
Figure 1.
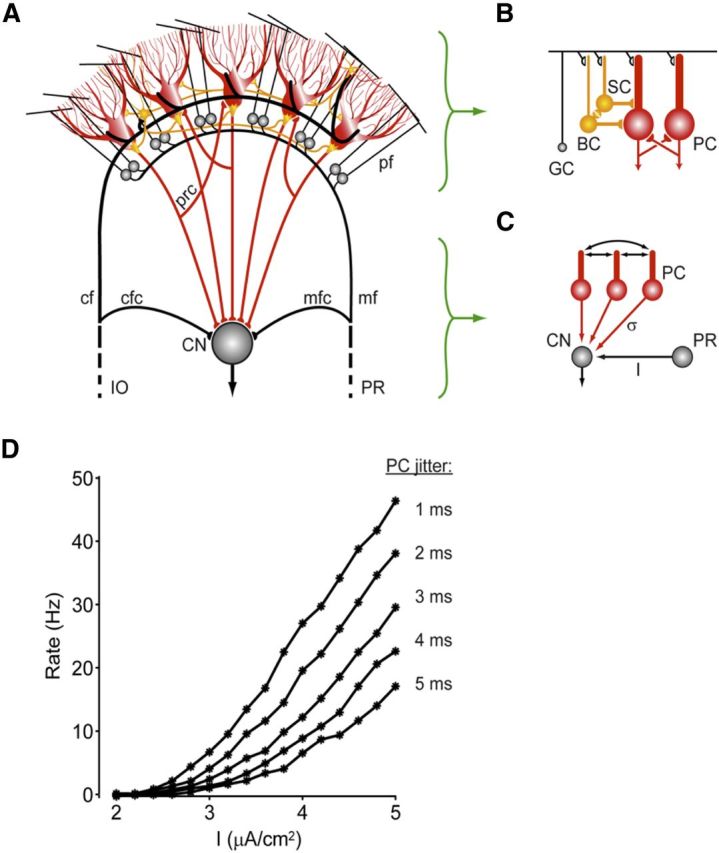
Scheme indicating how activity of Purkinje cells in the cerebellar cortex may influence activity in the cerebellar nuclei. A, The cerebellar cortex receives main excitatory inputs (black) from the inferior olive (IO) and pontine regions (PR) and provides via the Purkinje cell axons an inhibitory feedback (red) to the CN. Whereas the external IO and PR signals, which enter the cerebellum via the climbing fibers (cf) and the mossy fiber (mf)–parallel fiber (pf) pathway, are all excitatory, the local transmissions by the axons of the molecular layer interneurons (yellow) and recurrent collaterals (prc) of the Purkinje cells (red) are all inhibitory. mfc, Mossy fiber collaterals; cfc, climbing fiber collaterals. The local Golgi cell inhibition of granule cells (GC) is not depicted in this drawing. B, Molecular layer interneurons, that is, basket cells (BC) and stellate cells (SC), are coupled by gap junctions (yellow), whereas Purkinje cells (PC) can influence one another via recurrent collaterals. C, Firing rate gain in the CN after excitation (I) by the mossy fiber collaterals from the PR may be regulated by synchronous, high-frequency oscillations in the inhibitory activities of the Purkinje cells. The PC spike-time dispersion (σ) is inversely related to the synchrony of this network oscillation. D, The CN firing rate versus mfc input current (I) plots are shown for different values of the PC jitter. As the jitter is decreased from 5 to 1 ms (from bottom to top), the gain of the CN responses to the excitatory mfc inputs is dramatically increased. This interaction between mfc inputs and PC synchrony might be one of the potential mechanisms by which high-frequency oscillations in the cerebellar cortex exert their effects (modified from De Zeeuw et al., 2008).