity. Thermotoga is an archaeon that was used as an outgroup.
Biological Responses: Microbial Metabolism
Despite the growth of bacterial isolates on Super Hume agar plates, we did not observe a significant relationship between bacterial productivity (BP) and DOC concentration in the experimental ponds (r2 = 0.0008, F1,10 = 0.007, P = 0.94, Fig. 8a). In contrast, bacterial respiration (BR) significantly increased along the DOC gradient (r2 = 0.52, F1,10 = 9.85, P = 0.012, Fig. 8b).
| (5) |
Figure 8. Microbial metabolism along a Super Hume gradient.

We created a DOC gradient in a set of experimental ponds by altering the supply rate of Super Hume. There was no relationship between bacterial productivity (BP) and DOC (A), but bacterial respiration (BR) significantly increased (B) and bacterial growth efficiency (BGE) significantly decreased (C) along the DOC gradient. Regression lines represent predicted values and 95% confidence intervals for significant (solid lines) and non-significant (dashed lines) models.
This change in BR resulted in a marginally significant decline in bacterial growth efficiency (BGE) along the DOC gradient (r2 = 0.30, F1,10 = 3.94, P = 0.079, Fig. 8c):
| (6) |
Discussion
In this study, we evaluated a commercially available source of humic substances (Super Hume) for studying the effects of terrestrial DOC on the chemistry, physics, and biology of aquatic ecosystems. Similar to natural organic matter, Super Hume attenuated solar radiation in ways that are not only likely to affect the stress physiology of aquatic taxa, but also the metabolism of entire ecosystems. Super Hume may also provide insight into the effects of allochthonous resource subsidies on aquatic ecosystems. Specifically, our results suggest that Super Hume can enter aquatic food webs through 1) the uptake of dissolved organic constituents by microorganisms, and 2) the consumption of particulate fractions by larger organisms (e.g. crustacean zooplankton). Although the C:N and C:P ratios, potassium content, and flocculation rates could be high, Super Hume had properties that were very similar to natural sources of DOC found in terrestrial and aquatic ecosystems. Thus, Super Hume and other Leonardite-derived humic substances may be useful for addressing questions about the effects of terrestrial carbon export on aquatic ecosystems.
Light Attenuation by Super Hume
The light-absorbing properties of Super Hume were comparable to those of natural DOC. Statistically, the parameters describing the relationship between Secchi Depth and DOC in a diverse set of north temperate lakes were similar to the parameters obtained from experimental ponds enriched with Super Hume (Fig. 2B). Likewise, when estimating light attenuation via color (a440), most observations from the Super Hume-enriched ponds fell within 95% prediction intervals associated with the color-DOC relationship observed for lakes. At high DOC concentrations, however, Super Hume absorbed more light than organic matter found in natural aquatic ecosystems (Fig. 2A). This difference could be due to unique chromophoric properties of Super Hume. Alternatively, the higher carbon-specific light absorbance could reflect features of the experimental ponds. For example, because our experiment took place over a relatively short period of time (4 months), Super Hume may have experienced less photobleaching than DOC found in natural lakes with longer residence times [62]. Our PARAFAC results provide additional insight into the spectral properties of Super Hume (Fig. 5), but more sophisticated optical analyses could help elucidate how DOC influences species interactions and ecosystem processes [63].
Microbes Subsidized by Super Hume
Super Hume may provide insight into the mechanisms influencing the variation of allochthony among aquatic ecosystems [8]. Because most terrestrial carbon enters aquatic ecosystems in a dissolved form [64], it is often assumed – at least in lentic systems – that allochthony is influenced by the uptake of DOC by aquatic bacteria [65]. We isolated bacteria from a DOC-enriched pond that were capable of growing in the laboratory on Super Hume as the sole carbon source. These bacteria came from diverse phyla and were related to other taxa that have been recovered from lakes using cultivation-based approaches (see Fig. 7). Owing to the biases of culture-based methods, it is not surprising that our isolates were not closely related to lake bacteria that have been characterized using cultivation-independent molecular techniques [66]. Nevertheless, the isolates from this study provide unique opportunities to identify genomic and physiological traits that allow aquatic microbes to use recalcitrant carbon substrates. Last, our field data suggest that Super Hume stimulated bacterial respiration (Fig. 8), which increased linearly with increasing Super Hume loading rate, while bacterial productivity remained constant, which led to a slight decrease in bacterial growth efficiency along the gradient. Such findings are consistent with the view that anabolic and catabolic processes can be decoupled [67] and that terrestrial carbon may be preferentially allocated to respiration over growth.
Is Super Hume Channeled Directly to Higher Trophic Levels?
Aquatic food webs can also be subsidized when non-microbial taxa consume terrestrial-derived particulate organic carbon (POC) [68]. Based on our size distribution analysis, 20% of Super Hume can be operationally defined as POC (>0.7 µm), which is comparable to the proportion of DOC and POC that enters natural lakes [69], [70]. This means that some of the Super Hume added to aquatic systems is potentially available for direct consumption by higher trophic levels, including filter-feeding zooplankton like Daphnia [71], [72]. We observed what appeared to be humic substances in the guts of gravid D. pulex x pulicaria from a Super Hume-enriched pond (Fig. 6a). This prompted us to conduct a life table experiment where we grew Daphnia on algae in treatments with or without Super Hume. Compared to the control, we documented that the age at first reproduction was reduced by one day and that there was a 10% increase in the intrinsic rate of increase for Daphnia in the Super Hume treatment (Fig. 6). This apparent stimulation of Daphnia growth by Super Hume could have been accompanied by changes in algal ingestion rates and assimilation rates, but there was no statistical support for this hypothesis. Additional experiments are needed to assess the interactions between terrestrial carbon inputs (DOC and POC) and autochthonous resources (e.g., algae) on the energetics and fitness of Daphnia and other consumer populations.
Implications of Super Hume Flocculation
The fate of Super Hume may also be influenced by interactions between humic substances and other biogeochemical features of aquatic ecosystems. For example, we observed substantial variation in the flocculation rates of Super Hume among ecosystems; this variability is important for understanding the effects of terrestrial carbon loading on planktonic food webs and sediment carbon storage. In the experimental ponds, we estimated that approximately 5–12% of the total DOC pool was lost each day to flocculation. However, flocculation rates were almost an order of magnitude lower when measured in a set of lakes in northern Wisconsin. Physical forces and microbiological activity are known to affect flocculation rates [70], [73], but the aggregation of DOC into colloids and POC can be influenced by water chemistry, too. For example, alkaline water bodies tend to have higher concentrations of Ca2+ and Mg2+, which can promote DOC flocculation [74]. The background alkalinity of the experimental ponds was approximately 3 meq L−1, which is typical for inland water bodies of southwestern Michigan. In contrast, the lakes sampled in northern Wisconsin are ionically dilute and have much lower alkalinity (0.1 meq L−1) [75]. Therefore, ionic composition is one important biogeochemical characteristic that should be taken into consideration when attempting to target DOC concentrations via Super Hume additions in aquatic ecosystems.
Opportunities for Isotopic Inference
Natural abundances of carbon isotopes are frequently used to gain insight into the trophic relationships and the degree of allochthony in aquatic ecosystems [76]. The δ13C of Super Hume is approximately −23‰, which is slightly enriched relative to the isotope signature of terrestrial carbon that typically enters north-temperate inland water bodies (−27‰) [17]. Owing to the overlap with other carbon endmembers, it is unlikely that that δ13C of Super Hume will be useful for resolving questions related to allochthony. However, carbon flow in aquatic ecosystems can also be inferred using 14C [77], [78]. The Δ14C of Super Hume is very depleted (−984.8‰), which means that it could be used in future studies to elucidate the effects of terrestrial carbon on aquatic food webs and ecosystem processes.
Other Chemical Considerations
In general, the chemical constituents of Super Hume were similar to natural sources of dissolved organic matter (DOM) found in soils and aquatic ecosystems (Figs. 3, 4, 5). For example, py-GC/MS data suggest that Super Hume has a chemical signature that is similar to soil organic matter, which is enriched in aromatic, aliphatic, and nitrogen-bearing compounds, but also lignin, polysaccharides, and proteins. Super Hume did not group with DOC from a dystrophic lake (Brandywine) based on py-GC/MS, suggesting that the chemistry of terrestrial organic matter is modified by physical and biological processes after being delivered to aquatic ecosystems. We also analyzed Super Hume using a PARAFAC modeling approach [49], which allowed us to compare its fluorescence signal to 379 samples from aquatic ecosystems around the world. Almost all of the variation (99%) in our Super Hume sample could be explained using the PARAFAC model, which means that Super Hume did not have any anomalous spectrofluorimetric properties. Rather, the PARAFAC model revealed that Super Hume was enriched in quinones and other humic substances commonly found in terrestrial-derived organic carbon.
The nitrogen and phosphorus content of organic matter can be important for understanding the effects of resource quality on consumer populations and ecosystem processes [79]. Super Hume had high C:N (440) and C:P (10,128) ratios. In terrestrial systems, the C:N ratio and C:P ratios of bulk soil organic matter are typically much lower (14∶1 and 186∶1, respectively) [80]. In aquatic ecosystems, the C:N and C:P ratios of DOM can be slightly higher than what is reported for soils, especially when attempts are made to physically isolate autochthonous and allochthonous fractions of the DOM pool. For example, the C:N ratio of fulvic acids in the Suwanee River was ∼90∶1 [81], while the C:P ratio of fulvic acids in a Colorado stream were sometimes >3,500∶1 [82]. Based on these comparisons, Super Hume would be considered a low quality resource for aquatic consumers (e.g., microbes and zooplankton). However, this feature of Super Hume may provide opportunities for understanding the effects of DOM stoichiometry on aquatic ecosystems. For example, researchers could add different amounts or forms (e.g., inorganic and organic) of nitrogen and phosphorus to Super Hume-enriched systems to address questions related to DOM quality.
In general, experimental additions of Super Hume should contribute minimally to the concentration of cations and anions found in many aquatic and terrestrial ecosystems (Fig. 3). One exception is the relatively high potassium content of Super Hume, presumably due to the fact that commercial processing of humic substances from Leonardite involves an alkaline (i.e., KOH) extraction. Because it is major constituent of the cytosol and is involved in ion regulation, elevated potassium could affect aquatic biota in different ways. It is generally assumed that potassium limitation is rare [83], but there are instances where enrichment can stimulate the growth of aquatic taxa, including algae [84] and fungal pathogens [85]. In contrast, some studies have suggested the possibility of potassium toxicity. For example, potassium inhibited Microcystis (Cyanobacteria) populations at concentrations >2,800 µmol L−1 [86] and Dinobryon spp. (Chrysophyceae) at concentrations >383 µmol L−1 [87]. Other lines of evidence, however, suggest that phytoplankton are robust to a wide range of potassium concentrations. Chrysophyte biomass was not correlated to potassium concentrations in a survey of Swedish lakes [88]. In addition, we tested for evidence of inhibition by examining the relationship between algal biomass (chlorophyll a) and potassium in the US Environmental Protection Agency’s National Lake Assessment database (http://water.epa.gov/type/lakes/lakessurvey_index.cfm). After accounting for the correlation between total phosphorus (TP) and chlorophyll a (r = 0.75) in a large set of lakes (n = 1,070), there was no effect of potassium concentration (range = 1.4–36,114 µmol L−1) on algal biomass (P = 0.301, r2 = 0.001). In summary, the potassium associated with commercially available humic substances extracted with KOH could be important in certain systems, but was not problematic in the current study (e.g., Fig. 6).
Caveats and Conclusions
In this study, we have explored the possibility of using a commercial source of humic substances to conduct browning experiments, and we have shown how this material is a good analog for natural DOC of terrestrial origin. Compared to natural DOC, Super Hume has similar organic chemical properties and optical effects on light transmission. Moreover, Super Hume was used by at least some heterotrophic bacteria and was not toxic to crustacean zooplankton (i.e., Daphnia pulex x pulicaria).
There are, however, a number of caveats that should be considered. First, the use of commercially available humic substances in ecotoxicology has been scrutinized because of certain distinct chemical properties [33], [35], which influence their interaction with DDT, PCBs, metals, and other contaminants [34]. Second, it is impossible for any single source of organic matter to capture the chemical and physical complexity of organic matter derived from diverse natural settings. Last, it is conceivable that the extraction and processing procedures may vary among companies or even among batches. For example, some companies add exogenous organic matter (e.g., seaweed) to their humic products; although these amendments may be beneficial for agricultural purposes, they risk confounding aquatic ecology experiments. As mentioned above, researchers should be aware of high potassium content, high C:N and C:P ratios, and possibly other attributes that were not considered in the current study. Nonetheless, commercially available humic substances like Super Hume may provide opportunities to address ecological questions using a fairly standardized product that can easily be obtained in large quantities at an affordable cost. Experimental additions of commercially available humic substances will allow researchers to gain insight into the complex physical, chemical, and biological responses that aquatic ecosystems might experience due to browning and other global change scenarios [16].
Acknowledgments
We acknowledge Leigh McCallister for the 14C analysis on the Super Hume sample, Dave Weed for analytical assistance, Brent Lehmkuhl for microbiological support, and Brian Wallace and Justin Rensch for field and laboratory assistance with the pond experiment and life table study. All data are available from the corresponding author upon request.
Funding Statement
This research was supported with funding from the National Science Foundation (DEB-0842441 to JTL an SEJ and DEB-0743402 to SKH and JTL), the Center for Water Sciences at Michigan State University, and the Huron Mountains Wildlife Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Cole JJ, Prairie YT, Caraco NF, McDowell WH, Tranvik LJ, et al. (2007) Plumbing the global carbon cycle: Integrating inland waters into the terrestrial carbon budget. Ecosystems 10: 171–184. [Google Scholar]
- 2. Battin TJ, Kaplan LA, Findlay S, Hopkinson CS, Marti E, et al. (2008) Biophysical controls on organic carbon fluxes in fluvial networks. Nat Geosci 1: 95–100. [Google Scholar]
- 3. Tranvik LJ, Downing JA, Cotner JB, Loiselle SA, Striegl RG, et al. (2009) Lakes and reservoirs as regulators of carbon cycling and climate. Limnol Oceanogr 54: 2298–2314. [Google Scholar]
- 4. Hader DP, Kumar HD, Smith RC, Worrest RC (2007) Effects of solar UV radiation on aquatic ecosystems and interactions with climate change. Photochem Photobiol Sci 6: 267–285. [DOI] [PubMed] [Google Scholar]
- 5. Karlsson J, Byström P, Ask J, Ask P, Persson L, et al. (2009) Light limitation of nutrient-poor lake ecosystems. Nature 460: 506–509. [DOI] [PubMed] [Google Scholar]
- 6. Jones RI, Salonen K, DeHaan H (1988) Phosphorus transformations in the epilimnion of humic lakes: abiotic interactions between dissolved humic materials and phosphate. Freshwat Biol 19: 357–369. [Google Scholar]
- 7.Maranger RJ, Pullin MJ (2003) Elemental complexation of dissolved organic matter in lakes: implications for Fe speciation and the bioavailability of Fe and P. In: Findlay SEG, Sinsabaugh RL, editors. Aquatic Ecosystems: Interactivity of Dissolved Organic Matter: Academic Press. 26–59.
- 8. Carpenter SR, Cole JJ, Pace ML, Van de Bogert M, Bade DL, et al. (2005) Ecosystem subsidies: Terrestrial support of aquatic food webs from 13C addition to contrasting lakes. Ecology 86: 2737–2750. [Google Scholar]
- 9.Ask J, Karlsson J, Jansson M (2012) Net ecosystem production in clear-water and brown-water lakes. Glob Biogeochem Cycle 26.
- 10. Findlay SEG (2005) Increased carbon transport in the Hudson River: unexpected consequence of nitrogen deposition? Front Ecol Environ 3: 133–137. [Google Scholar]
- 11. Monteith DT, Stoddard JL, Evans CD, de Wit HA, Forsius M, et al. (2007) Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 450: 537–U539. [DOI] [PubMed] [Google Scholar]
- 12. Stanley EH, Powers SM, Lottig NR, Buffam I, Crawford JT (2012) Contemporary changes in dissolved organic carbon (DOC) in human-dominated rivers: is there a role for DOC management? Freshwat Biol 57: 26–42. [Google Scholar]
- 13. Parn J, Mander U (2012) Increased organic carbon concentrations in Estonian rivers in the period 1992–2007 as affected by deepening droughts. Biogeochemistry 108: 351–358. [Google Scholar]
- 14. Freeman C, Evans CD, Monteith DT, Reynolds B, Fenner N (2001) Export of organic carbon from peat soils. Nature 412: 785–785. [DOI] [PubMed] [Google Scholar]
- 15. Evans C, Goodale C, Caporn S, Dise N, Emmett B, et al. (2008) Does elevated nitrogen deposition or ecosystem recovery from acidification drive increased dissolved organic carbon loss from upland soil? A review of evidence from field nitrogen addition experiments. Biogeochemistry 91: 13–35. [Google Scholar]
- 16.Hanson PC, Hamilton DP, Stanley EH, Preston N, Langman OC, et al.. (2011) Fate of allochthonous dissolved organic carbon in lakes: a quantitative approach. PLoS ONE 6. [DOI] [PMC free article] [PubMed]
- 17. Lennon JT, Faiia AM, Feng XH, Cottingham KL (2006) Relative importance of CO2 recycling and CH4 pathways in lake food webs along a dissolved organic carbon gradient. Limnol Oceanogr 51: 1602–1613. [Google Scholar]
- 18. Anesio AM, Graneli W, Aiken GR, Kieber DJ, Mopper K (2005) Effect of humic substance photodegradation on bacterial growth and respiration in lake water. Appl Environ Microbiol 71: 6267–6275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Lennon JT, Cottingham KL (2008) Microbial productivity in variable resource environments. Ecology 89: 1001–1014. [DOI] [PubMed] [Google Scholar]
- 20. Geddes P (2009) Decoupling carbon effects and UV protection from terrestrial subsidies on pond zooplankton. Hydrobiologia 628: 47–66. [Google Scholar]
- 21. Klug JL (2005) Bacterial response to dissolved organic matter affects resource availability for algae. Can J Fish Aquat Sci 62: 472–481. [Google Scholar]
- 22. Vinebrooke RD, Leavitt PR (1998) Direct and interactive effects of allochthonous dissolved organic matter, inorganic nutrients, and ultraviolet radiation on an alpine littoral food web. Limnol Oceanogr 43: 1065–1081. [Google Scholar]
- 23. Lennon JT (2004) Experimental evidence that terrestrial carbon subsidies increase CO2 flux from lake ecosystems. Oecologia 138: 584–591. [DOI] [PubMed] [Google Scholar]
- 24. Amon RMW, Benner R (1996) Bacterial utilization of different size classes of dissolved organic matter. Limnol Oceanogr 41: 41–51. [Google Scholar]
- 25. Arvola L, Kankaala P, Tulonen T, Ojala A (1996) Effects of phosphorus and allochthonous humic matter enrichment on the metabolic processes and community structure of plankton in a boreal lake (Lake Pääjärvi). Can J Fish Aquat Sci 53: 1646–1662. [Google Scholar]
- 26. Hansson L-A, Nicolle A, Granéli W, Hallgren P, Kritzberg ES, et al. (2013) Food-chain length alters community responses to global change in aquatic systems. Nature Clim Change 3: 228–233. [Google Scholar]
- 27. Kasprzak P (1993) The use of an artificially divided bog lake in food web studies. Verh Internat Verein Limnol 25: 652–656. [Google Scholar]
- 28. Christensen DL, Carpenter SR, Cottingham KL, Knight SE, LeBouton JP, et al. (1996) Pelagic responses to changes in dissolved organic carbon following division of a seepage lake. Limnol Oceanogr 41: 553–559. [Google Scholar]
- 29. McKnight DM, Hornberg GM, Bencala KE, Boyer EW (2002) In-stream sorption of fulvic acid in an acidic stream: A stream-scale transport experiment. Water Resour Res 38: 10.1029/2001WR000269. [Google Scholar]
- 30. Carpenter SR (1990) Large-scale perturbations - opportunities for innovation. Ecology 71: 2038–2043. [Google Scholar]
- 31. Quilty JR, Cattle SR (2011) Use and understanding of organic amendments in Australian agriculture: a review. Soil Research 49: 1–26. [Google Scholar]
- 32.Ozdoba DM, Blyth JC, Eugler KF, Dinel H, Schnitzer M (2001) Leonadite and humified organic matter. In: Ghabbour EA, Davies G, editors. Humic Substances: Structures, Models and Functions Royal Society of Chemistry.
- 33. Malcolm RL, Maccarthy P (1986) Limitation in the use of commercial humic acids in water and soil research. Environ Sci Technol 20: 904–911. [DOI] [PubMed] [Google Scholar]
- 34. Chiou CT, Kile DE, Brinton TI, Malcolm RL, Leenheer JA, et al. (1987) A comparison of water solubility enhancements of organic solutes by aquatic humic materials and commerical humic acids. Environ Sci Technol 21: 1231–1234. [Google Scholar]
- 35. Grasso D, Chin YP, Weber WJ (1990) Structural and behavioral characteristics of a commercial humic acid and natural dissolved aquatic organic matter. Chemosphere 21: 1181–1197. [Google Scholar]
- 36. Steiner CF (2003) Variable dominance in pond communities: Assessing spatiotemporal variation in competition and predation intensity. Ecology 84: 982–990. [Google Scholar]
- 37.Jones SE, Lennon JT Tipping points in ecosystem stability triggered by spatial resource subsidies. In Review.
- 38. Cuthbert ID, del Giorgio P (1992) Toward a standard method of measuring color in freshwater. Limnol Oceanogr 37: 1319–1326. [Google Scholar]
- 39.Wetzel RG, Likens GE (2000) Limnological Analysis: Springer. 429 p. [Google Scholar]
- 40. Pace ML, Cole JJ (2002) Synchronous variation of dissolved organic carbon and color in lakes. Limnol Oceanogr 47: 333–342. [Google Scholar]
- 41.Neter J, Kutner M, Nachtsheim CJ, Wasserman W (1996) Applied Linear Statistical Models. Chicago: Irwin. 1408 p. [Google Scholar]
- 42.APHA (1998) Standard Methods for the Examination of Water and Wastewater; Association APH, editor. Washington, D.C.
- 43. Hamilton SK, Tank JL, Raikow DF, Wollheim WM, Peterson BJ, et al. (2001) Nitrogen uptake and transformation in a midwestern US stream: A stable isotope enrichment study. Biogeochemistry 54: 297–340. [Google Scholar]
- 44.Hamilton SK, Kurzman AL, Arango C, Jin LX, Robertson GP (2007) Evidence for carbon sequestration by agricultural liming. Glob Biogeochem Cycle 21.
- 45. Hamilton SK, Bruesewitz DA, Horst GP, Weed DB, Sarnelle O (2009) Biogenic calcite-phosphorus precipitation as a negative feedback to lake eutrophication. Can J Fish Aquat Sci 66: 343–350. [Google Scholar]
- 46. Grandy AS, Strickland MS, Lauber CL, Bradford MA, Fierer N (2009) The influence of microbial communities, management, and soil texture on soil organic matter chemistry. Geoderma 150: 278–286. [Google Scholar]
- 47.Oksanen J, Banchet FG, Kindt R, Legendre P, Minchin PR, et al. (2012) vegan: Community Ecology Package. R package version 2.0–5. Available: http://CRAN.R-project.org/package=vegan.
- 48. Fellman JB, Miller MP, Cory RM, D’Amore DV, White D (2009) Characterizing dissolved organic matter using PARAFAC modeling of fluorescence spectroscopy: a comparison of two models. Environ Sci Technol 43: 6228–6234. [DOI] [PubMed] [Google Scholar]
- 49. Cory RM, McKnight DM (2005) Fluorescence spectroscopy reveals ubiquitous presence of oxidized and reduced quinones in dissolved organic matter. Environ Sci Technol 39: 8142–8149. [DOI] [PubMed] [Google Scholar]
- 50. Stuiver M, Polach HA (1977) Reporting of 14C data - discussion. Radiocarbon 19: 355–363. [Google Scholar]
- 51. Kilham SS, Kreeger DA, Lynn SG, Goulden CE, Herrera L (1998) COMBO: a defined freshwater culture medium for algae and zooplankton. Hydrobiologia 377: 147–159. [Google Scholar]
- 52. Lennon JT, Smith VH, Williams K (2001) Influence of temperature on exotic Daphnia lumholtzi and implications for invasion success. J Plankton Res 23: 425–434. [Google Scholar]
- 53. Meyer JS, Ingersoll CG, McDonald LL, Boyce MS (1986) Estimating uncertainty in population growth rates: jackknife vs. bootstrap techniques. Ecology 67: 1156–1166. [Google Scholar]
- 54. Nielsen MV, Olsen Y (1989) The dependence of the assimilation efficiency in Daphnia magna on the 14C labeling period of the food alga Scenedesmus acutus . Limnol Oceanogr 34: 1311–1315. [Google Scholar]
- 55. Stemberger RS (1981) A general approach to the culture of planktonic rotifers. Can J Fish Aquat Sci 38: 721–724. [Google Scholar]
- 56. Lennon JT, Martiny JBH (2008) Rapid evolution buffers ecosystem impacts of viruses in a microbial food web. Ecol Lett 11: 1177–1188. [DOI] [PubMed] [Google Scholar]
- 57. Wang Q, Garrity GM, Tiedje JM, Cole JR (2007) Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol 73: 5261–5267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, et al. (2009) Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol 75: 7537–7541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kirchman DL (1993) Leucine incorporation as a measure of biomass production by heterotrophic bacteria. In: Kemp PF, Sherr BF, Sherr EB, Cole JJ, editors. Handbook of Methods in Aquatic Microbial Ecology. Boca Raton: Lewis Publishers. 509–512.
- 60.Briand E, Pringault O, Jacquet S, Torreton J (2004) The use of oxygen microprobes to measure bacterial respiration for determining bacterioplankton growth efficiency. Limnol Oceanogr Methods 2.
- 61. Cory RM, Miller MP, McKnight DM, Guerard JJ, Miller PL (2010) Effect of instrument-specific response on the analysis of fulvic acid fluorescence spectra. Limnol Oceanogr Methods 8: 67–78. [Google Scholar]
- 62. Curtis PJ, Schindler DW (1997) Hydrologic control of dissolved organic matter in low-order Precambrian Shield Lakes. Biogeochemistry 36: 125–138. [Google Scholar]
- 63. Stomp M, Huisman J, Stal LJ, Matthijs HCP (2007) Colorful niches of phototrophic microorganisms shaped by vibrations of the water molecule. ISME J 1: 271–282. [DOI] [PubMed] [Google Scholar]
- 64.Mulholland PJ (2003) Large-scale patterns in dissolved organic carbon concentrations, flux, and sources. In: Findlay SEG, Sinsabaugh RL, editors. Aquatic Ecosystems: Interactivity of Dissolved Organic Matter: Academic Press. 26–59.
- 65. Berggren M, Strom L, Laudon H, Karlsson J, Jonsson A, et al. (2010) Lake secondary production fueled by rapid transfer of low molecular weight organic carbon from terrestrial sources to aquatic consumers. Ecol Lett 13: 870–880. [DOI] [PubMed] [Google Scholar]
- 66. Jones SE, Newton RJ, McMahon KD (2009) Evidence for structuring of bacterial community composition by organic carbon source in temperate lakes. Environ Microbiol 11: 2463–2472. [DOI] [PubMed] [Google Scholar]
- 67. Russell JB (2007) The energy spilling reactions of bacteria and other organisms. J Mol Microbiol Biotechnol 13: 1–11. [DOI] [PubMed] [Google Scholar]
- 68. Cole JJ, Carpenter SR, Pace ML, Van de Bogert MC, Kitchell JL, et al. (2006) Differential support of lake food webs by three types of terrestrial organic carbon. Ecol Lett 9: 558–568. [DOI] [PubMed] [Google Scholar]
- 69. Richey JE, Wissmar RC, Devol AH, Likens GE, Eaton JS, et al. (1978) Carbon flow in 4 lake ecosystems: a structural approach. Science 202: 1183–1186. [DOI] [PubMed] [Google Scholar]
- 70.Wetzel RG (2001) Limnology: Lake and River Ecosystems. New York: Academic Press. 1006 p. [Google Scholar]
- 71. Jurgens K (1994) Impact of Daphnia on planktonic microbial food webs - a review. Mar Microb Food Webs 8: 295–324. [Google Scholar]
- 72. Brett MT, Kainz MJ, Taipale SJ, Seshan H (2009) Phytoplankton, not allochthonous carbon, sustains herbivorous zooplankton production. Proc Natl Acad Sci 106: 21197–21201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. von Wachenfeldt E, Bastviken D, Tranvik LJ (2009) Microbially induced flocculation of allochthonous dissolved organic carbon in lakes. Limnol Oceanogr 54: 1811–1818. [Google Scholar]
- 74.Tranvik LJ, von Wachenfeldt E (2009) Interactions of dissolved organic matter and humic substances. In: Likens GE, editor. Encyclopedia of Inland Waters. San Diego: Academic Press. 754–760.
- 75.Carpenter SR, Kitchell JF (1993) The Trophic Cascade in Lakes. Cambridge: Cambridge University Press.
- 76. Grey J, Jones RI, Sleep D (2001) Seasonal changes in the importance of the source of organic matter to the diet of zooplankton in Loch Ness, as indicated by stable isotope analysis. Limnol Oceanogr 46: 505–513. [Google Scholar]
- 77. McCallister SL, Bauer JE, Cherrier JE, Ducklow HW (2004) Assessing sources and ages of organic matter supporting river and estuarine bacterial production: A multiple-isotope (δ14C, δ13C, and δ15N) approach. Limnol Oceanogr 49: 1687–1702. [Google Scholar]
- 78. Caraco N, Bauer JE, Cole JJ, Petsch S, Raymond P (2010) Millennial-aged organic carbon subsidies to a modern river food web. Ecology 91: 2385–2393. [DOI] [PubMed] [Google Scholar]
- 79.Sterner RW, Elser JJ (2002) Ecological Stoichiometry. Princeton: Princeton University Press. 439 p. [Google Scholar]
- 80. Cleveland CC, Liptzin D (2007) C : N : P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 85: 235–252. [Google Scholar]
- 81.McKnight DM, Hood E, Klapper L (2003) Trace Organic Moieties of Dissolved Organic Material in Natural Waters. In: Findlay SE, Sinsabaugh RL, editors. Aquatic Ecosystems: Interactivity of Dissolved Organic Matter: Elsivier. 71–96.
- 82. Hood E, Williams MW, McKnight DM (2005) Sources of dissolved organic matter (DOM) in a Rocky Mountain stream using chemical fractionation and stable isotopes. Biogeochemistry 74: 231–255. [Google Scholar]
- 83. Talling JF (2010) Potassium - a non-limiting nutrient in fresh waters? Freshw Rev 3: 97–104. [Google Scholar]
- 84. Jaworski GHM, Talling JF, Heaney SI (2003) Potassium dependence and phytoplankton ecology: an experimental study. Freshwater Biol 48: 833–840. [Google Scholar]
- 85. Civitello DJ, Penczykowski RM, Hite JL, Duffy MA, Hall SR (2013) Potassium stimulates fungal epidemics in Daphnia by increasing host and parasite reproduction. Ecology 94: 380–388. [DOI] [PubMed] [Google Scholar]
- 86. Parker DL, Kumar HD, Rai LC, Singh JB (1997) Potassium salts inhibit growth of the cyanobacteria Microcystis spp. in pond water and defined media: Implications for control of microcystin-producing aquatic blooms. Appl Env Microbiol 63: 2324–2329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Lehman JT (1976) Ecological and nutritional studies on Dinobryon Ehrenb - seasonal periodicity and phosphate toxicity problem. Limnol Oceanogr 21: 646–658. [Google Scholar]
- 88.Sandgren CD (1988) The ecology of chrysophte flagellates: their growth and perennation strategies as freshwater phytoplankton. In: Sandgren CD, editor. Growth and Reproductive Strategies of Freshwater Phytoplankton. Cambridge: Cambridge University Press. 9–104.