
Fig. 1.
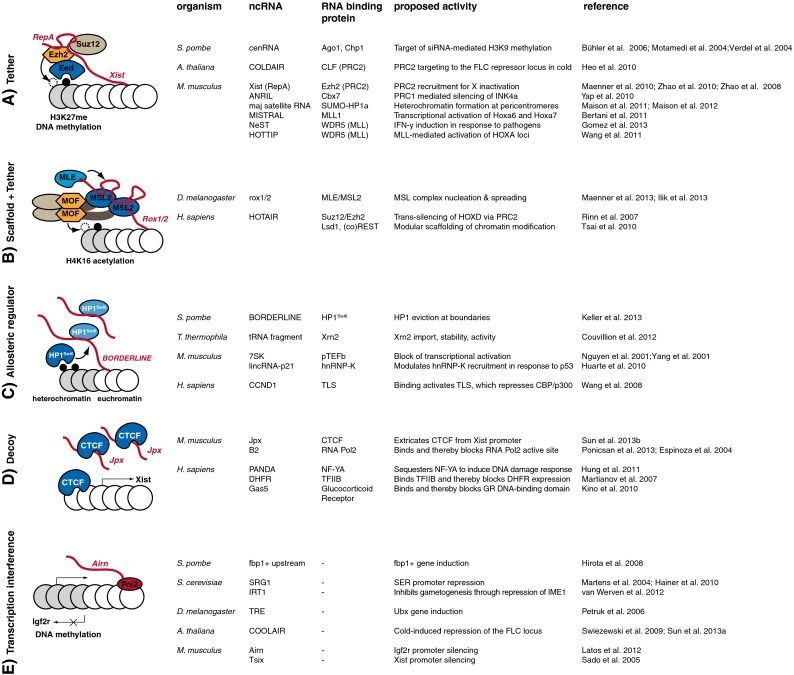
Various chromatin-associated activities of long ncRNAs. (A) ncRNA tethers link activities in cis via a nascent transcript. For example, the ncRNA Xist is transcribed from the X-chromosome and interacts via a conserved stem-loop sequence termed RepA with the PRC2 complex members Ezh2 and Suz12. This triggers H3K27 methylation, which is recognized by the H3K27 reader Eed. This leads to inactivation of the X chromosome. (B) In addition to acting as tethers, some long ncRNAs may also act as scaffolds. The ncRNA (e.g. rox1/2) not only serves as a nucleation site but is also an integral part of the chromatin-modifying activity (e.g. the MSL complex). In the case of the Drosophila MSL complex, this leads to H4K16 acetylation via the histone acetyltransferase MOF and transcriptional upregulation on the male X chromosome. (C) ncRNAs acting as allosteric regulators affect chromatin effectors by direct binding and tuning of their molecular properties. For example, RNA binding to the hinge region of HP1Swi6 induces conformational changes in the HP1Swi6 CD, which is incompatible with stable H3K9me3 association. This leads ultimately to HP1Swi6 eviction from heterochromatin. (D) Decoys mimicking a natural ligand of the molecule to be regulated compete for the binding site. For example, Jpx RNA contains a motif that effectively competes with CTCF DNA binding. This extricates CTCF from the Xist promoter and thereby activates Xist transcription, inducing X chromosome inactivation in female mice. (E) In the transcriptional interference model, the action of the transcribing RNA polymerase rather than the RNA product is functionally relevant. One example is transcriptional overlap of the Airn ncRNA, which is required for Igf2r promoter silencing. The table lists representative examples of each mode of action