

Abstract
Background and Aim. It is well known that androgen deprivation relates to penile fibrosis, so we hypothesize that long-term treatment with 5-alphareductase inhibitors (5ARIs) may increase the risk of fibrosis of prostate. Patients and Methods. Thirty-two BPH patients who underwent transurethral resection of the prostate were enrolled. The patients were divided into two groups: group one, 16 patients underwent TURP who had been treated with tamsulosin for 2 years; group two, 16 patients underwent TURP who had been treated with combination of tamsulosin and dutasteride for at least 1 year. We evaluated the expressions of nNOS, iNOS, eNOS, TGF-β1, TGF-β2, phosphorylated-Smad2/3 (p-Smad2/3), E-cadherin, N-cadherin, and α-smooth muscle actin in the resected prostate tissues by western blotting, and the TGF-β concentration was determined by ELISA kit. Results. The expressions of 3 isoforms of NOS were significantly increased in group 2 except of eNOS in lateral prostate, and the expressions of TGF-β1, TGF-β2, and p-Smad2/3 increased about 2-fold compared with group 1. In group 2, the E-cadherin expression decreased while N-cadherin expression increased significantly. Conclusions. The overexpression of nNOS may contribute to prostate smooth muscle relaxation; however, long-time treatment with 5 ARI increases the risk of fibrosis of prostate.
1. Introduction
The 5-alpha reductase (5 AR) inhibitor (5 ARI) was developed to treat the patients with symptomatic benign prostatic hyperplasia (BPH) and decrease the frequency and risk of BPH-related morbidities [1, 2]. Presently available orally administered 5 ARIs, finasteride and dutasteride, inhibit 5 AR, an enzyme that catalyzes the irreversible reduction of testosterone (T) to dihydrotestosterone (DHT), with NADPH as the hydrogen donor [3]. Dutasteride is a selective inhibitor of 5 AR type 1 (5 AR1) and 5 AR type 2 (5 AR2). Finasteride is considered mainly an inhibitor of 5 AR2 and is approximately 50 times weaker in inhibiting 5 AR1 than 5 AR2 [4]. The development of BPH is an androgen-dependent process, and androgen suppression causes regression primarily of the epithelial elements of the prostate, resulting in a reduction in the size of the gland and improvement in symptoms [5, 6].
As we know, androgen deprivation by surgical or medical castration is associated with penile cavernosal fibrosis resulting in penile tissue atrophy, alterations in dorsal nerve structure, alterations in endothelial morphology, reductions in trabecular smooth muscle content, increases in deposition of extracellular matrix, and increases in accumulation of adipocytes in the subtunical region of the corpus cavernosum [7]. Transforming growth factor-β (TGF-β) signaling pathway plays the most important role in the fibrotic process. TGF-β reduces collagenase production and stimulates the expression of tissue inhibitor of metalloproteinases, resulting in an overall inhibition of extracellular matrix (ECM) degradation and leading to excessive matrix accumulation [8, 9].
As androgen deprivation by surgical or medical castration induces cavernosal fibrosis, it may also affect the prostate tissue. In the present study, we investigated the effect of long-time treatment with 5 ARI on TGF-β signaling pathway in the prostate of BPH patients.
2. Patients and Methods
2.1. Patients
Total 32 BPH patients complaining of moderate-to-severe lower urinary tract symptoms (LUTS) who underwent transurethral resection of the prostate (TURP) were enrolled in the present study. The patients were divided into two groups: group 1, 16 patients underwent TURP who had been treated with alpha blocker (tamsulosin) for 2 years; group 2, the other 16 patients were treated with combination of alpha blocker (tamsulosin) and 5 ARI (dutasteride) for at least 1 year before the operation (Table 1).
Table 1.
Patients characteristics.
| Variants | Group 1 | Group 2 |
|---|---|---|
| (n = 16) | (n = 16) | |
| Age (years) | 68.9 ± 7.4 | 69.4 ± 6.9 |
|
| ||
| Preoperative PSA | 3.3 ± 1.92 | 4.8 ± 3.9 |
| Total prostate volume | 45.9 ± 18.7 | 48.2 ± 16.5 |
| T-zone volume | 21.4 ± 13.0 | 20.3 ± 12.1 |
| Resection volume | 19.8 ± 15.3 | 18.6 ± 16.6 |
PSA means prostate specific antigen. T-zone means transitional zone.
2.2. Tissue Preparation
Approximately 1 g of each lateral and anterior portion of the prostate tissue resected from each patient was obtained during the TURP. The tissues were immediately frozen at −80°C. For western blot analysis, the removed tissues were placed in protease-inhibitor buffer containing 1% Nonidet P-40, 50 mM HEPES, pH 7.5, 100 mM NaCl, 2 mM EDTA, 1 mM pyrophosphate, 10 mM sodium orthovanadate, 1 mM phenylmethylsulfonyl fluoride, and 100 mM sodium fluoride and were homogenized. The homogenate was centrifuged at 13,000 rpm for 30 min. The supernatant was collected and the volume was recorded.
2.3. Western Blotting
The samples were normalized for total protein content with a pierce BCA protein assay kit (Thermo Fisher Scientific, Rockford, IL, USA). 30 μg aliquots were suspended in an equal volume of 2× Laemmli buffer containing 2-mercaptoethanol and boiled for 5 min before performing SDS-PAGE. Proteins were transferred to Immobilon-P membranes (Millipore, Billerica, MA, USA), blocked with skim milk (5%), and incubated with various primary antibodies recognizing NOS (Santa Cruz Biotechnology, Santa Cruz, CA, USA), TGF-β (Santa Cruz Biotechnology, Santa Cruz, CA, USA), p-Smad2/3 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), Smad2/3 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), E-cadherin (Cell Signaling Technology, Danvers, MA, USA), N-cadherin (Cell Signaling Technology, Danvers, MA, USA), α-smooth muscle actin (Cell Signaling Technology, Danvers, MA, USA), and actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Antibody detection was accomplished via horseradish peroxidase-conjugated protein A or goat anti-rabbit immunoglobulin G (Santa Cruz Biotechnology, Santa Cruz, CA, USA), a chemiluminescent substrate (Millipore, Billerica, MA, USA), and exposure to X-ray film (Eastman Kodak Co., Rochester, NY, USA).
2.4. Human TGF-β Immunoassay
The samples were normalized for total protein content with a BCA kit and tested immediately or preserved at −80°C. TGF-β levels were analyzed using a commercially available ELISA (R&D Systems, Minneapolis, MN, USA) per the manufacturer's protocol. Briefly, this assay employs the quantitative sandwich enzyme immunoassay technique. A monoclonal antibody specific for TGF-β1 has been precoated onto a microplate. Standard, controls, and samples are pipetted into the wells and any TGF-β1 present is bound by the immobilized antibody. After washing away any unbound substances, an enzyme-linked polyclonal antibody specific for TGF-β1 is added to the wells to sandwich the TGF-β1 immobilized during the first incubation. Following a wash to remove any unbound antibody-enzyme reagent, a substrate solution is added to the wells and color develops in proportion to the amount of TGF-β1 bound in the initial step. The color development is stopped and the intensity of the color is measured.
2.5. Statistical Analyses
Data were presented as means ± SD. The statistical significances of the differences were calculated by one-way ANOVA followed by Bonferroni's multiple comparison. The values were compared using Student's paired t-test. A P value < 0.05 was considered statistically significant.
3. Results
3.1. The Expression of nNOS, iNOS, and eNOS
The expressions of nNOS and iNOS were significantly increased in group 2. However, the expression of eNOS in lateral portion of prostate was decreased (Figure 1, P < 0.05).
Figure 1.
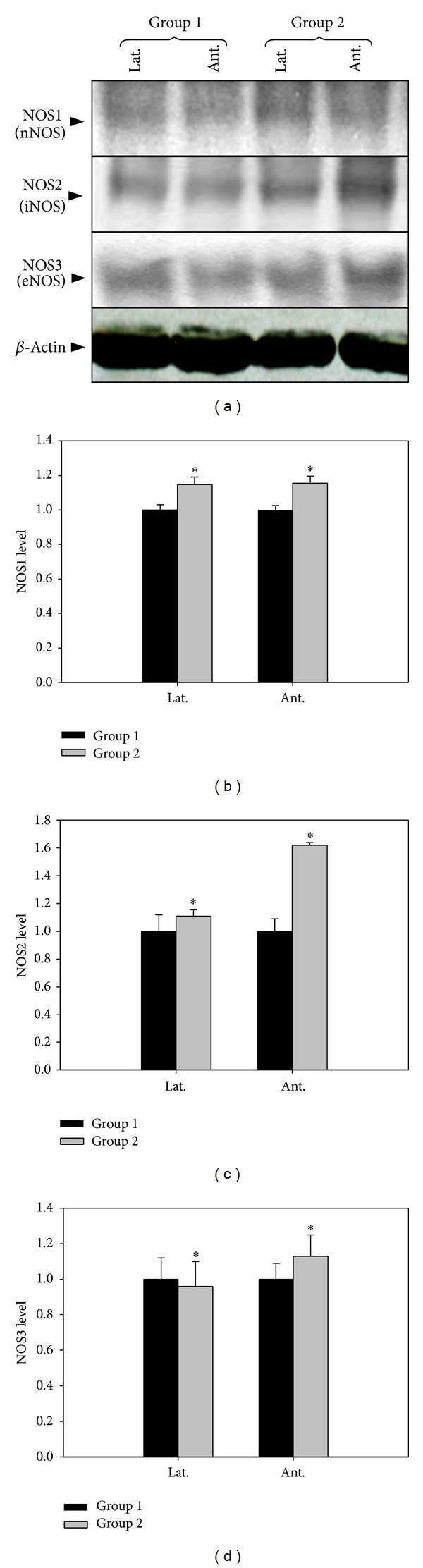
The effect of 5-alpha reductase inhibitor on expressions of nNOS, iNOS, and eNOS. nNOS: neuronal NO synthase, iNOS: inducible NO synthase, eNOS: endothelial NO synthase. Lat. means lateral portion of the prostate and Ant. means anterior wall. Each value is expressed as the mean ± SD of six independent experiments. *P < 0.05 versus control.
3.2. The Expression of TGF-β
Western blot of TGF-β was shown in Figure 2. The upregulated expressions of TGF-β1 and TGF-β2 were observed in group 2. The result showed that lateral portion of prostate expressed more TGF-β1, TGF-β2 than anterior wall. P-Smad2/3 expression was also increased in group 2, but there was no difference between the lateral portion and anterior wall (Figure 2, P < 0.05).
Figure 2.
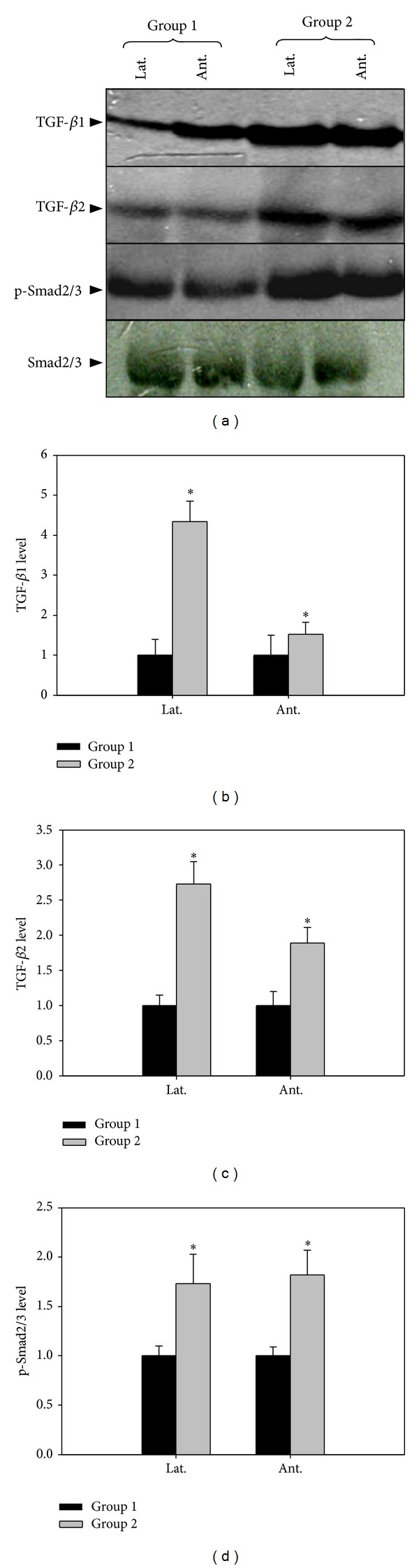
The effect of 5-alpha reductase inhibitor on expression of TGF-β1, TGF-β2, and p-Smad2/3. Lat. means lateral portion of the prostate and Ant. means anterior wall. Each value is expressed as the mean ± SD of six independent experiments. *P < 0.05 versus control.
The TGF-β level was determined by ELISA, and the results indicated that TGF-β was significantly increased in group 2 (Figure 3, P < 0.05).
Figure 3.
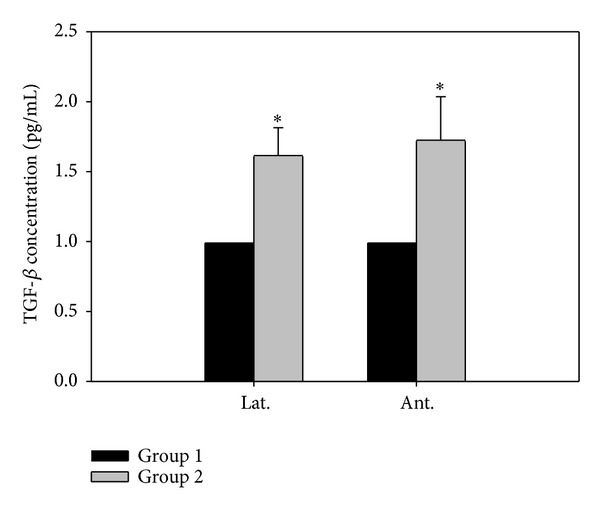
Human TGF-β Immunoassay of prostate. Lat. means lateral portion of the prostate and Ant. means anterior wall. Each value is expressed as the mean ± SD of six independent experiments. *P < 0.05 versus control.
3.3. The Expression of E-Cadherin and N-Cadherin
The western blotting results of E-cadherin and N-cadherin were shown in Figure 4. The E-cadherin level decreased and N-cadherin increased significantly in group 2. Alpha smooth muscle actin was markedly increased in group 2 and expressed more in lateral portion of the prostate.
Figure 4.
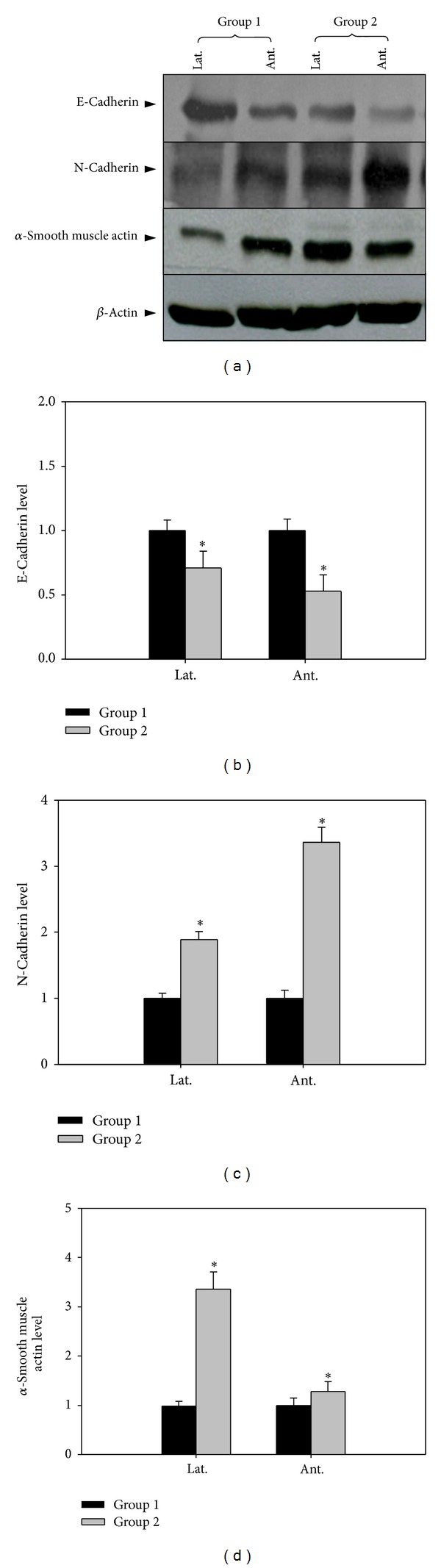
The effect of 5-alpha reductase inhibitor on expression of E, N-cadherin (Fibrosis). Lat. means lateral portion of the prostate and Ant. means anterior wall. Each value is expressed as the mean ± SD of six independent experiments. *P < 0.05 versus control.
4. Discussion
TGF-β participates in the pathogenesis of multiple cardiovascular diseases, including hypertension, atherosclerosis, cardiac hypertrophy, and heart failure [10–12]. Also, it has been shown to be a regulator of stromal proliferation and differentiation in the prostate [13]. It is well known that androgen plays an important role in the pathogenesis of BPH. The action of androgen is mediated through actions of a host of soluble growth factors, among which TGF-β is the most versatile in its ability to regulate proliferation, growth arrest, differentiation, and apoptosis of prostatic stromal cells [14]. It has been demonstrated that the effect of 5 ARI on BPH was associated with suppression of the insulin like growth factor 1 gene and with an increase in TGF-β gene expression [15]. TGF-β predominantly transmits the signals through cytoplasmic proteins called Smads, which translocates into the cell nucleus acting as transcription factors. Eight different members of the Smad family have been identified in mammals, in which Smad2 and Smad3 are specific mediators of TGF-β/activin pathways [10, 11]. In this study, the results showed that long-term treatment with 5 ARI significantly increased the expression of TGF-β1, TGF-β2, and p-Samd2/3 compared with the control group, suggesting 5 ARI-induced cell death was regulated by TGF-β-Smad pathway. Decreased expression of E-cadherin accompanied by increased expression of N-cadherin resulted in a loss of epithelial characteristics and indicated that the epithelial cell underwent epithelial to mesenchymal transition (EMT).
TGF-β-Smad pathway is very important stream also involved in fibrotic process. In vascular smooth muscle cells, endothelial cells, and fibroblasts, TGF-β1 increases the synthesis of ECM proteins, such as fibronectin, collagens, and activator inhibitor-1 (PAI-1), even at low concentrations [8, 9]. TGF-β induces expression of fibronectin which is required for enhancement of α-smooth muscle actin and collagen type I expression [8]. PAI-1 is a serpin class protease inhibitor, important in tissue remodeling by modulating thrombosis, inflammation, migration, and ECM.
TGF-β reduces collagenase production and stimulates the expression of tissue inhibitor of metalloproteinases (TIMP), resulting in an overall inhibition of ECM degradation and leading to excessive matrix accumulation [8, 9]. The mechanisms involved in TGF-β mediated vascular fibrosis are complex, including activation of Smad proteins, protein kinases, production of mediators, and crosstalk between pathways. TGF-β also acts as a mediator of vascular fibrosis induced by several agents involved in cardiovascular diseases, including mechanical stress, angiotensin II, high glucose, and advanced glycation products [16–18]. In Peyronie's disease, the fibrosis is characterized by an increase in collagen over the intracellular compartment and is associated with the production of profibrotic factors, such as TGF-β1, PAI-1, and reactive oxygen species during oxidative stress [19–21]. This fibrosis is accompanied by the induction of iNOS, which acts as an endogenous antifibrotic mechanism in response to the profibrotic processes [19, 22].
The presence of NOS has been demonstrated in the human prostate that the eNOS is related to the maintenance of local vascular perfusion, whereas the nNOS is mainly involved in the control of smooth muscle tone and glandular function, including proliferation of epithelial and subepithelial cells [23, 24]. Until today, the inducible form of NOS, the iNOS, has not been found in normal prostate tissue; however, there are hints that the enzyme is expressed in hyperplastic and malignant tissue [25, 26]. The reduction in glandular tissue by 5 ARI is achieved by the induction of apoptosis, which is histologically manifested by ductal atrophy. Inhibition also diminishes the number of blood vessels in the prostate because of a reduction in vascular-derived endothelial growth factor which downregulates the eNOS expression in lateral portion of prostate [27].
5. Conclusions
The overexpression of nNOS may contribute to prostate smooth muscle relaxation and improve the symptoms; however, long-time treatment with 5 ARI increases the risk of fibrosis by the TGF-β-p-Smad signaling pathway.
What is Known?
The 5 ARIs have been successfully used for treatment of BPH by inhibiting the conversion of T to DHT, resulting in a reduction in the size of the gland and improvement in symptoms.
Androgen deprivation by surgical or medical castration is associated with penile cavernosal fibrosis.
What is New?
Androgen deprivation induced by long-term treatment with 5 ARI increases the risk of fibrosis via the TGF-β-p-Smad signaling pathway.
The upregulated nNOS expression may contribute to prostate smooth muscle relaxation and improve the symptoms.
Conflict of Interests
The authors do not have conflict of interests to declare and also do not have anything to declare about Santa Cruz Biotechnology, Thermo Fisher Scientific, Cell Signaling Technology, and Eastman Kodak Co.
Authors' Contribution
Hye Kyung Kim, Chen Zhao, Bo Ram Choi, Han Jung Chae, Do Sung Kim, and Jong Kwan Park were involved in the laboratory procedures, data collection, and analysis. Hye Kyung Kim and Jong Kwan Park were involved in the preparation of the paper and the review of the paper.
Acknowledgment
This study was supported by Grants from the Korea Healthcare Technology R&D Project, Ministry for Health, Welfare & Family Affairs, Republic of Korea (HI13C0104).
References
- 1.McConnell JD, Bruskewitz R, Walsh P, et al. The effect of finasteride on the risk of acute urinary retention and the need for surgical treatment among men with benign prostatic hyperplasia. The New England Journal of Medicine. 1998;338(9):557–563. doi: 10.1056/NEJM199802263380901. [DOI] [PubMed] [Google Scholar]
- 2.Roehrborn CG, Boyle P, Nickel JC, Hoefner K, Andriole G. Efficacy and safety of a dual inhibitor of 5-alpha-reductase types 1 and 2 (dutasteride) in men with benign prostatic hyperplasia. Urology. 2002;60(3):434–441. doi: 10.1016/s0090-4295(02)01905-2. [DOI] [PubMed] [Google Scholar]
- 3.Russell DW, Wilson JD. Steroid 5α-reductase: two genes/two enzymes. Annual Review of Biochemistry. 1994;63:25–61. doi: 10.1146/annurev.bi.63.070194.000325. [DOI] [PubMed] [Google Scholar]
- 4.Aggarwal S, Thareja S, Verma A, Bhardwaj TR, Kumar M. An overview on 5α-reductase inhibitors. Steroids. 2010;75(2):109–153. doi: 10.1016/j.steroids.2009.10.005. [DOI] [PubMed] [Google Scholar]
- 5.Coffey DS, Walsh PC. Clinical and experimental studies of benign prostatic hyperplasia. Urologic Clinics of North America. 1990;17(3):461–475. [PubMed] [Google Scholar]
- 6.Peters CA, Walsh PC. The effect of nafarelin, acetate, a luteinizing-hormone-releasing hormone agonist, on benign prostatic hyperplasia. The New England Journal of Medicine. 1987;317(10):599–604. doi: 10.1056/NEJM198709033171004. [DOI] [PubMed] [Google Scholar]
- 7.El-Sakka AI, Yassin AA. Review: amelioration of penile fibrosis: myth or reality. Journal of Andrology. 2010;31(4):324–335. doi: 10.2164/jandrol.109.008730. [DOI] [PubMed] [Google Scholar]
- 8.Leask A, Abraham DJ. TGF-β signaling and the fibrotic response. The FASEB Journal. 2004;18(7):816–827. doi: 10.1096/fj.03-1273rev. [DOI] [PubMed] [Google Scholar]
- 9.Verrecchia F, Mauviel A. Transforming growth factor-β signaling through the Smad pathway: role in extracellular matrix gene expression and regulation. Journal of Investigative Dermatology. 2002;118(2):211–215. doi: 10.1046/j.1523-1747.2002.01641.x. [DOI] [PubMed] [Google Scholar]
- 10.Massagué J, Seoane J, Wotton D. Smad transcription factors. Genes and Development. 2005;19(23):2783–2810. doi: 10.1101/gad.1350705. [DOI] [PubMed] [Google Scholar]
- 11.Javelaud D, Mauviel A. Mammalian transforming growth factor-βs: smad signaling and physio-pathological roles. International Journal of Biochemistry and Cell Biology. 2004;36(7):1161–1165. doi: 10.1016/S1357-2725(03)00255-3. [DOI] [PubMed] [Google Scholar]
- 12.Moustakas A, Pardali K, Gaal A, Heldin C-H. Mechanisms of TGF-β signaling in regulation of cell growth and differentiation. Immunology Letters. 2002;82(1-2):85–91. doi: 10.1016/s0165-2478(02)00023-8. [DOI] [PubMed] [Google Scholar]
- 13.Huang X, Lee C. Regulation of stromal proliferation, growth arrest, differentiation and apoptosis in benign prostatic hyperplasia by TGF-β. Frontiers in Bioscience. 2003;8:s740–s749. doi: 10.2741/1093. [DOI] [PubMed] [Google Scholar]
- 14.Timme TL, Truong LD, Slawin KM, Kadmon D, Sang Hee Park SHP, Thompson TC. Mesenchymal-epithelial interactions and transforming growth factor-β1 expression during normal and abnormal prostatic growth. Microscopy Research and Technique. 1995;30(4):333–341. doi: 10.1002/jemt.1070300408. [DOI] [PubMed] [Google Scholar]
- 15.Wu S-F, Sun H-Z, Qi X-D, Tu Z-H. Effect of epristeride on the expression of IGF-1 and TGF-β receptors in androgen-induced castrated rat prostate. Experimental Biology and Medicine. 2001;226(10):954–960. doi: 10.1177/153537020122601012. [DOI] [PubMed] [Google Scholar]
- 16.Ruiz-Ortega M, Ruperez M, Esteban V, Egido J. Molecular mechanisms of angiotensin II-induced vascular injury. Current Hypertension Reports. 2003;5(1):73–79. doi: 10.1007/s11906-003-0014-0. [DOI] [PubMed] [Google Scholar]
- 17.Li JH, Huang XR, Zhu H-J, Johnson R, Lan HY. Role of TGF-β signaling in extracellular matrix production under high glucose conditions. Kidney International. 2003;63(6):2010–2019. doi: 10.1046/j.1523-1755.2003.00016.x. [DOI] [PubMed] [Google Scholar]
- 18.Li JH, Huang XR, Zhu H-J, et al. Advanced glycation end products activate Smad signaling via TGF-beta-dependent and independent mechanisms: implications for diabetic renal and vascular disease. The FASEB Journal. 2004;18(1):176–178. doi: 10.1096/fj.02-1117fje. [DOI] [PubMed] [Google Scholar]
- 19.Gonzalez-Cadavid NF, Rajfer J. Mechanisms of disease: new insights into the cellular and molecular pathology of Peyronie’s disease. Nature Clinical Practice Urology. 2005;2(6):291–297. doi: 10.1038/ncpuro0201. [DOI] [PubMed] [Google Scholar]
- 20.El-Sakka AI, Hassoba HM, Pillarisetty RJ, Dahiya R, Lue TF. Peyronie’s disease is associated with an increase in transforming growth factor-β protein expression. Journal of Urology. 1997;158(4):1391–1394. [PubMed] [Google Scholar]
- 21.Bivalacqua TJ, Diner EK, Novak TE, et al. A rat model of Peyronie’s disease associated with a decrease in erectile activity and an increase in inducible nitric oxide synthase protein expression. Journal of Urology. 2000;163(6):1992–1998. [PubMed] [Google Scholar]
- 22.Davila HH, Magee TR, Vernet D, Rajfer J, Gonzalez-Cadavid NF. Gene transfer of inducible nitric oxide synthase complementary DNA regresses the fibrotic plaque in an animal model of Peyronie’s disease. Biology of Reproduction. 2004;71(5):1568–1577. doi: 10.1095/biolreprod.104.030833. [DOI] [PubMed] [Google Scholar]
- 23.Burnett AL, Maguire MP, Chamness SL, et al. Characterization and localization of nitric oxide synthase in the human prostate. Urology. 1995;45(3):435–439. doi: 10.1016/S0090-4295(99)80012-0. [DOI] [PubMed] [Google Scholar]
- 24.Gradini R, Realacci M, Ginepri A, et al. Nitric oxide synthases in normal and benign hyperplastic human prostate: immunohistochemistry and molecular biology. The Journal of Pathology. 1999;189:224–229. doi: 10.1002/(SICI)1096-9896(199910)189:2<224::AID-PATH422>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 25.Klotz T, Bloch W, Volberg C, Engelmann U, Addicks K. Selective expression of inducible nitric oxide synthase in human prostate carcinoma. Cancer. 1998;82:1897–1903. [PubMed] [Google Scholar]
- 26.Baltaci S, Orhan D, Gögüs Ç, Türkölmez K, Tulunay Ö, Gögüs O. Inducible nitric oxide synthase expression in benign prostatic hyperplasia, low- and high-grade prostatic intraepithelial neoplasia and prostatic carcinoma. BJU International. 2001;88(1):100–103. doi: 10.1046/j.1464-410x.2001.02231.x. [DOI] [PubMed] [Google Scholar]
- 27.Marks LS, Partin AW, Dorey FJ, et al. Long-term effects of finasteride on prostate tissue composition. Urology. 1999;53(3):574–580. [PubMed] [Google Scholar]