

Abstract
Why does performing certain tasks cause the aversive experience of mental effort and concomitant deterioration in task performance? One explanation posits a physical resource that is depleted over time. We propose an alternate explanation that centers on mental representations of the costs and benefits associated with task performance. Specifically, certain computational mechanisms, especially those associated with executive function, can be deployed for only a limited number of simultaneous tasks at any given moment. Consequently, the deployment of these computational mechanisms carries an opportunity cost – that is, the next-best use to which these systems might be put. We argue that the phenomenology of effort can be understood as the felt output of these cost/benefit computations. In turn, the subjective experience of effort motivates reduced deployment of these computational mechanisms in the service of the present task. These opportunity cost representations, then, together with other cost/benefit calculations, determine effort expended and, everything else equal, result in performance reductions. In making our case for this position, we review alternate explanations both for the phenomenology of effort associated with these tasks and for performance reductions over time. Likewise, we review the broad range of relevant empirical results from across subdisciplines, especially psychology and neuroscience. We hope that our proposal will help to build links among the diverse fields that have been addressing similar questions from different perspectives, and we emphasize ways in which alternate models might be empirically distinguished.
Keywords: evolutionary psychology, mental effort, neuroeconomics, phenomenology, self-control
I have no expectation that the laws of mental fatigue will be formulated in the immediate future.
— Dodge (1917, p. 89)
Remarkably, given that fatigue has been studied formally for well over 100 years, there is still no scientifically mature theory of its origins and functions.
— Hockey (2011, p. 167)
1. Introduction
For some of the brain’s functions, such as the regulation of body temperature and heart rate, performance is maintained without noticeable impairment over time. Similarly, the visual system executes its functions, from the retina to V1 to object recognition systems, and so on, more or less continuously during waking hours. The operation of these systems carries no phenomenology of effort, and performance reductions, if any, are slight. These observations imply that at least some of the brain’s functions can continue over sustained periods with minimal reduction in performance and without any conscious sensation of effort. In contrast, other mental tasks (e.g., scanning a display for infrequent, subtle signals; doing mental arithmetic, etc.) give rise to the conscious sensation of effort and seem difficult to execute continuously over time (Ackerman 2011).
Why are some, but not all, mental operations performed without the sensation of effort and without performance loss? Our goal here is to sketch a computational explanation for both the subjective phenomenology of mental effort and the associated behavioral performance reductions. Our interest ranges broadly, from tasks such as the Stroop (Webb & Sheeran 2003) to math problems (Arai 1912) and complex decision making (Masicampo & Baumeister 2008; Vohs et al. 2008). We propose that both phenomenology and performance in these mental tasks rest on a common foundation: computations of their benefits and costs relative to other operations to which the same processes might be applied. Subjective effort, on this view, is the conscious, experienced measurement of the cost – especially the opportunity cost – of continuing the task. The subjective experience of mental effort, which is generally aversive, in turn motivates reallocation of computational processes to relatively more valuable tasks. Our explanation contrasts with proposals that attribute performance reductions to depletion of a resource or to “willpower” (e.g., Gailliot & Baumeister 2007).
1.1. Phenomena to be explained
In one of the earliest studies of mental effort, Arai (1912) practiced multiplying pairs of four-digit numbers in her head until, after several months, she had reached a plateau in performance. She then completed a 4-day marathon of solving multiplication problems continuously, 12 hours per day, observing that it took her longer to solve problems over each successive day’s session and concluding that “difficult and disagreeable continued work brings about a decrease in the efficiency of the function exercised” (p. 114). Huxtable et al. (1946) replicated Arai’s experiment with three graduate student participants. Performance decrements over the course of each day were measurable but slight in magnitude and not as consistent as participants’ reports of extreme weariness, restlessness, and boredom. In retrospect, one participant commented that she “[w]ould not repeat these 4 days for $10,000” (p. 52).
1.1.1. Within-task performance reductions and associated phenomenology
More recently, performance in “vigilance tasks,” which require monitoring visual displays or auditory streams for infrequent signals (e.g., Mackworth 1948), has been shown to reliably decrease over time, with concomitant increases in perceived mental effort (Scerbo 2001; Warm et al. 2008). Likewise, after long periods of time in flight simulators, pilots are more easily distracted by noncritical signals and less able to detect critical signals (Warm et al. 2008). Ratings of boredom in vigilance tasks increase rapidly above pre-task levels typically (Scerbo & Holcomb 1993), but the increase in boredom can be delayed by minor variations in task parameters, such as increasing stimulus variety (Scerbo 2001).
Performance reductions have also been observed in a variety of other tasks that require sustained attention – for example, in “flanker’ tasks, in which subjects are asked to respond to a central target stimulus (e.g., by indicating the direction of an arrow) while ignoring adjacent stimuli with incongruent information (e.g., arrows that point in directions opposite from the target arrow) that are making the task more difficult. In one version of the task, where the central target is a letter and flanking stimuli are other letters, performance generally worsens after 90 min. (Lorist et al. 2005). Likewise, performance (as measured by reaction time and accuracy) decreases over time in “task-switching” paradigms, in which subjects are asked to respond to different features of the stimulus (e.g., the color, the size) depending on the trial (Lorist et al. 2000). Similarly, in a data entry task intended to induce fatigue, Healy et al. (2004) found that accuracy declined over time.
Broadly, tasks that engage executive functions show performance decrements over time (Holding 1983; van der Linden et al. 2003). Notably, rewards improve performance in executive function tasks (e.g., Krebs et al. 2010), suggesting that performance reductions are not mandatory, as one might expect if reductions were due to processes akin to mechanical breakdowns.
1.1.2. Between-task performance reductions and associated phenomenology
A separate experimental literature shows that performance reductions also occur when subjects perform two different tasks in sequence. In a typical experiment, subjects in the experimental condition are asked to perform a first task (sometimes referred to as the “depleting” task, although so naming the task prejudges the issue) that is assumed to require volitional control of attention, emotion, behavior, or cognition, and immediately thereafter, a second task (sometimes referred to as the “dependent” task) that is assumed to require volitional control in a different domain. Subjects in the control condition typically perform an “easy” version of the experimental first task and the same dependent task as the experimental group. This dual-task paradigm (Baumeister et al. 1998) is generally used to test the prediction that performing the so-called depleting task will reduce performance on the second.
A recent meta-analysis by Hagger et al. (2010a) identified 83 published experimental studies that included 198 independent tests of this effect. The overall effect size for performance impairment in the dependent task as a function of condition was medium-to-large (d=0.62, p<.001), with substantial heterogeneity across studies (I2=35%). The same meta-analysis found that in such studies, participants rate the experimental task as more demanding than the control task, with medium-to-large effect sizes on self-reported effort (d=0.64), perceived difficulty (d=0.94), and self-reported fatigue (d=0.44) (Hagger et al. 2010a). In contrast, other dimensions of subjective experience, including positive affect (d=−0.03) and negative affect (d=0.14), are minimally changed in such experiments (Hagger et al. 2010a).
As with within-task studies, manipulating participants’ motivation (e.g., providing incentives for performance) can attenuate or eliminate performance decrements in dual-task studies. Hagger et al. (2010a) found that in three studies comprising 10 independent tests of the effect of motivational strategies on performance in dual-task experiments, the effect size for the interaction was d=1.05.
1.2. Outline
To explain the above patterns surrounding the phenomenology of effort and concomitant reductions in task performance, we proceed as follows: In the first part of section 2, we describe key assumptions underlying our model: that the mechanisms that comprise the mind have evolved functions, that some version of the computational theory of mind is true, and that subjective experience can be understood as functioning to motivate adaptive behavior.
Next we describe the adaptive problem of simultaneity and its general solution, prioritization. We argue that certain mental processes can be flexibly deployed to multiple purposes – but not all at the same time. Choosing to do one thing with such a mental process necessarily requires choosing not to do another, and making such trade-offs optimally entails prioritizing options of greatest net value. We propose that the conscious experience of mental effort indexes opportunity costs, motivating the reallocation of computational processes toward the best alternative. We also link our account with similar, previous proposals.
In section 3, we discuss alternate accounts for both the phenomenology of effort and reductions in task performance, highlighting some potential difficulties with these models and articulating predictions that follow from our account that diverge from those made by alternate accounts. In section 4, we review empirical findings from neuroscience, especially regarding brain metabolism and representations of value, which collectively raise doubts about alternate explanations but are consistent with our view.
The final section summarizes and concludes.
2. Our model: Mental effort as opportunity cost computation
2.1. Assumptions
Our argument rests on three basic assumptions. First, we assume the brain is functionally organized to generate adaptive behavior. Because evolution by natural selection is the only known natural explanation for complex functional organization, we assume that all aspects of biological design, including the human brain, have an explanation in terms of evolved function (Pinker 1997; Tooby & Cosmides 1992). We note that this assumption does not commit us to the view that all behavior is adaptive (Symons 1992), to the position that all traits are adaptations, or to the view that the mind is optimally designed. Among other reasons, systems designed for ancestral environments can have positive or negative effects in modern environments, because our contemporary circumstances differ in any number of ways from those of our evolutionary ancestors (Burnham & Phelan 2000). Likewise, an adaptation that promotes functional behavior in most situations can in certain situations generate dysfunctional behavior.
Second, we assume that some version of the computational theory of mind is true (see Pinker 1997). That is, we embrace the view that the mind is an information-processing system. Understanding these computations – including the functions they serve and the details of the way the brain implements these functions – is required for explaining behavior.
Third, we assume that subjective experience can be understood computationally as motivating the organism to behave adaptively (Lazarus 1993; Tooby et al. 2008). We reject the view, occasionally referred to as “naïve realism,” that the external (or internal) world is directly and veridically experienced (Brain 1951). Instead, we suggest that qualia are the experiential component of computational outputs or measurements, information that serves a function in the context of decision making (Damasio 1999). For example, the emotion of jealousy can be understood as indexing the potential loss of a valued relationship, motivating actions to reduce the likelihood of such loss (Buss et al. 1992). Another example is the sensation of hunger. Hunger is a mental representation of the body’s current caloric needs, integrating signals from organs in the periphery and the stomach, and, in virtue of those needs, the present marginal value of eating. This computation gives rise to the conscious sensation we label “hunger,” motivating appropriate behavior toward food. (For two excellent reviews, see Barsh & Schwartz [2002] and Grill & Kaplan [2002].)
Because we take these three ideas to be our assumptions, we do not defend them here. Instead, we draw on them to consider the puzzle of mental effort. Specifically, given that many tasks associated with feelings of mental effort seem to have good outcomes – working hard yields professional success, resisting chocolate leads to good health – one might have supposed that engaging in such tasks would generate positive, rather than negative, sensations. Why, if revising a manuscript contributes to the achievement of key long-term goals, does it feel aversively “effortful”? What might the sensation of effort be measuring, and what adaptive outcomes might it be designed to bring about?
2.2. Adaptive problem: Simultaneity
In this section, we sketch the basics of our model. Following the usual process in adaptationist analysis (Williams 1966), we begin by specifying the adaptive problem that we believe the computational mechanisms in question might be designed to solve (Tooby & Cosmides 1992). We then address the computations, along with inputs and outputs, that might be able to solve the problem we identify (see also Marr 1982; Pinker 1997). Subsequent to this analysis, we review the existing data and how our proposal might explain previous results.
At the most general level, the adaptive problem we believe to be at stake here is the problem of simultaneity – not everything can be done at once – and the concomitant solution of prioritization – that is, choosing what to do at the expense of other options. In the context of behavior, one cannot work toward multiple goals at the same time to the extent that there are incompatibilities in reaching those goals.
Simultaneity is a problem that confronts any system designed to accomplish multiple goals. In the mechanical (as opposed to computational) domain, the problem is clear in cases such as ducking versus jumping. Doing one precludes the other. We hasten to add that some goals can be advanced simultaneously. For instance, fleeing from a predator might well accomplish an immediate survival goal, and at the same time have beneficial effects on cardiovascular health. The problems of simultaneity and prioritization depend on the tasks in question and the processes required for their execution.
2.3. General solution: Prioritization
The solution to the problem of simultaneity is prioritization. For example, with a sprained ankle, prioritizing rest is sensible when there is no pressing need, such as escaping the presence of a predator. But if a predator is present, the cost/benefit computations change, and resting the ankle (reducing the chance of continued damage) is less important than is using it to flee. Decision making in this respect is in part driven by a weighing of the motivational outputs – the pain of putting weight on the ankle set against the fear of a predator, which motivates fleeing.
The problem of prioritization exists for mental operations, as well. The mind accomplishes many tasks at the same time because there are a large number of mechanisms that act in parallel (Alexander & Crutcher 1990; Evans 2008; Fodor 1983; Minsky 1985; Nassi & Callaway 2009; Rousselet et al. 2002; Rummelhart et al. 1986; Sigman & Dehaene 2008; Sperber 1994). To the extent that two different tasks require the same computational mechanisms, they cannot both be accomplished simultaneously with uncompromised effectiveness. Consider decisions about where to direct one’s gaze. The rich, high-resolution perceptual apparatus in the fovea is finite, and it cannot be used at the same time for the entire visual field. The eyes must be directed somewhere, and foveating one part of the world necessarily precludes foveating other parts of the visual scene. The fovea and the computational apparatus downstream of it cannot simultaneously be applied to everything.
Working memory is similarly constrained in a way that mirrors the deployment of the fovea. A limited number of data structures can be actively maintained in working memory at any given time (Evans 2008; Miller 1956; Miller & Cohen 2001), leading naturally to the necessity of decisions about what gets maintained. Given the problem of simultaneity, a means is needed to evaluate the value of using computational systems such as working memory for mutually exclusive tasks. To return to the example above, attending to what is in the visual array might reduce processing of information in the auditory stream. Limited attention, in this sense, can be thought of as a trade-off in extracting information between these two information channels.
These considerations locate the solution to the adaptive problem of simultaneity in prioritizing among possible computations – that is, identifying which of the possible actions or computations ought to be performed. In turn, solving the problem of prioritization, very generally, requires the assignment of costs and benefits to candidate options. In the context of computations, this means, of course, computing the costs and benefits of candidate computations and then comparing these.
A computational challenge for making these trade-offs is that costs and benefits come in many different currencies. From a functional standpoint, the value of an act depended on its eventual net contribution to fitness. Computational mechanisms, of course, cannot directly compute fitness outcomes, so they must use proxy variables, evaluating the benefits of possibilities in terms of local variables (Symons 1992). That is, the design of these mechanisms can be understood in the context of evolutionary selection for systems that assigned weights in a way that maximized reproductive success (Glimcher 2003).
2.3.1. Specific solution: Prioritization using opportunity costs
The problem of simultaneity is illustrated by foraging organisms, which can feed in only one patch at any given time and, therefore, must decide when to stay in their current patch and when to leave it in search of a new one (Charnov 1976). Feeding at the current patch carries opportunity costs – that is, the value of the next-best alternative to the current choice. When foraging organisms change location, they do so because the rate of return falls below some threshold (Gallistel 1990); for instance, the running average rate of return of foraging in similar patches. To implement this, the mind of the organism contains counters, of sorts, that monitor benefits over time (Gallistel 1990).
For the present model, we propose that the allocation of mental processes to a task carries opportunity costs equal to the value of the next-best use of those mental processes. For example, the Stroop task engages the visual system and word recognition systems, among other mechanisms. It might not be possible to simultaneously perform other tasks that require one or more of the same systems. Similarly, working memory, we presume, cannot simultaneously be used for two different tasks that require it. Computations to prioritize its use must be made, and the analysis is identical to the analysis for behavioral options. Therefore, in the context of tasks such as the Stroop, the costs of performing the task X include the potential benefits of doing those other tasks (A, B, C, etc.) being precluded because the systems required for the task X cannot be used for alternatives A, B, or C. Performing any given task carries opportunity costs, and the size of these opportunity costs depends on the details of the systems recruited by the task. To the extent that a task recruits many systems, particularly those systems that are required for a large number of other tasks, it carries a large opportunity cost.
2.3.2. Phenomenology of perceived opportunity cost
Above we argued that phenomenology can be understood as the felt or experienced output of motivational systems, directing behavior toward net-positive fitness outcomes and away from net-negative fitness outcomes. We argued that felt sensations are the outputs of mechanisms designed to produce inputs to decision-making systems. This view resonates with other approaches to phenomenology (Bloom 2010; Thornhill 1998). Positive experiences in the domains of food (Rozin & Vollmecke 1986); environments (Orians & Heerwagen 1992); bodies (Buss 1989; Singh 1993); the arts (Kurzban 2007; 2012); and, of course, emotions (Tooby et al. 2008) can all be neatly explained in this way.
Using the same logic, and similar to recent proposals (Boksem et al. 2005; Boksem & Tops 2008; Botvinick 2007; Hockey 2011; Kool et al. 2010; Lorist et al. 2005), the crux of our argument is that the sensation of “mental effort” is the output of mechanisms designed to measure the opportunity costs of engaging in the current mental task (Kurzban 2010b; see Fig. 1). The function of these cost representations is to direct the reallocation of particular computational mechanisms away from the present task and toward the task that yields greater benefits.
Figure 1.
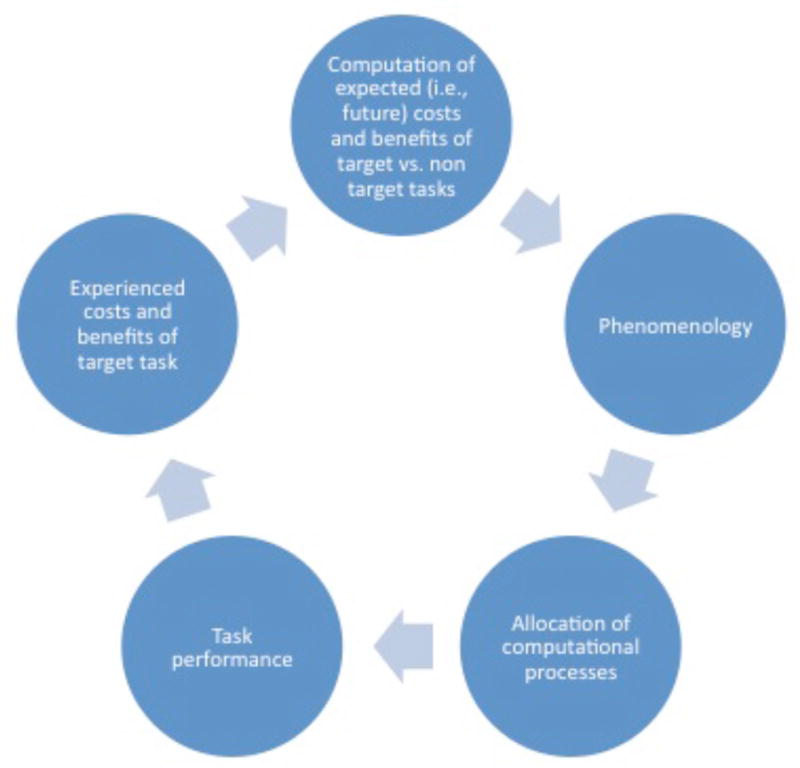
A schematic diagram of the proposed opportunity cost model. The expected costs and benefits of target and nontarget tasks are estimated (top). These computations give rise to phenomenology (e.g., qualia such as frustration, boredom, flow), which, in turn, motivates the allocation of computational processes to tasks that are expected to optimize costs and benefits. This allocation determines performance, on both the target and nontarget tasks. The experienced costs and benefits then recursively feed into another iteration of the same sequence, with continued adjustment of allocation decisions, but without depletion of any physical resource.
Our view resembles that of Kool et al. (2010), who proposed “that cognitive demand weighs as a cost in the cost/benefit analyses underlying decision making (p. 677).” Similarly, Hockey (2011) suggested that fatigue is “an adaptive state that signals a growing conflict in control activity between what is being done and what else might be done” (p. 168). Hockey’s (2011) model similarly posits an “effort monitor,” which functions to evaluate the value of pursuing the current goal relative to alternate goals: “Maintaining a specific cognitive goal means suppressing all others (investigating novel environmental events, attending to emerging thoughts, making a phone call, replying to an email). It is argued that the fatigue state has a metacognitive function, interrupting the currently active goal and allowing others into contention” (p. 173). In the same vein, van der Linden (2011) has suggested that “fatigue might be considered as a stop emotion” (p. 153, italics original), an idea proposed more than a century earlier by Thorndike: “Feelings of fatigue … serve as a sign to us to stop working long before our actual ability to work has suffered any important decrease” (quoted in Arai 1912, pp. 72–73).
Our model explains the well-documented experiences of boredom and mental effort associated with vigilance tasks. Performing such tasks requires deploying attention to the stimulus object. Monitoring the Mackworth Clock, for example, requires computations to determine whether the movement of the clock corresponds to the motion specified by task instructions, which presumably recruits working memory and other systems, which therefore cannot otherwise be engaged. To the extent that there are no offsetting benefits – other than, for example, compliance with experimenter requests to persist – the relationship between perceived costs and benefits can become less favorable over time, just as in the foraging example offered above.
We can also apply this idea to the experimental psychology literature on “self-control” (Baumeister et al. 2007). The tasks used in this literature – making complex choices (as opposed to simply remembering), keeping an instruction in working memory (e.g., “Do not think of a white bear”), inhibiting prepotent responses, math problems, and so on – all require systems that have many possible uses (Miller & Cohen 2001; Miyake et al. 2000; Stuss & Alexander 2000). As in the case of the vigilance tasks, we believe that it is useful to conceptualize executing self-control tasks as carrying the opportunity costs associated with these systems, and the phenomenon of effort to be the felt output of a motivational system designed to optimize the deployment of computations that cannot be used simultaneously, especially those associated with executive function.
In sum, many experiences, particularly the more or less unpleasant sensations discussed here (i.e., effort, boredom, fatigue), can be profitably thought of as resulting from (1) monitoring mechanisms that tally opportunity costs, which (2) cause an aversive state that corresponds in magnitude to the cost computed, which (3) enters into decision making, acting as a kind of a “vote,” influencing the decision ultimately taken.
2.4. Simple formal model
Here we sketch a formal model of our proposal to explain how our theory can account for the perception of effort, corresponding performance decrements, and the dynamics of both of these. Developing more detailed computational models that make quantitative predictions in specific tasks should be a critical aim of future research, but it goes beyond our current goal.
We start with the assumption that organisms solve the prioritization problem by estimating the utilities of different possible actions, and then selecting the action that has maximal expected utility. (See sect. 4.2 for neural evidence supporting this assumption.) We therefore start with the standard assumptions of rational choice, applying this logic to prioritizing mental actions. These assumptions are analogous to the approach in psychophysics, in which value maximization is (likely) the “ideal observer” solution for trade-off and prioritization problems. This makes it a natural starting point for thinking about the computations involved in solving trade-off and prioritization problems from a functional point of view. Of course, as is often the case in psychophysics (and elsewhere), cognitive mechanisms might only approximate the ideal observer solution.
2.4.1. An illustrative example
Consider, as an illustrative example, a research participant asked to perform a set of simple math calculations of the sort Arai (1912) and Huxtable et al. (1946) investigated. We can think of this participant as having a choice between performing those calculations or, alternately, daydreaming (and therefore not performing the problems). Performing the math calculations leads to various benefits in different currencies (e.g., monetary, class credit, social approval). Daydreaming’s benefits are more difficult to identify but may include reflection upon past experience and scenario planning for the future (Gilbert & Wilson 2007). The costs of these mental activities are simply their opportunity costs. In situations such as these, the opportunity cost of a chosen action is the value of the next-best possible action. Therefore, the opportunity costs of doing the math calculations are the forgone benefits of daydreaming.
(Note that we offer daydreaming as only one example of the kinds of “background” processes that one’s brain could engage in. Others might be planning future activities, reevaluating past actions, scanning the environment, etc.)
Suppose that we add a third possible action for our research participant. Sitting next to him (or her) is his smartphone, which he could use to check his email, log into Facebook, check sports scores, and so on. We assume that people are motivated to do these activities because they derive from them lots of valuable social information (e.g., who is trying to get in touch with them, who likes their latest status update, whether their team is winning the soccer match, etc.); but in this context, smartphone activities carry the potential cost of social disapproval from the experimenter. So let us assume that in the experimental context, playing with the smartphone is more valuable than daydreaming but less valuable than doing the experiment. Further, we will assume that we can attach a single number to each activity that is proportional to its expected utility (U). (See Fig. 2.)
Figure 2.
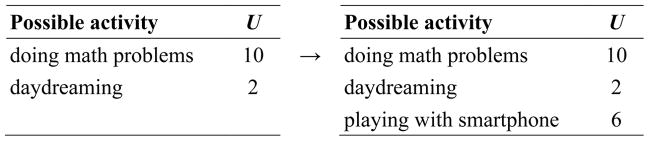
Hypothetical utilities of different actions a research participant might engage in, illustrating how opportunity costs depend on the set of actions available.
With the smartphone available, the opportunity costs of doing the math problems are now greater, since the forgone benefits of using the smartphone are greater than those of daydreaming. Our model predicts that doing the math problems in the presence of the smartphone will be perceived as more effortful than when the smartphone was absent, because the opportunity cost now is higher.
Perception of mental effort might correspond to different specific computational parameters, including (in the smartphone example) the opportunity cost of the current action (6), the ratio of that opportunity cost to the utility of the current action (6/10=0.6), or the difference between the two actions (10–6=4). We do not take a position here on exactly which of these most closely corresponds to the perception of mental effort, but we believe that this question could be answered empirically.
Experiencing mental effort does not always result in ceasing the current activity, and in the above example the participant should continue to do the (now more effortful) math problems. In some cases, though, the experience of mental effort precedes abandoning a task altogether. Returning to our example, imagine the experimenter leaves the room, changing the calculus of benefits for doing the experimental task (social disapproval for shirking is now less of an issue), as shown in Figure 3:
Figure 3.

How hypothetical utilities of different actions might change for a research participant with the experimenter present/absent, illustrating opportunity costs and the optimal action changing in different contexts.
The person doing math problems in this example should then cease this activity and shift to playing with their smartphone.
2.4.2. Allocating computational processes
The foregoing assumes that only one task at a time can be executed. In this section, we assume that the critical computational processes necessary for task performance can be divided among multiple tasks, that these processes can be allocated in different portions to different tasks, and that task performance varies with the degree to which computational processes are allocated to the task. We stress that in this view, mental “resources” are finite, dynamic, and divisible at any given point in time, rather than finite and depletable over time. A good analogy would be a computer with multiple processors that are dynamically allocated to computational tasks; the brain similarly has a finite number of mental “processors” that can be allocated to different tasks.
To see how these additional assumptions can explain decrements in task performance, consider again the math problems. Take the simplest possible case, in which there are just two mental processors and two possible activities (task 1, task 2). As shown in Figure 4, the value or utility (U) of allocating the processors to the tasks depends on how many processors are allocated to each task:
Figure 4.

Hypothetical utilities of dedicating computational processes to one task or dividing them between two tasks, illustrating how opportunity costs apply not just to the selection of tasks but also the allocation of processes among tasks.
Under the conditions shown in Figure 4, the participant with both mental processors allocated to the math problems (U=10) should shift to having the processors divided between doing math problems and daydreaming (U=11). If performance on math problems varies monotonically with the number of mental processors dedicated to a task, which is a likely assumption, then such a shift would result in decreased performance.
In this simplified case, dividing processors between two mental tasks should occur only if the marginal utility gained by reallocating one processor’s capacity to the next-best task is greater than the marginal utility lost by reallocating one processor from the best task to the next-best one.
To illustrate this with a simple mathematical example, consider the case where a person can focus on only one task or perform two tasks at once, when doing two tasks simultaneously achieves a fixed fraction (β) of the utility of doing each task in isolation:
where a1 and a2 are two tasks; U(a1,a1) is the value gained from doing only task a1; U(a2,a2) is the value gained from doing only task a2; and U(a1,a2) is that value gained from doing tasks a1 and a2 at the same time. β is an index of diminishing marginal utility, where 1 ≥ β ≥ 0. When β is high (near 1), the person already receives most of the possible value from a task under conditions where processing capacity is simultaneously divided between two tasks.
We can define the relative utility (RU) of the next-best action (a2) as the fraction of its utility relative to the utility of the best action (a1),
The conditions under which a person should do both tasks simultaneously is expressed thus:
Figure 5 shows the parameters under which the person should divide processing capacity between two actions rather than devote processing capacity exclusively to the highest-valued action. This occurs when the relative utility of the next-best action (RU(a2)) is high, and when there is diminishing marginal utility to devoting processing capacity entirely to one task relative to dividing it between two tasks (β is high). These two parameters control the opportunity cost of devoting processing capacity exclusively to the most valuable task. When the marginal value gained from the best task by dedicating processing capacity entirely to it is less than the marginal value gained from the next-best task by dividing processing capacity, processing capacity should be divided between the two tasks.
Figure 5.

For the simple model outlined in the text, whether one should focus attention on only the highest valued action or divide attention between the two best actions, as a function of the relative utility (RU) of the next-best action and the fraction of the value (β) one gains from a task when dividing processing capacity. These two factors determine the opportunity cost, and it is better to divide attention when the opportunity cost is high. The locations x and y in (B) provide an example of how to think about the dynamics of effort and performance. A person will feel an increased sense of effort, and be motivated to reallocate attention/mental processes in a way that reduces performance on a task, when the perceived costs and benefits of the task move from position x to position y.
Our examples above are clearly simplifications, but these examples have been intended to be illustrative only. Microeconomic models could provide a much richer framework to model these kinds of effects, one that does not depend on restrictive assumptions such as the utility from a task being directly proportional to performance, or the utility functions of the two tasks being similar in form. This richer framework would involve “production functions” that describe performance on multiple tasks as a function of the number of processors allocated to them, and “utility functions” that describe one’s preferences over performance levels on the different tasks. Such a framework was already offered some time ago, as an alternative to resource theories of attention (Navon 1984). Our hope here is that such a framework will gain greater traction in the field if reintroduced.
2.4.3. Dynamics of effort and performance
Empirically, cumulative time on task has been found to be the best predictor of sensations of fatigue (Kanfer 2011; see also Boksem et al. 2006). Why are some tasks perceived as progressively more and more effortful over time? Related, why does performance on vigilance tasks decline over time? And, why would performance on a second task decline after having done a first task?
Our view is that a person’s experience with a task over time provides information that updates estimates of expected utility. Figure 5 illustrates the optimal allocation between two tasks. Dynamics arise in how one reaches that optimal allocation. For example, imagine someone is currently devoting her (or his) entire processing capacity to one task, but would, because she is “at” point y, in the shaded portion of Figure 5, be better off dividing her processing capacity between the two tasks. In this case, we would expect the person to experience a sense of effort that would cause her to shift allocation and divide processing capacity between the two tasks.
A situation where processing allocations are suboptimal can arise for at least two reasons. First, the relative utility of the next-best action might be stable but the person does not know this value with any certainty, and so he or she has to learn it over time. This situation likely obtains anytime someone begins performing a novel task for the first time. Second, the relative utility of the next-best action might be changing over time, such that a previously optimal allocation is no longer optimal (as illustrated in Fig. 5 with a change from point x in the white portion of the figure to point y in the shaded portion). Thus, our theory explains dynamics of effort and performance as a result of learning the utilities and opportunity costs over time, as opposed to dynamic changes in the level of a resource.
Finally, we note that a framework explaining changes in mental effort and task performance as the result of dynamic learning processes can easily be expanded to incorporate trade-offs between exploration and exploitation. Even when the perceived utilities of the two best tasks are stable, it could be adaptive for there to be a small bias away from continuing to allocate processing capacity to the same task over time, which would also contribute to decrements in performance over time. As discussed extensively in the literature on reward learning (Cohen et al. 2007), such an exploration bonus would trade off exploitation of knowledge about the current task for gaining new and potentially valuable knowledge about different tasks.
3. Comparing our model with previous models
Broadly, two types of explanations have previously been proposed for reductions in performance in tasks that require vigilance or effort over time. One view is that information-processing resources or capacities are dynamically allocated in response to task demands. These resources/capacities have been conceptualized as unitary and domain-general (Kahneman 1973; Moray 1967) or multiple and domain-specific (e.g., Gopher et al. 1982; Navon & Gopher 1979; Wickens 2002). Some accounts have hypothesized that mental effort and task performance decrements are caused by the literal depletion of a resource (Gailliot & Baumeister 2007; Gailliot et al. 2007). Other accounts have located their explanation in the notion of motivation (Boksem et al. 2006; Boksem & Tops 2008; Hockey 2011; Nix et al. 1999; Robinson et al. 2010), positing that the repetitive, tedious nature of the task leads observers to withdraw effort over time and instead divert attention to other tasks. Some accounts combine these two approaches; still others draw on other computational frameworks (Gonzalez et al. 2011; Gunzelmann et al. 2009). Although it is beyond the scope of this article to address all alternate conceptualizations, this section describes how our model explains existing data, and distinguishes our model from some of these previous accounts.
Accounts of mental effort and task performance that rely on some notion of “resources” or “capacities” use these concepts with varying degrees of specificity, falling into two broad categories. Some accounts use the idea of resources loosely and analogically; researchers infer from task performance outcomes that the underlying cognitive system of interest behaves “as if” it were constrained by a limited resource, or that it has a “limited capacity” of some sort. Less common but recently rising in prominence are limited resource accounts in which the resource is specified. These two categories of resource/capacity accounts are briefly described below.
The most prominent account of mental effort as a limited capacity is probably Kahneman’s (1973) capacity model of attention. Kahneman’s account does not seek to explain the phenomenology of effort; rather, in his model, effort (which he refers to interchangeably as “attention” or “capacity”) is an assumed constraint for certain kinds of tasks with particular characteristics and therefore a constraint on task performance. The total amount of effort that can be used at any one time is limited and is used according to an allocation policy that changes over time based on task demands. Effort is thought to increase in response to demands such as the relative task “difficulty,” time pressure, and especially when two tasks are being done at the same time. In this model, effort is not literally a resource; instead, it is dynamic (allocated in response to changing task demands) but is not depletable. In this sense it is similar to models of attention that preceded it, most notably Moray’s (1967) model of attention, and also to later models of working memory (e.g., Baddeley & Hitch 1974; Posner & Snyder 1975; Posner et al. 1980).
Whereas Kahneman’s (1973) model of effort relied on a unitary and limited capacity (see also Moray 1967; Rolfe 1971), other models posit multiple capacities or resources. For example, Navon and Gopher (1979) proposed a model of multiple capacity usage analogous to the production of a firm, whereby performance on two simultaneous tasks depends on trade-offs resulting from shared inputs, the degree of demands on those inputs, and the chosen allocation policy (see also Gopher et al. 1982; Gopher & Navon 1980). In their model and other multiple capacity models (e.g., Wickens 2002), the putative resources are dynamic but, as in Kahneman’s (1973) model of effort, not depletable.
Other accounts that attempt to explain diminished task performance (and secondarily mental effort) use the idea of resources literally. Perhaps the most prominent nonmotivational account for explaining the sorts of effects we are interested here is the “ego depletion” model, found in the psychology literature on self-control. Tasks in this literature are similar to vigilance tasks (e.g., Davies & Parasuraman 1982; Head 1923; Mackworth 1948; Warm 1984; Warm et al. 2008), showing reductions in performance over time and giving rise to the phenomenology of effort. The principal focus is on performance reductions; measurement of subjective effort is typically used as a manipulation check (e.g., Muraven et al. 1998). This account suggests that performance on these tasks relies on a resource that can be depleted and has spawned a tremendous amount of research (recently reviewed by Hagger et al. [2010b]), and arguably represents the most influential model of diminished task performance after a putatively “difficult” task in the psychological literature. More recently, researchers in this tradition have attempted to specify the resource that is depleted and that leads to subsequent performance decrements (Gailliot & Baumeister 2007; Gailliot et al. 2007).
Numerous other accounts of mental effort and task performance rely on some notion of “motivation.” Although the term can be vague (see Niv et al. [2006] for a useful discussion), we believe that motivation has a role to play in explaining mental effort. (See especially Berridge [2004] for a thorough and useful discussion of motivation.) Indeed, previous models have linked costs and benefits with the notion of motivation. Among these models, the view that most closely resembles our own is Hockey’s (2011) “motivational control theory of mental fatigue.” Hockey suggests that the feeling of mental effort is a signal that functions to cause goal switching in humans. A rapidly growing literature echoes this focus on the adaptive nature of mental effort, whereby the expected costs and benefits motivate behavior toward more rewarding activities and away from less rewarding ones (e.g., Boksem et al. 2005; 2006; Boksem & Tops 2008; Kool et al. 2010; Kurniawan et al. 2011).
3.1. How the opportunity cost account explains existing data
As discussed above, our view bears a resemblance to proposals that explain reductions in performance as due to motivation. However, our view of motivation is a particular one and committed to the idea that the “motivation” to devote computational processes or attention to a task depends on the history of costs and benefits of executing the task. Our proposal also goes beyond previous motivational theories in not just specifying that mental activity is costly, but also identifying the source of the cost – namely, that engaging computational processes or attention on a task entails opportunity costs. Because our proposal relies on the computation of the relative costs and benefits of persisting on a given task, and so commits to a representation of value, we refer to our account as an opportunity cost model. Our view resonates with models such as the “sociometer” model of self-esteem, which suggests that self-esteem can be thought of as a measure of one’s value to others (Kirkpatrick & Ellis 2001; Kirkpatrick et al. 2002; Leary & Baumeister 2000; Leary et al. 1995).
Specifically, in the context of the Mackworth Clock task, our view is that when subjects comply with experimenter requests to attend to the task, the costs of doing so are represented – specifically the opportunity costs of the computational systems required for the task. In vigilance tasks, targets are rare. As a person gains more and more experience with the task, their estimate of the probability of a target, and therefore the expected benefit of fully attending to the task, declines. (Note that this explanation predicts that vigilance should increase right after a target occurs; other reinforcing stimuli should have similar effects.) With learning, the representations of costs grow with time on task and, absent offsetting benefits, are experienced as the sensation of fatigue, boredom, and/or stress – aversive subjective states, which in turn encourage disengagement with the task, and ultimately, performance reductions. In short, we would explain vigilance decrements with reference to subjects’ learning (implicitly or explicitly) about the value of devoting attention to the vigilance task versus dividing attention between the task and mind wandering (Gilbert & Wilson 2007).
Similarly, our account suggests that the difference between the consistent Stroop and the inconsistent Stroop is that the inconsistent Stroop requires systems that inhibit prepotent responses that are themselves useful for a number of other computations. The recruitment of these (executive) systems carries opportunity costs, which in turn are experienced as effort, eventually reducing performance.
What about performance effects in sequential paradigms, such as when one’s persistence on unsolvable anagrams is lower after having previously completed a Stroop task? Sequential effects can be explained by our account if there is some link between the expected utility of the second task and the costs and benefits of having performed the first task – perhaps because the two tasks are similar in some way, or maybe just by the virtue of both tasks being part of the same social interaction.
Feelings of mental effort are limited when extrinsic incentives are sufficiently high (Boksem et al. 2006; Lorist et al. 2005; Tops et al. 2004). Similarly, when a second self-control task is perceived as sufficiently important (e.g., it leads to money, it may help others or oneself), prior engagement with a first, “depleting” task has no effect on performance or perseverance (e.g., Muraven & Slessareva 2003). Because it is unclear what sort of a “resource” might be restored when the subject is paid or otherwise incentivized (see below), these effects point to a motivational account for explaining the results of studies in the self-control literature.
We propose, in short, that the phenomenology of effort is attenuated if one experiences reward of various forms. This is necessarily the other half of the cost/benefit equation. Activities will seem less aversive, and therefore allow persistence, to the extent that benefits of various forms are received. These predictions already have some support (see sect. 3.3.2).
In this framework, beliefs and perceptions can lead to increases in task performance, again through learning. For example, this is how our theory would explain increased effort at resisting smoking at time 2 after having successfully resisted smoking at time 1 (O’Connell et al. 2008); the first successful effort likely increases one’s belief that subsequent efforts will also be successful.
Costs, of course, also matter. Consider that when subjects participate in laboratory experiments, they are doing so, generally, because they are receiving compensation in the form of either partial course credit or monetary payment. Hence, experimental sessions are explicitly exchanges in which the subjects give their time (and “effort”) in exchange for credit or cash. This explicit exchange – along with implicit norms that govern the relationship between subjects and experimenters in such contexts (Orne 1962) – explains why subjects comply with experimenter instructions and requests.
There are, of course, limits to what subjects will do. Subjects’ decisions to comply can be affected by the amount of effort that is appropriate given the compensation they expect to receive (Akerlof & Yellen 1990; Fehr et al. 2009). That is, people expend effort as a function of what they construe as “just” or “fair” given the exchange relationship (Fehr et al. 2009). Studies have shown, for instance, that subjects are more likely to do favors having previously received an unsolicited gift (Regan 1971); that surveys are more likely to be completed and returned when accompanied by an up-front small payment than by the offer of a large payment upon completion (James & Bolstein 1992); and that tips are more likely when food servers offer customers a candy with their check (Lynn & McCall 2000).
So, to the extent experimental participants in a self-control treatment perceive themselves as having discharged more of this obligation than those in a control treatment, participants might be expected to expend less effort on the subsequent task. Given that “self-control” tasks usually evoke a sense of effort, the perception of having discharged an obligation might explain why subjects in self-control conditions exert less effort. This locates the similarity of results across self-control tasks not in a resource but in the felt sense of effort these tasks evoke, in concert with the construal of the experimental context as an exchange.
One challenge to this argument comes from data showing that the size of depletion effects is not reduced by changing experimenters between the initial depleting task and the later task, nor by presenting the two tasks as a single experiment (Hagger et al. 2010a). However, if subjects find the self-control treatment aversive (Hagger et al. 2010a), and therefore understand their obligation to give a certain amount of effort in exchange for the credit that they are receiving, then they may understand their obligation to be reduced after the expenditure of effort even if a new experimenter is encountered in a second part of an experimental session. Indeed, consistent with this type of interpretation, DeWall et al. (2007), for example, reported that participants behaved more aggressively after performing a self-control task (see also Stucke & Baumeister 2006). In short, devoting attention to the task might be represented as a cost paid to offset the benefit (e.g., course credit) they are to receive. As they discharge more of the benefit over time, the residual they “owe” for the hour of credit diminishes. This might help to explain task carryover effects – over time, subjects owe less attention, and the endurance of the sensation of effort, in return for credit.
3.2. Comparison with resource accounts
The accounts that are perhaps most different from ours are resource models, in which performance depends on a depletable resource. A version of the resource model proposed by Baumeister and colleagues is also the most prominent explanation for performance decrements in the self-control literature in psychology. It is therefore instructive to explicitly consider their model and similar resource accounts in some detail.
Muraven and Baumeister (2000) presented five assumptions of this model: (1) Self-control “strength” is necessary for self-control. (2) Self-control strength is limited. (3) The resource on which this strength is based is used across self-control operations. (4) Task performance depends on one’s self-control strength (although “impulse strength,” among other factors, might also influence performance). (5) Exerting control exhausts self-control strength. These assumptions give rise to a family of models, depending on how performance “depends on” the level of the resource (assumption 4), as we review below.
Note that in this literature, researchers tend to use the experimental structure described above, in which a subject does one task that putatively requires the self-control resource, ranging from not eating tempting brownies, to doing an inconsistent Stroop task, to showing no emotion while watching a funny video, and so on. (Subjects who have completed such a task are referred to as “depleted.”) Subsequently, subjects do a second task that also putatively requires the self-control resource.
3.2.1. Theoretical assumptions of resource models
Distinguishing our model from resource models is challenging because resource models have multiple interpretations. On one interpretation, performance could depend on the level of a resource in a very strict way, with the level of the resource putting an absolute upper limit on performance. As an analogy, consider an electric pepper grinder; as the batteries get close to being drained, operation is limited by the remaining charge. According to this model, for any given amount of resource, there is a fixed maximum level of performance. We will refer to this as the strict capacity model, because it holds that the causal locus of observed performance reductions is the capacity for performance. This model carries the very strong entailment that, as a literal and physical matter, nothing could improve the performance of depleted subjects, as in the case of a pepper grinder with a nearly depleted battery. As Baumeister and Vohs (2007) put it, using a reservoir analogy: “If the tank were truly and thoroughly empty, it is unlikely that increasing incentives would counteract depletion” (p. 125). The large amount of data showing that incentives do counteract “depletion” is strong evidence that the strict capacity model is false (Baumeister & Vohs 2007; Baumeister et al. 2007; Muraven & Slessareva 2003).
Indeed, as Baumeister and Vohs (2007) put it: “Ego depletion effects thus indicate conservation of a partly depleted resource, rather than full incapacity because the resource is completely gone.” This suggests a second type of model – that the amount of the putative resource puts, in principle, an upper (capacity) limit on self-control performance, but that performance reductions are not a strict necessity (Muraven et al. 2006). This view suggests that “depleted” subjects could – perhaps by virtue of changed incentives – perform without any decrement or perform worse than controls. As an analogy, consider a soldier taking fewer shots because she is running low on ammunition but not yet out.
The second model, then, is that “depleted” and “nondepleted” subjects are capable of equal performance, but “depleted” subjects do not deploy self-control resources. This carries the implication that all of the effects in this literature are due to a decision by the subjects not to use self-control resources, rather than a limit on their capacity for self-control per se. In other words, this model holds that the reduction in the resource is not the immediately proximate causal variable, but only indirectly related. As Muraven et al. (2006) write, “The moderation of depletion by motivation suggests that self-control suffers in many situations because individuals are not unable but instead are not willing to exert sufficient self-control to overcome the impulse” (p. 525).
This model implies that no data can be directly explained by the capacity restriction. Instead, all the data are explained by a reduced capacity that caused a change in motivation to persist, and that this reduction in motivation directly caused performance reductions. A related view is that the amount of the putative resource matters, but so too does motivation, such that the level of the resource and motivation jointly determine self-control performance.
Muraven and Slessareva (2003), for instance, argued that their data support the view that “depletion of self-control strength does not prevent the subsequent exertion of self-control” (p. 897). This implies that the putative resource is not necessary for self-control, or minimally that self-control can be exerted in the absence of some quantity of the putative resource. The problem with such a view is that any observations of performance reduction can be accommodated by the claim that something was depleted and resources husbanded. Observation of continued performance can be accommodated by the view that something was depleted but no husbanding took place. Without independent means of measuring the resource and motivation, no data can falsify the model. This model runs into the problem faced by resource accounts in general, as pointed out by Navon (1984), who observed that the
frequent cases in which the predictions do not bear out are dismissed by resorting to built-in escapes in the theory, such as, data limits, operation below full capacity, disparate resource composition, and so forth. This is probably the source of the self-reinforcing nature of the concept and the unfalsifiable status of the theory.” (p. 231, emphasis added)
It could be that one route to evaluating this model would be studies in which performance was compared between “depleted” and “nondepleted” subjects, with motivation held constant. However, because “depleting” tasks, we would argue, can affect motivation, this design represents a methodological challenge in the absence of good tools to measure motivation and the putative resource accurately.
Finally, a third model is that the amount of the resource that is available directly limits performance, but that motivation can (in some way) causally influence the amount of the resource. On this model, motivation is an antecedent variable that influences self-control performance indirectly – that is, the order of the two causal variables is reversed as compared with the strict husbanding model. For instance, Tice et al. (2007) showed that when subjects performed an initial self-control task, there were no adverse effects on a subsequent self-control task when they experienced positive affect in the intervening time period – either from watching a funny video or receiving an unexpected gift. Tyler and Burns (2008) found similar effects with relaxation interventions, and Schmeichel and Vohs (2009) found similar effects with self-affirmation interventions. Tice et al. (2007) argued that positive affect might be able to “effectively replenish the depleted resource” (p. 380). We are uncertain what sort of mechanism might literally have this effect. We also note that this view is inconsistent with the view that the resource is something physical (e.g., glucose; see below).
3.2.2. Empirics of resource models
In addition to the concerns in the previous section, there are empirical results that seem hard for resource views to accommodate. Martijn et al. (2002) had subjects watch a brief video and had some subjects suppress their emotional expression, a task previously shown to yield performance reductions (Muraven et al. 1998). Martijn et al. then manipulated beliefs about self-control, telling some subjects that the intuitive theory that self-control is a limited resource is incorrect. The dependent measure was the difference in performance on a handgrip task before and after watching the video. The resource model predicts no effect of such beliefs. However, people who were given the emotion suppression manipulation but also told that the intuitive resource model of self-control was false showed an increase in performance on the handgrip task. Along similar lines, Job et al. (2010) recently showed that “depletion” effects depend on people’ beliefs. Those who did not indicate agreement with the idea that energy is depleted by a taxing mental task did not show the reduction in performance frequently observed in a two-task design.
In addition, Converse and DeShon (2009), drawing on research on “learned industriousness” (Eisenberger 1992), had subjects complete a perceptual task – finding differences between two images – then a math task (for which subjects were financially incentivized to answer correctly), and then an anagram task. One group of subjects were given perceptual and math tasks that were more taxing than the other group’s. The more taxing tasks should lead to performance reductions in these subjects; however, the reverse occurred: Subjects in the more difficult condition persisted longer on the anagram task than the other group did. This effect was replicated when different “depleting” tasks were used (and incentives in the second task omitted).
Similarly, Dewitte et al. (2009) had subjects perform a “response reversal” task – performing one action when they saw particular stimuli, but reversing the action when shown those same stimuli under particular conditions. Subjects who suppressed thoughts of a white bear subsequently performed worse on this task compared with controls, as predicted by resource models. However, consistent with their predictions derived from “control theory” (Miller & Cohen 2001), Dewitte et al. found that subjects who did one response reversal task subsequently performed better than both controls and those who had engaged in thought suppression did. Similarly, subjects who did task reversal twice performed better the second time than the first time. (For similar results, see Eisenberger & Masterson 1983; Hickman et al. 1998.) Such improvements are difficult for a resource model to explain, although they could perhaps be accommodated to the extent that these results could be attributed to practice effects.
Likewise, framing a laboratory task such as squeezing a handgrip as long as possible as a test of a subject’s “willpower” improves performance compared with a neutral framing (Laran & Janiszewski 2011; Magen & Gross 2007). Finally, Ackerman et al. (2009) found that participants asked to mentally simulate the perspective of another person exerting self-control subsequently showed less self-control themselves.
In short, the theoretical and empirical difficulties for resource accounts suggest that alternatives, such as our proposal here, might be of value in accounting for the array of effects in this literature.
3.3. Comparison of models and predictions
Above we discussed evidence from prior empirical studies that in our view supports an opportunity cost model of mental effort. Here we summarize how our model’s predictions diverge from alternate accounts of mental effort, some of which are supported by prior studies, but most of which have yet to be directly tested.
First and foremost, whereas both our model and the resource account posit limits to mental activity, the nature of the limitations is different. In the resource account, mental resources are depletable: finite and destroyed with use. In our proposal, computational processes are dynamic: finite but not destroyed with use. The resource view holds that performance reductions result because some physical substrate in the brain (e.g., glucose) is literally depleted during self-control tasks. In contrast, our model suggests that performance reductions reflect the operation of a system designed to motivate disengagement with the present task when the opportunity costs are sufficiently high. Because computational processes are dynamically allocated rather than irreversibly (over short time spans) depleted, our model predicts that performance in self-control tasks might under specific circumstances improve over time, even in the absence of practice effects.
A second distinction concerns phenomenology. We suggest that the estimation of opportunity costs gives rise to the phenomenology of mental effort. These feelings (e.g., fatigue, boredom) in turn motivate the reallocation of computational processes away from a task to alternate, higher utility activities. The phenomenology of mental effort in our view is generally adaptive, favoring changes in behavior that are, in most circumstances, beneficial to the individual. The resource account, in contrast, suggests that the (perception of the) literal depletion of some substance gives rise to the phenomenology of mental effort. Whereas subjective experience in the resource account is, hence, both veridical and epiphenomenal, our view holds that subjective experience of effort is a representation that is neither always veridical (insofar as estimates can be wrong) nor epiphenomenal (insofar as feelings motivate behavior).
Third, our model specifically locates the costs of mental effort in opportunity costs. Several prior models have suggested that the mental effort precipitates an aversive subjective experience, which people seek to avoid. However, our model is distinct insofar as we specify what, in particular, makes mental tasks feel effortful – namely, the expected value of the next-best alternate use of the same computational processes. Importantly, it is not only the costs and benefits of performing the task at hand that give rise to the phenomenology of mental effort, but also the costs and benefits of rival activities to which the same computational processes might otherwise be directed. Crucially, and in line with existing data, tasks that recruit mechanisms that can be flexibly deployed should feel more effortful and demonstrate the most precipitous declines in performance, whereas mechanisms that are singular in their function should not. Solving four-digit multiplication problems feels “hard” in this view, because the required computational processes could be deployed to an alternate, profitable use (including prospection, daydreaming, and other “off-task” varieties of mentation). Vision, which also entails substantially complex computational processing, does not feel like anything at all, because the required computational processes are specialized for a particular purpose and cannot be flexibly deployed to alternate tasks unrelated to vision.
We have suggested that within-individual changes in the performance of mental tasks depend on estimates of their expected utility. Accordingly, one class of experiments useful in distinguishing accounts might replicate the two-task experimental paradigm from the resource model literature with one important modification: parametric variation of the expected utility of the second task. Our model predicts either declines or improvements in performance on the second task depending on the experienced costs and benefits of the first task. In contrast, only declines in performance – not improvements – are predicted by the resource model. Already, several published studies have shown that input to a variety of reward systems (in the form of calories, positive feedback, a gift) directly following the first task indeed improves performance on the second task (e.g., Eisenberger 1992; Gailliot et al. 2007; Tice et al. 2007). Additional studies might test whether other forms of reward produce the same pattern of findings, whether associating rewards more explicitly with performance in the first task strengthens these effects, and whether parametrically varying rewards produces systematic dose-response improvements in performance.
A second class of predictions to which our view is committed is that alternate activities one might be able to do should influence performance. Parametrically varying the appeal of an alternative – a more- versus less-rewarding alternate activity to the one that is being performed – should lead to systematic differences in performance. In the limiting case, participants performing self-control tasks without any alternate activity are predicted to perform better than participants performing the same tasks with an appealing alternative (e.g., using their smartphone) available. Likewise, the well-documented decrement in performance in the single-task vigilance paradigm should be potentiated or attenuated using the same manipulations. Performance in the target task should also be influenced by the expected utility of less obvious alternatives, such as daydreaming. The expected utility of, say, prospection and scenario planning might be increased or decreased by manipulating people’s beliefs about these activities. Our model predicts that making the benefits of off-task mental activity salient should decrease performance on the target task, whereas making the costs of off-task mental activity salient should increase performance on the target task.
Our model makes similar commitments in terms of predictions regarding phenomenology, although research in this area has been limited. Indeed, in a recent review, Ackerman (2011) noted that whereas “(f)ew studies have involved explicit measurement of changes in subjective fatigue in the context of higher order cognitive task performance” (p. 25), we agree with his prediction that “it can be expected that most task situations that result in mean decrements in performance with additional time on task will also show a marked increase in subjective fatigue” (p. 27). (We, however, recognize that performance and phenomenology might be dissociated in rare pathological cases; see Naccache et al.[2005].) For instance, manipulations that change performance should also change the corresponding subjective experience of mental effort (e.g., feelings of boredom, stress, etc.). Because we claim phenomenology drives behavior, we also expect changing phenomenology to change performance. Positive mood inductions before the second task should improve performance (Tice et al. 2007); in contrast, inducing feelings of boredom (e.g., perhaps by having the participant do an easy but extremely repetitious task) before the second task should impair performance. Blunting the phenomenology itself, for instance, by suggesting to participants that their mood will be stabilized by a (placebo) pill (Cialdini et al. 1987), should improve performance on self-control tasks; suggesting to participants that they pay careful attention to their feelings might have the opposite effect. Manipulating attributions of boredom or effort should also have an effect. Indeed, framing a task as a test of willpower, as Magen and Gross (2007) did, might have improved performance because it changed attributions of mental effort.
We recognize that a serious challenge for our model is that many effects in the experimental literature are found in studies with two different tasks, both of which require self-control but are quite different from each other. The variety of effects from one task to another is a key feature of this literature, and it might seem at odds with a cost/benefit account. As indicated above, however, any use of the relevant systems might be represented as a cost. In such a case, carryover effects are possible, just as in the resource case, because related mechanisms are used across tasks. To the extent that the mechanism (or mechanisms) that computes costs takes as input only the fact that (some subset of) executive systems are being used, rather than which ones in particular are being deployed and/or what they are being used for, such carryover effects are possible. Further, as indicated above in section 3.1, persisting in tasks steadily reduces the debt owed for experimental credit, perhaps explaining reductions in effort.
Disentangling these accounts might be difficult. We predict that similarity across tasks – in the sense of which executive function systems are engaged – will lead to greater decrements in performance, but similar tasks also might show learning effects. The more similar the tasks, the lower the expected value of the second task given a poor experience (i.e., low perceived benefits) on the first task. Research on tasks in which subjects are at ceiling might be of use to limit learning effects while allowing the use of similar tasks at time 1 and time 2.
Our model also makes an important prediction regarding interventions aimed at increasing self-control. Specifically, we suggest that self-controlled behavior is reinforced over extended periods of time only when it is practiced and rewarded, whereas proponents of the resource account posit that repeated exertion of self-control followed by rest should improve performance regardless of whether behavior is rewarded. In other words, we believe that individuals will improve in self-control through a learning process, whereas a resource account suggests a mindless process akin to muscle building in which performance-contingent rewards are irrelevant.
Some data from the field are interesting in this respect. O’Connell et al. (2008) found in a prospective, longitudinal study of individuals who were trying to quit smoking that resisting urges to smoke predicted fewer – not more – subsequent lapses in the immediately ensuing 4-hour period. That is, exerting self-control increased, rather than reduced, subsequent self-control efforts, “providing a direct challenge to a resource depletion model of self-control” (p. 492). We suggest that smokers who are trying to quit might construe a period of sustained abstinence to be a victory – and, therefore, a reward that motivates further abstinence. More generally, we predict that interventions that provide performance-contingent feedback and/or external rewards should be more effective than interventions that do not.
Finally, our model entails certain requirements for its neural implementation that differ from those entailed by a resource account. A resource account predicts that there should be some physical resource that is depleted by mental tasks, and that there is a link between the level of this resource and task performance. In contrast, our model predicts that there should be neural systems that can be used flexibly for different tasks, thus creating a simultaneity problem; that tasks that feel effortful engage these neural systems; and that there are neural representations of costs and benefits appropriate for guiding decisions about continued task engagement. We now turn to the neuroscience evidence bearing on these issues.
4. The neuroscience of resources and motivation
A wealth of evidence from neuroscience is relevant to debates regarding subjective effort and task performance. This section considers resource accounts and the proposed opportunity cost account in this context.
4.1. The neuroscience of resources: The role of glucose in mental tasks
One proposal is that glucose is the putative resource depleted when effortful tasks are executed (Gailliot & Baumeister 2007; Gailliot et al. 2007; for the related view that the issue is the allocation of glucose, see Beedie & Lane 2012). There are, however, reasons to doubt this account. Indeed, Hockey (2011) recently suggested that the reason that fatigue has remained mysterious despite intense study is “the irresistible tendency to think of it in terms of a loss of energy resources,” that there is “no evidence” for the claim that “fatigue is the result of glucose depletion,” concluding that “there is little doubt that the energy-depletion perspective has been a source of distraction in the search for a theory of fatigue” (p. 167). However, because of the prominence of the idea, we address it very briefly here. (See also Kurzban 2010b.)
Although there is some evidence that cognitively taxing tasks reduce blood glucose levels (Fairclough & Houston 2004; Scholey et al. 2001), such results are inconsistent (Gibson & Green 2002) and leave open the possibility that reductions are due to activity in the peripheral systems, such as the heart, rather than in the brain. Recent reviews of the relevant empirical work in this area have generally converged on the view that any changes in blood glucose are unlikely to be due to increased uptake in the brain (Clarke & Sokoloff 1998; Gibson 2007; Messier 2004). Further, recent research using sensitive measuring devices has confirmed that blood glucose levels do not go down when participants perform a “self-control” task (Molden et al. 2012); and reanalysis of Gailliot et al.’s (2007) data has shown that their inferences were statistically unsound, rendering their conclusions “incredible” (Schimmack, in press).
This conclusion resonates with quantitative analyses of brain metabolism. Local changes in cerebral metabolism due to engaging in an experimental task are very small relative to the rate of metabolism at rest (Raichle & Gusnard 2002). The largest local changes in glucose consumption (~25%) are observed in visual cortex in response to opening one’s eyes (Newberg et al. 2005). So, if blood glucose were the resource, the visual system would be most sensitive to performance decrements; and if nutrient consumption caused sensations of effort, seeing would feel effortful. Further, under reasonable assumptions, the overall difference between self-control tasks and control tasks – the inconsistent Stroop versus the consistent Stroop, for instance – is miniscule in terms of calories consumed (Kurzban 2010a). In addition, exercise, which consumes orders of magnitude more glucose, improves – rather than impairs – subsequent performance on tasks such as the Stroop (Tomporowski 2003; see also Hillman et al. 2008; 2009).
The effects of glucose administration on task performance are often cited as support for blood glucose acting as a resource (see Gibson 2007 for a review). However, another possibility is that glucose is a signal rather than a resource. Consider that glucose is known to act on the brain’s reward circuitry, both through receptors on dopamine neurons (Hommel et al. 2006) and indirectly (i.e., with delivery of glucose into the mouth; McClure et al. 2003; O’Doherty et al. 2003). Further, glucose can have behavioral effects similar to that of drugs of abuse that target the same circuitry (Avena et al. 2008). Glucose can therefore invigorate subsequent behavior in the same manner as other rewards, and quite independent from the calories provided (Hagger et al. 2009). Consistent with this, in the context of physical performance, improvements can occur when glucose is only swished around the mouth, rather than digested (Chambers et al. 2009; Jeukendrup & Chambers 2010). Indeed, recent work shows that swishing without swallowing the glucose solution eliminates the “depletion” effect (Molden et al. 2012).
In sum, the empirical evidence weighs heavily against the claim that glucose is the resource on which performance on self-control tasks depends.
We know of no other explicit proposals identifying the putative resource, but acknowledge that there are many possibilities beyond glucose. Any such theory, however, will need to explain (1) what the resource is, (2) how that resource is depleted by effortful tasks, (3) how depletion of the resource is sensed and leads to subsequent decrements in task performance, and (4) why some kinds of mental/neural activity, but not others, lead to resource depletion. This fourth point could turn on differences in architecture across brain regions, but we know of no proposal that has identified the specific resource and the important architectural differences.
4.2. The neuroscience of costs and benefits
Abundant evidence exists for neural signals related to the costs and benefits of engaging in different tasks (Kable & Glimcher 2009; Lee et al. 2007; Rangel et al. 2008; Rangel & Hare 2010). Signals of exactly this type would be required by any computational mechanism that adjusts performance in accordance with cost/benefit trade-offs.
These signals are most prominent in an interconnected network that involves the prefrontal cortex and basal ganglia (Haber 2003; Haber & Knutson 2009). One part of this network involves the prefrontal cortex and a part of the basal ganglia called the striatum: The prefrontal cortex directly projects to the striatum, which sends indirect projections back through the globus pallidus (another part of the basal ganglia) and thalamus. Another part of this network involves dopaminergic neurons, which are located in other nuclei of the basal ganglia and send and receive prominent connections to both the prefrontal cortex and striatum. Further, these prefrontal-striatal-dopaminergic loops are partially segregated. Cost/benefit signals are most prominent in the orbital and medial sectors of the prefrontal cortex and the corresponding ventral sectors of the striatum (Kable & Glimcher 2009; Lee et al. 2007; Rangel et al. 2008; Rangel & Hare 2010). Lateral prefrontal cortex and associated striatal regions appear to have a different function, as discussed further below.
One prominent hypothesis is that the dopaminergic neurons encode a reward prediction error signal, equal to the difference between the reward expected and the reward obtained (Montague et al. 1996; Schultz et al. 1997). This kind of signal is used in computational algorithms for reinforcement learning. These algorithms learn from experience the overall values of states and actions, integrated over the various costs and benefits associated with those states and actions (Sutton & Barto 1998). Although the initial evidence for this hypothesis came from animal models (Schultz et al. 1997), evidence consistent with it has recently been obtained with pharmacological (Pessiglione et al. 2006; Rutledge et al. 2009), functional imaging (D’Ardenne et al. 2008), and neural recording (Zaghloul et al. 2009) techniques in humans.
An extension of this hypothesis is that the prefrontal and striatal neurons receiving dopaminergic input encode the overall integrated value of different states and actions (Kable & Glimcher 2009). In other words, they encode the quantities that can be learned from reward prediction errors. Evidence consistent with this hypothesis has been gleaned from single neuron recording (Lau & Glimcher 2008; Padoa-Schioppa & Assad 2006; 2008; Samejima et al. 2005), functional imaging (Kable & Glimcher 2007; Plassmann et al. 2007; Tom et al. 2007), and lesion studies (Camille et al. 2011; Fellows & Farah 2007; Rudebeck et al. 2008).
Though there are alternative views regarding the prefrontal-basal ganglia network (e.g., Berridge 2007), the debates concern the precise nature of the signals carried in different regions. All theories share the core notion that this network plays a critical role in motivation and reward.
Importantly, orbital/medial prefrontal and ventral striatal regions respond to multiple categories of rewards and integrate multiple factors to encode reward value. These properties, which allow for the incorporation of diverse kinds of benefits, from food to social approval, are required for computing the overall benefits of task performance. Increased activity in ventral striatum has been observed in response to primary rewards such as food (McClure et al. 2003; O’Doherty et al. 2003), secondary rewards such as money (Kuhnen & Knutson, 2005), and social rewards such as positive social comparison or one’s rivals experiencing pain (Fliessbach et al. 2007; Hein et al. 2010; Singer et al. 2006). During decision making, prefrontal and striatal activity reflects the perceived value of potential outcomes, integrating over diverse factors such as the taste and health value of foods (Hare et al. 2009); the magnitude, delay, and risk of monetary rewards (Kable & Glimcher 2007; Tom et al. 2007); or the benefit to others and costs to oneself of social exchange (Harbaugh et al. 2007; Hare et al. 2010).
4.2.1. Neural systems for effort trade-offs
Much evidence illustrates the importance of this prefrontal-basal ganglia network in regulating the performance of tasks that require physical effort (for review, see Kurniawan et al. 2011; we discuss mental effort below in sect. 4.4.2). For example, in one well-studied paradigm, animals choose between climbing a small barrier to obtain a less desirable food reward and climbing a large barrier to obtain a more desirable one. In this paradigm, depletion of dopamine in the ventral striatum shifts animals’ preferences away from the high effort–high reward option (Salamone et al. 2009).
A computational account of this result and others begins with the proposal that if dopamine neurons phasically respond to reward prediction errors, then the tonic baseline level of dopamine in the ventral striatum would be proportional to the average reward rate in a given environment (Niv et al. 2007). This quantity is important, because if animals are deciding how fast to work (one measure of effort), then the average reward rate is exactly the opportunity cost of working more slowly.
Other evidence suggests an important role for the anterior cingulate cortex (a specific region on the medial prefrontal surface) in making effort trade-offs. Lesions to the anterior cingulate also shift animals’ preferences away from high effort–high reward options (Rudebeck et al. 2006; Walton et al. 2003), and the costs of physical effort are robustly encoded in this region (Croxson et al. 2009; Kennerley et al. 2009; Kurniawan et al. 2010; Prevost et al. 2010). Further, the anterior cingulate is well positioned to compute the overall costs of task performance, because it responds to diverse kinds of costs, ranging from physical pain (Botvinick et al. 2005; Singer et al. 2004) to decrements in reward (Bush et al. 2002) to social disapproval (Klucharev et al. 2009). The anterior cingulate also responds to opportunity costs, such as what one would have received having chosen differently (Hayden et al. 2009).
4.3. The neuroscience of executive function
4.3.1. Effortful tasks engage a prefrontal executive network
The preceding section outlined evidence for a brain network that computes costs and benefits, and the involvement of this network in calibrating performance of tasks that require physical effort. Here we turn to what is known about the brain networks engaged by effortful mental tasks.
Put briefly, the effortful tasks that show decrements in performance all engage prefrontal regions associated with executive function. Different executive function tasks all reliably engage a network of brain regions that includes the lateral prefrontal cortex (inferior and middle frontal gyrus), dorsomedial prefrontal cortex (superior frontal gyrus and anterior cingulate), and posterior parietal cortex (typically intraparietal sulcus) (Buchsbaum et al. 2005; Derrfuss et al. 2005; Laird et al. 2005; Nee et al. 2007; Neumann et al. 2005; Wager & Smith 2003). Although different executive function tasks more strongly engage different parts of this network, the pattern of activation in executive function tasks as a class is distinguishable from patterns observed in perception, language, or semantic or episodic memory tasks (Cabeza & Nyberg 2000; Wager & Smith 2003).
Many of the tasks used to study mental effort or performance decrements are identical to those used in cognitive neuroscience to study executive function. This includes the sustained attention tasks used in vigilance experiments (Coull et al. 1998; Lim et al. 2010; Paus et al. 1997) and the Stroop and working memory tasks used in “depletion” experiments (Derrfuss et al. 2005; Laird et al. 2005; Neumann et al. 2005; Schmeichel 2007; Wager & Smith 2003; Wright et al. 2007). In other cases, the tasks used in the two literatures are not identical but are quite similar. For instance, two of the more widely used tasks to elicit decrements in performance – a crossing out letters task (“Cross out all e’s except for those adjacent to a vowel”) and a focus-of-attention task (“Attend to the person in the video and ignore the words”) – are similar to widely studied response inhibition and attentional control tasks such as the “go no-go” (Buchsbaum et al. 2005; Nee et al. 2007) and attention networks test (Fan et al. 2002).
Other tasks used to study mental effort or performance decrements also engage the prefrontal regions associated with executive functions. This includes tasks used to elicit subsequent decrements in performance, including regulating emotional responses (Ochsner & Gross 2005), suppressing a specific unwanted thought (Mitchell et al. 2007; Wyland et al. 2003), and turning down unhealthy but “tempting” foods (Hare et al. 2009). This also includes tasks used to measure decrements in performance, such as solving anagrams (Schneider et al. 1996), solving mathematical problems (Dehaene et al. 1999; Nieder & Dehaene 2009), or logical reasoning (Goel 2007).
4.3.2. Engaging the prefrontal executive network entails opportunity costs
The lateral prefrontal cortex regions engaged by effortful tasks play an important role in “controlled” aspects of cognition. The prefrontal cortex receives input from all modalities and, in addition to reciprocal connections to posterior regions, also sends output to the motor system. It is therefore anatomically well situated to influence how sensory and internal regulatory signals affect motor behavior. Miller and Cohen (2001) proposed that by actively maintaining information such as task goals and rules, the prefrontal cortex biases the flow of neural activity in other brain regions so that actions are affected by the behavioral context. This general idea, that the prefrontal cortex exerts a modulatory influence over information processing in other brain regions, forms the basis of more specific proposals regarding prefrontal function in attention (Desimone & Duncan 1995) and language (Thompson-Schill et al. 2005).
Consistent with this role in controlled cognition, the lateral prefrontal cortex is engaged by an array of different tasks, spanning different cognitive domains. This is apparent at the region level in the neuroimaging studies cited in the previous section. It is also apparent at the level of single neurons: The same lateral prefrontal neurons have been shown to respond to very different stimuli under different task conditions (Freedman et al. 2001; Rainer et al. 1998; Rao et al. 1997). Duncan (2001) argues that such “adaptive coding” in response to task demands is a special characteristic of the prefrontal cortex.
The prefrontal cortex is also subject to simultaneity constraints, in that there is a capacity limitation to the number of computational operations that the prefrontal cortex can engage in at any given time (Miller & Cohen 2001). Although the precise nature of the capacity limitation is unknown, our view echoes Miller and Cohen’s: “[N]o theory has provided an explanation of the capacity limitation itself. This could reflect an inherent physiological constraint, such as the energetic requirements of actively maintaining representations in the PFC. More likely, it reflects fundamental computational properties of the system” (2001, p. 192, italics added for emphasis).
These factors imply that there will be large opportunity costs to performing tasks that recruit the prefrontal cortex, given all of the tasks that cannot be performed simultaneously because they require the same prefrontal processes. To the extent that engaging these processes at all also requires disengaging others, such as the “default mode network” (Raichle et al. 2001), the functions achieved by these other processes would also contribute to the opportunity costs.
4.3.3. Other constraints: Specialization in the prefrontal cortex
A potentially important set of observations that any theory of effort must account for is that there is anatomical specialization within lateral prefrontal regions. Although there is significant debate about how to best synthesize existing data, evidence exists for specialization based on the kind of processing and on the nature of the information being processed, as well along the anatomical dimensions of left-right, dorsal-ventral, and anterior-posterior (Badre & D’Esposito 2009; Botvinick 2008; Courtney 2004; D’Esposito et al. 2000; Fuster 1997; Koechlin & Hyafil 2007; Petrides 2000; Smith & Jonides 1998; Wager & Smith 2003). Neuroanatomical specialization is broadly consistent with the behavioral evidence for separable components of executive control (Friedman & Miyake 2004; Friedman et al. 2006; Miyake et al. 2000). Such specialization implies that the degree to which engaging in a difficult task affects performance on a subsequent task might depend on the degree to which the two tasks tap similar executive functions and engage similar prefrontal regions. This idea, which has not been systematically explored (although see Persson et al. 2007), contrasts sharply with the notion that carryover effects are uniform across diverse tasks that all tap a unitary “self-control” mechanism (Muraven & Baumeister 2000).
Specialization could also contribute to increasing opportunity costs as more prefrontal neurons are recruited to a given task. Suppose prefrontal neurons can be used for several processes but are best suited for specific processes (by virtue of their connectivity, for example), and the “best-suited” neurons are recruited to a task first. Then the marginal opportunity costs will increase as more neurons are recruited to a task, because the neurons recruited “at the margin” are less and less effective at the current task and more and more effective at other tasks (Just et al. 1999).
4.3.4. Links between executive and motivational circuits
Because tasks that are associated with mental effort all engage a prefrontal executive network, a cost/benefit account requires some mechanism by which neural signals regarding costs and benefits can modulate the performance of this executive network. Although this question has not been widely studied, there are two potential links between prefrontal executive circuits and the motivational circuits discussed above. These links mirror the two mechanisms discussed above for making trade-offs regarding physical effort.
One possibility is that dopamine levels in the prefrontal cortex reflect opportunity costs, similar to proposals regarding dopamine levels in the striatum (Niv et al. 2007). There are direct projections from dopaminergic neurons to the lateral prefrontal cortex, and classic studies from Goldman-Rakic and colleagues (Goldman-Rakic 1996; Goldman-Rakic et al. 2000) demonstrate that the stability of prefrontal activity is a function of local dopamine levels. Given other evidence linking the stability of prefrontal activity to performance (Funahashi et al. 1989), this provides one possible mechanism through which signals about recent reward history could strengthen or weaken prefrontal engagement on the current task (Braveret al. 1999). Aston-Jones and Cohen (2005) proposed a similar idea, arguing that norepinephrine rather than dopamine provides the critical signal regarding the benefit of continued engagement.
Another possibility is that the anterior cingulate cortex functions to link executive and motivational circuits. As discussed above, the anterior cingulate carries signals regarding various costs, such as physical effort, during reward-based decision-making tasks. The anterior cingulate is also part of the prefrontal executive network. In executive function tasks, the anterior cingulate has been associated with monitoring information-processing conflicts. Botvinick (2007) has proposed that these two roles share the same general performance-monitoring function: Information-processing conflicts serve as a negative feedback signal that promotes more efficient task performance in the same way that various other costs serve as signals that promote changes in task performance.
4.4. Neural changes accompanying changes in mental effort and performance
4.4.1. Neural signals related to the subjective cost of mental effort
There have been a limited number of functional imaging studies that have explicitly focused on the phenomenology of mental effort. McGuire and Botvinick (2010) used a paradigm in which subjects had to switch between two tasks, judging the magnitude or parity (odd/even) of single digits. Behaviorally, the frequency of task-switches was associated with a greater self-reported sense of cognitive demand, and people avoided high-demand in favor of low-demand versions of the task when given the opportunity (Kool et al. 2010; McGuire & Botvinick 2010). Rewards after high-demand blocks were also associated with decreased activity in the ventral striatum, consistent with the notion that cognitive demand is costly (Botvinick et al. 2009). Across two further fMRI experiments using this task, bilateral activity in the lateral prefrontal cortex was correlated across blocks with subjective ratings of demand (controlling for objective differences, such as reaction times and errors) and across subjects with the behavioral tendency to avoid high-demand versions of the task. These results suggest that activity in lateral prefrontal regions during demanding cognitive tasks is associated with a subjective cost, and that this cost can motivate subsequent task avoidance.
4.4.2. Neural changes accompanying performance decrements
Other functional imaging studies are of interest because they examine the neural changes that accompany performance decrements. Although small in number, the studies offer a consistent picture: Decrements in performance are associated with decreased engagement of prefrontal regions associated with executive function.
Three published studies have examined neural activity with functional imaging during prolonged (>20 min) sustained attention tasks (Coull et al. 1998; Lim et al. 2010; Paus et al. 1997). All three studies found a vigilance decrement (i.e., increase in reaction times with time-on-task) and an associated decrease in right lateral prefrontal activity over the course of the task. This region has previously been implicated in sustained attention processes (Posner & Petersen 1990).
Two studies have used fMRI to examine neural activity associated with performance decrements in two-task carryover paradigms (Hedgcock et al. 2012; Persson & Reuter-Lorenz 2010). Although the tasks used in these studies differed greatly, both reported that activity in a lateral prefrontal region was greater when the first task was more difficult, and that this same lateral prefrontal region exhibited less activity during the second task when this was preceded by the more difficult initial task. Interestingly, the region of the lateral prefrontal cortex showing this effect was different in the two studies (left inferior frontal gyrus vs. right middle frontal gyrus), consistent with neuroanatomical specialization within the lateral prefrontal cortex.
Several additional studies have examined the neural correlates of performance decrements using event-related potentials. These studies have focused on the error-related negativity (ERN), which is believed to index anterior cingulate activity related to task monitoring. Inzlicht and Gutsell (2007) found that the ERN in a Stroop task was smaller after suppressing emotional responses to a sad movie clip, compared with freely expressing emotion. A similar decrease in the ERN has been observed with sustained (2-hr) performance of an effortful cognitive task (Boksem et al. 2006; Lorist et al. 2005). Importantly, these changes in anterior cingulate activity, like the concomitant behavioral decrements, can be reversed by providing additional incentives for performance (Boksem et al. 2006). Such responsiveness to incentives is consistent with the proposal that the anterior cingulate tracks costs and benefits; it also shows that brain activity, like performance, does not decrease in an obligatory manner with sustained effort.
4.4.3. Distinguishing resource and cost/benefit accounts
The findings in the preceding two sections do not, in themselves, distinguish between resource versus cost/benefit accounts of mental effort and performance. Decreased activity in lateral and dorsomedial prefrontal regions could be due to the depletion of a physical resource necessary for continued high levels of activity, or it could reflect a decision to engage these regions to a lesser degree given the costs and benefits of performance. Lateral prefrontal activity might be associated with a subjective cost because it expends a physical resource, or because it comes with a substantial opportunity cost – precluding any other task that would require the same neural processes.
However, a computational account seems more likely to explain both these results and others regarding these brain regions within a common framework. Previous studies have demonstrated that these regions exhibit changes in neural activity linked to changes in performance on a much faster timescale. For example, in the Stroop task, subjects are generally faster to respond to incongruent trials when the previous trial was also incongruent. Kerns and colleagues (Kerns 2006; Kerns et al. 2004) demonstrated that the size of this sequential adjustment effect was associated with trial-to-trial changes in anterior cingulate and lateral prefrontal activity, specifically (1) greater anterior cingulate activity on the previous trial and (2) greater dorsolateral prefrontal activity on the current trial. These results are consistent with the hypothesis of Botvinick et al. (2001) that the anterior cingulate monitors for information-processing conflicts, which then triggers the subsequent recruitment of lateral prefrontal regions in order to reduce these conflicts.
Note that this hypothesis has the same structure as the one we propose. The anterior cingulate cortex encodes a cost (here, the information-processing conflicts that result from low cognitive control), and lateral prefrontal activity and associated performance adjust accordingly (here, activity increases and performance improves). The direction of the changes in prefrontal activity and performance differ from our proposal, although Botvinick (2007) has already taken the first steps to incorporate both kinds of adjustments in one computational model. Trial-to-trial changes also present a difficulty for resource accounts in that they demonstrate increased lateral prefrontal activity and better performance subsequent to a difficult trial. If performance were determined only by the level of a resource, and this resource can only go down during the task, then such trial-to-trial improvements in task performance should not be possible.
4.5. Summary of neurophysiology
There is little neurophysiological evidence consistent with a resource account of mental effort and performance. Existing evidence does not support the claim that glucose is the physical resource. Effortful tasks do not reliably reduce glucose, things that do reliably reduce glucose such as exercise improve performance on cognitive tasks, and the beneficial effects of glucose on cognitive performance are due to its rewarding properties rather than its caloric content (Kurzban 2010b). Although there could be other potential candidate resources besides glucose, there is no other mature theory of the resource; in particular, there is no theory of the resource that can explain why some kinds of mental activity but not others are effortful.
In contrast, there is abundant neurophysiological evidence consistent with a cost/benefit account of mental effort and performance. A cost/benefit model first requires that the brain encode costs and benefits in a way that integrates across very different kinds of costs and very different kinds of benefits. A ventromedial prefrontal-ventral striatal network encodes such signals (Kable & Glimcher 2009; Lee et al. 2007; Rangel et al. 2008; Rangel & Hare 2010). A cost/benefit model also requires that there be neural processes that (1) can be used for a variety of different tasks and (2) have a limited capacity at any one point in time. A lateral prefrontal “executive” network fulfills these two criteria and is engaged by effortful mental tasks (Duncan 2001; Miller & Cohen 2001). Finally, a cost/benefit model requires a way for cost/benefit signals to influence the engagement of the limited capacity network, and we point to recent proposals describing how feedback signals in the anterior cingulate cortex, or dopamine levels in the prefrontal cortex, could achieve this (Botvinick 2007; Niv et al. 2007). This integrated proposal is consistent with the available evidence regarding neural activity during effortful tasks and performance reductions. Lateral prefrontal regions are engaged by effortful tasks, their engagement is accompanied by the sensation of mental effort, reductions in lateral prefrontal activity accompany reductions in task performance, and changes in lateral prefrontal activity are preceded by feedback signals about costs and benefits in the anterior cingulate.
The above proposal leans heavily on existing computational models describing how these same neural mechanisms calibrate the expenditure of physical effort (Niv et al. 2007) or modulate lateral prefrontal performance in response to information-processing costs (Botvinick et al. 2001), as well as on recent efforts to extend these models to the domain of mental effort (Botvinick 2007; Botvinick et al. 2009; Kool et al. 2010; McGuire & Botvinick 2010). Although these models are well known in cognitive psychology and cognitive neuroscience, they appear to have had little influence on theorizing regarding some of the paradigms we focus on here, such as the vigilance decrement in sustained attention and the reduction in task performance in the dual-task paradigm. Clearly, extending these models to these domains is possible, and likely to be a fruitful enterprise.
5. Conclusion
Some, perhaps even many or most, phenomenological experiences are reasonably easy to understand from a functional perspective. The positive, rewarding sensations of behaviors ranging from sexual activity (Diamond 1997) to coming to be valued by others (Leary et al. 1995) can be understood as the output of motivational systems designed to bring about adaptive behavior. These positive sensations correspond in a reasonably straightforward way to behaviors related to fitness gains. To the extent that phenomenology is understood as part of the motivational system, driving organisms toward good fitness outcomes, many experiences – especially the valence of these experiences – make a great deal of sense.
In this context, the phenomenology of effort presents something of a puzzle. Many of the real-world tasks that evoke a sensation of effort lead to favorable outcomes in the long run – persisting on difficult tasks such as writing, doing math problems, and so on – yet the phenomenology is unpleasant rather than pleasant. Further, these sensations seem to be systematically related to performance reductions. Why do these “good” things feel “bad”?
We have tried to sketch one sort of solution to this puzzle. The central element of our argument is that the sensation of effort is designed around a particular adaptive problem and its solution, simultaneity and prioritization. Because some systems, especially those associated with executive function, have multiple uses to which they can be put, the use of these systems carries opportunity costs. We propose that these costs are experienced as “effort,” and have the effect of reducing task performance. This connects the sensation of effort to other qualia, explaining the valence of the experience as a cost of persisting.
We also want to emphasize that our explanation is, of course, not wholly novel. Dodge (1917), for example, suggested the subjective experience of fatigue had to do with subjects’ desire to attend to something other than the task before them, and the general idea of fatigue as a problem of choosing to do what one ought can be traced back perhaps still further (Thorndike 1904). We have similarly tried to acknowledge throughout areas where our view overlaps, sometimes in substantial part, those of others (Boksem et al. 2005; 2006; Boksem & Tops 2008; Botvinick 2007; Hockey 2011; Kool et al. 2010; Lorist et al. 2005; McGuire & Botvinick 2010; van der Linden 2011).
Finally, we wish to point out that to some extent, the literatures on “self-control” in psychology and “executive function” in cognitive psychology and neuroscience have not been as tightly integrated as they could be, and part of our agenda in writing this piece is to highlight that these streams of research should be in closer contact with each other. Whether or not our particular computational explanation for these effects turns out to be correct, some computational explanation will eventually be required, and our hope is that this paper moves closer to that goal.
Acknowledgments
We thank Geoffrey Goodwin for his immensely valuable input in preparing this manuscript. Angela Duckworth’s contribution was supported by grant K01-AG033182 from the National Institute on Aging. Joe Kable’s contribution to this article was partially supported by NIH grant DA029149.
Contributor Information
Robert Kurzban, Email: kurzban@psych.upenn.edu.
Angela Duckworth, Email: duckworth@psych.upenn.edu.
Joseph W. Kable, Email: kable@psych.upenn.edu.
Justus Myers, Email: justusm@psych.upenn.edu.
References
[Robert Kurzban, Angela Duckworth, Joseph W. Kable, and Justus Myers] [aRK]
- Ackerman JM, Goldstein NJ, Shapiro JR, Bargh JA. You wear me out: The vicarious depletion of self-control. Psychological Science. 2009;20(3):326–32. doi: 10.1111/j.1467-9280.2009.02290.x. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ackerman PL. 100 years without resting. In: Ackerman PL, editor. Cognitive fatigue: Multidisciplinary perspectives on current research and future applications. American Psychological Association; 2011. pp. 11–43. [aRK] [Google Scholar]
- Akerlof GA, Yellen JL. The fair wage–effort hypothesis and unemployment. The Quarterly Journal of Economics. 1990;105(2):255–83. [aRK] [Google Scholar]
- Alexander GE, Crutcher MD. Functional architecture of basal ganglia circuits: Neural substrates of parallel processing. Trends in Neurosciences. 1990;13(7):266–71. doi: 10.1016/0166-2236(90)90107-l. [aRK] [DOI] [PubMed] [Google Scholar]
- Arai T. Mental fatigue. Teachers College, Columbia University; 1912. [aRK] [Google Scholar]
- Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: Adaptive gain and optimal performance. Annual Review of Neuroscience. 2005;28:403–50. doi: 10.1146/annurev.neuro.28.061604.135709. [aRK] [DOI] [PubMed] [Google Scholar]
- Avena NM, Rada P, Hoebel BG. Evidence for sugar addiction: Behavioral and neurochemical effects of intermittent, excessive sugar intake. Neuroscience and Biobehavioral Reviews. 2008;32(1):20–39. doi: 10.1016/j.neubiorev.2007.04.019. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baddeley AD, Hitch G. Working memory. In: Bower GA, editor. Recent advances in learning and motivation. Vol. 8. Academic Press; 1974. pp. 47–90. [aRK] [Google Scholar]
- Badre D, D’Esposito M. Is the rostro-caudal axis of the frontal lobe hierarchical? Nature Reviews Neuroscience. 2009;10(9):659–69. doi: 10.1038/nrn2667. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barsh GS, Schwartz MW. Genetic approaches to studying energy balance: Perception and integration. Nature Reviews Genetics. 2002;3(8):589–600. doi: 10.1038/nrg862. [aRK] [DOI] [PubMed] [Google Scholar]
- Baumeister RF, Bratslavsky E, Muraven M, Tice DM. Ego depletion: Is the active self a limited resource? Journal of Personality and Social Psychology. 1998;74(5):1252–65. doi: 10.1037//0022-3514.74.5.1252. [aRK] [DOI] [PubMed] [Google Scholar]
- Baumeister RF, Vohs KD. Self-regulation, ego depletion, and motivation. Social and Personality Psychology Compass. 2007;1(1):115–28. [aRK] [Google Scholar]
- Baumeister RF, Vohs KD, Tice DM. The strength model of self-control. Current Directions in Psychological Science. 2007;16(6):351–55. [aRK] [Google Scholar]
- Beedie CJ, Lane AM. The role of glucose in self-control. Personality and Social Psychology Review. 2012;16(2):143–53. doi: 10.1177/1088868311419817. [aRK] [DOI] [PubMed] [Google Scholar]
- Berridge KC. Motivation concepts in behavioral neuroscience. Physiology & Behavior. 2004;81(2):179–209. doi: 10.1016/j.physbeh.2004.02.004. [aRK] [DOI] [PubMed] [Google Scholar]
- Berridge KC. The debate over dopamine’s role in reward: The case for incentive salience. Psychopharmacology. 2007;191(3):391–431. doi: 10.1007/s00213-006-0578-x. [aRK] [DOI] [PubMed] [Google Scholar]
- Bloom P. How pleasure works: The new science of why we like what we like. W. W. Norton; 2010. [aRK] [Google Scholar]
- Boksem MAS, Meijman TF, Lorist MM. Effects of mental fatigue on attention: An ERP study. Cognitive Brain Research. 2005;25(1):107–16. doi: 10.1016/j.cogbrainres.2005.04.011. [aRK] [DOI] [PubMed] [Google Scholar]
- Boksem MAS, Meijman TF, Lorist MM. Mental fatigue, motivation, and action monitoring. Biological Psychology. 2006;72(2):123–32. doi: 10.1016/j.biopsycho.2005.08.007. [aRK] [DOI] [PubMed] [Google Scholar]
- Boksem MAS, Tops M. Mental fatigue: Costs and benefits. Brain Research Reviews. 2008;59(1):125–39. doi: 10.1016/j.brainresrev.2008.07.001. [aRK] [DOI] [PubMed] [Google Scholar]
- Botvinick MM. Conflict monitoring and decision making: Reconciling two perspectives on anterior cingulate function. Cognitive, Affective, & Behavioral Neuroscience. 2007;7(4):356–66. doi: 10.3758/cabn.7.4.356. [aRK] [DOI] [PubMed] [Google Scholar]
- Botvinick MM. Hierarchical models of behavior and prefrontal function. Trends in Cognitive Science. 2008;12(5):201–208. doi: 10.1016/j.tics.2008.02.009. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botvinick MM, Braver TS, Barch DM, Carter CS, Cohen JD. Conflict monitoring and cognitive control. Psychological Review. 2001;108(3):624–52. doi: 10.1037/0033-295x.108.3.624. [aRK] [DOI] [PubMed] [Google Scholar]
- Botvinick MM, Huffstetler S, McGuire JT. Effort discounting in human nucleus accumbens. Cognitive, Affective, & Behavioral Neuroscience. 2009;9(1):16–27. doi: 10.3758/CABN.9.1.16. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botvinick M, Jha AP, Bylsma LM, Fabian SA, Solomon PE, Prkachin KM. Viewing facial expressions of pain engages cortical areas involved in the direct experience of pain. NeuroImage. 2005;25(1):312–19. doi: 10.1016/j.neuroimage.2004.11.043. [aRK] [DOI] [PubMed] [Google Scholar]
- Brain WR. Mind, perception, and science. Blackwell; 1951. [aRK] [Google Scholar]
- Braver TS, Barch DM, Cohen JD. Cognition and control in schizophrenia: A computational model of dopamine and prefrontal function. Biological Psychiatry. 1999;46(3):312–28. doi: 10.1016/s0006-3223(99)00116-x. [aRK] [DOI] [PubMed] [Google Scholar]
- Buchsbaum BR, Greer S, Chang WL, Berman KF. Meta-analysis of neuroimaging studies of the Wisconsin card-sorting task and component processes. Human Brain Mapping. 2005;25(1):35–45. doi: 10.1002/hbm.20128. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnham T, Phelan J. Mean genes: From sex to money to food: Taming our primal instincts. Perseus; 2000. [aRK] [Google Scholar]
- Bush G, Vogt BA, Holmes J, Dale AM, Greve D, Jenike MA, Rosen BR. Dorsal anterior cingulate cortex: A role in reward-based decision making. Proceedings of the National Academy of Sciences. 2002;99(1):523–28. doi: 10.1073/pnas.012470999. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buss DM. Sex differences in human mate preferences: Evolutionary hypotheses tested in 37 cultures. Behavioral and Brain Sciences. 1989;12(1):1–14. [aRK] [Google Scholar]
- Buss DM, Larsen RJ, Westen D, Semmelroth J. Sex differences in jealousy: Evolution, physiology, and psychology. Psychological Science. 1992;3(4):251–55. [aRK] [Google Scholar]
- Cabeza R, Nyberg L. Imaging cognition II: An empirical review of 275 PET and fMRI studies. Journal of Cognitive Neuroscience. 2000;12(1):1–47. doi: 10.1162/08989290051137585. [aRK] [DOI] [PubMed] [Google Scholar]
- Camille N, Griffiths CA, Vo K, Fellows LK, Kable JW. Ventromedial frontal lobe damage disrupts value maximization in humans. Journal of Neuroscience. 2011;31(20):7527–32. doi: 10.1523/JNEUROSCI.6527-10.2011. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambers ES, Bridge MW, Jones DA. Carbohydrate sensing in the human mouth: Effects on exercise performance and brain activity. The Journal of Physiology. 2009;587(8):1779–94. doi: 10.1113/jphysiol.2008.164285. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charnov EL. Optimal foraging, the marginal value theorem. Theoretical Population Biology. 1976;9(2):129–36. doi: 10.1016/0040-5809(76)90040-x. [aRK] [DOI] [PubMed] [Google Scholar]
- Cialdini RB, Schaller M, Houlihan D, Arps K, Fultz J, Beaman AL. Empathy-based helping: Is it selflessly or selfishly motivated? Journal of Personality and Social Psychology. 1987;52:749–58. doi: 10.1037//0022-3514.52.4.749. [aRK] [DOI] [PubMed] [Google Scholar]
- Clarke DD, Sokoloff L. Circulation and energy metabolism of the brain. In: Siegel GJ, Agranoff BW, Albers RW, Molinoff PB, editors. Basic neurochemistry: Molecular, cellular, and medical aspects. 6. Lippincott-Raven; 1998. pp. 637–69. [aRK] [Google Scholar]
- Cohen JD, McClure SM, Yu AJ. Should I stay or should I go? How the human brain manages the trade-off between exploitation and exploration. Philosophical Transactions of the Royal Society B: Biological Sciences. 2007;362(1481):933–42. doi: 10.1098/rstb.2007.2098. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Converse PD, DeShon RP. A tale of two tasks: Reversing the self-regulatory resource depletion effect. Journal of Applied Psychology. 2009;94(5):1318–24. doi: 10.1037/a0014604. [aRK] [DOI] [PubMed] [Google Scholar]
- Coull JT, Frackowiak RS, Frith CD. Monitoring for target objects: Activation of right frontal and parietal cortices with increasing time on task. Neuropsychologia. 1998;36(12):1325–34. doi: 10.1016/s0028-3932(98)00035-9. [aRK] [DOI] [PubMed] [Google Scholar]
- Courtney SM. Attention and cognitive control as emergent properties of information representation in working memory. Cognitive, Affective, & Behavioral Neuroscience. 2004;4(4):501–16. doi: 10.3758/cabn.4.4.501. [aRK] [DOI] [PubMed] [Google Scholar]
- Croxson PL, Walton ME, O’Reilly JX, Behrens TE, Rushworth MF. Effort-based cost-benefit valuation and the human brain. Journal of Neuroscience. 2009;29(14):4531–41. doi: 10.1523/JNEUROSCI.4515-08.2009. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- D’Ardenne K, McClure SM, Nystrom LE, Cohen JD. BOLD responses reflecting dopaminergic signals in the human ventral tegmental area. Science. 2008;319(5867):1264–67. doi: 10.1126/science.1150605. [aRK] [DOI] [PubMed] [Google Scholar]
- D’Esposito M, Postle BR, Rypma B. Prefrontal cortical contributions to working memory: Evidence from event-related fMRI studies. Experimental Brain Research. 2000;133(1):3–11. doi: 10.1007/s002210000395. [aRK] [DOI] [PubMed] [Google Scholar]
- Damasio AR. The feelings of what happens: Body and emotion in the making of consciousness. Houghton Mifflin Harcourt; 1999. [aRK] [Google Scholar]
- Davies DR, Parasuraman R. The psychology of vigilance. Academic Press; 1982. [aRK] [Google Scholar]
- Dehaene S, Spelke E, Pinel P, Stanescu R, Tsivkin S. Sources of mathematical thinking: Behavioral and brain-imaging evidence. Science. 1999;284(5416):970–74. doi: 10.1126/science.284.5416.970. [aRK] [DOI] [PubMed] [Google Scholar]
- Derrfuss J, Brass M, Neumann J, von Cramon DY. Involvement of the inferior frontal junction in cognitive control: Meta-analyses of switching and Stroop studies. Human Brain Mapping. 2005;25(1):22–34. doi: 10.1002/hbm.20127. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desimone R, Duncan J. Neural mechanisms of selective visual attention. Annual Review of Neuroscience. 1995;18:193–222. doi: 10.1146/annurev.ne.18.030195.001205. [aRK] [DOI] [PubMed] [Google Scholar]
- DeWall CN, Baumeister RF, Stillman TF, Gailliot MT. Violence restrained: Effects of self-regulation and its depletion on aggression. Journal of Experimental Social Psychology. 2007;43(1):62–76. [aRK] [Google Scholar]
- Dewitte S, Bruyneel S, Geyskens K. Self-regulating enhances self-regulation in subsequent consumer decisions involving similar response conflicts. Journal of Consumer Research. 2009;36(3):394–405. [aRK] [Google Scholar]
- Diamond J. Why is sex fun? The evolution of human sexuality. Basic Books; 1997. [aRK] [Google Scholar]
- Dodge R. The laws of relative fatigue. Psychological Review. 1917;24(2):89–113. [aRK] [Google Scholar]
- Duncan J. An adaptive coding model of neural function in prefrontal cortex. Nature Reviews Neuroscience. 2001;2(11):820–29. doi: 10.1038/35097575. [aRK] [DOI] [PubMed] [Google Scholar]
- Eisenberger R. Learned industriousness. Psychological Review. 1992;99(2):248–67. doi: 10.1037/0033-295x.99.2.248. [aRK] [DOI] [PubMed] [Google Scholar]
- Eisenberger R, Masterson FA. Required high effort increases subsequent persistence and reduces cheating. Journal of Personality and Social Psychology. 1983;44(3):593–99. [aRK] [Google Scholar]
- Evans SBT. Dual-processing accounts of reasoning, judgment, and social cognition. Annual Review of Psychology. 2008;59:255–78. doi: 10.1146/annurev.psych.59.103006.093629. [aRK] [DOI] [PubMed] [Google Scholar]
- Fairclough SH, Houston K. A metabolic measure of mental effort. Biological Psychology. 2004;66(2):177–90. doi: 10.1016/j.biopsycho.2003.10.001. [aRK] [DOI] [PubMed] [Google Scholar]
- Fan J, McCandliss BD, Sommer T, Raz A, Posner MI. Testing the efficiency and independence of attentional networks. Journal of Cognitive Neuroscience. 2002;14(3):340–47. doi: 10.1162/089892902317361886. [aRK] [DOI] [PubMed] [Google Scholar]
- Fehr E, Goette L, Zehnder C. A behavioral account of the labor market: The role of fairness concerns. Annual Review of Economics. 2009;1:355–84. [aRK] [Google Scholar]
- Fellows LK, Farah MJ. The role of ventromedial prefrontal cortex in decision making: Judgment under uncertainty or judgment per se? Cerebral Cortex. 2007;17(11):2669–74. doi: 10.1093/cercor/bhl176. [aRK] [DOI] [PubMed] [Google Scholar]
- Fliessbach K, Weber B, Trautner P, Dohmen T, Sunde U, Elger CE, Falk A. Social comparison affects reward-related brain activity in the human ventral striatum. Science. 2007;318(5854):1305–08. doi: 10.1126/science.1145876. [aRK] [DOI] [PubMed] [Google Scholar]
- Fodor JA. The modularity of mind: An essay on faculty psychology. MIT Press; 1983. [aRK] [Google Scholar]
- Freedman DJ, Riesenhuber M, Poggio T, Miller EK. Categorical representation of visual stimuli in the primate prefrontal cortex. Science. 2001;291(5502):312–16. doi: 10.1126/science.291.5502.312. [aRK] [DOI] [PubMed] [Google Scholar]
- Friedman NP, Miyake A. The relations among inhibition and interference control functions: A latent-variable analysis. Journal of Experimental Psychology:General. 2004;133(1):101–35. doi: 10.1037/0096-3445.133.1.101. [aRK] [DOI] [PubMed] [Google Scholar]
- Friedman NP, Miyake A, Corley RP, Young SE, Defries JC, Hewitt JK. Not all executive functions are related to intelligence. Psychological Science. 2006;17(2):172–79. doi: 10.1111/j.1467-9280.2006.01681.x. [aRK] [DOI] [PubMed] [Google Scholar]
- Funahashi S, Bruce CJ, Goldman-Rakic PS. Mnemonic coding of visual space in the monkey’s dorsolateral prefrontal cortex. Journal of Neurophysiology. 1989;61(2):331–49. doi: 10.1152/jn.1989.61.2.331. [aRK] [DOI] [PubMed] [Google Scholar]
- Fuster JM. Network memory. Trends in Neuroscience. 1997;20(10):451–59. doi: 10.1016/s0166-2236(97)01128-4. [aRK] [DOI] [PubMed] [Google Scholar]
- Gailliot MT, Baumeister RF. The physiology of willpower: Linking blood glucose to self-control. Personality and Social Psychology Review. 2007;11(4):303–27. doi: 10.1177/1088868307303030. [aRK] [DOI] [PubMed] [Google Scholar]
- Gailliot MT, Baumeister RF, DeWall CN, Maner JK, Plant EA, Tice DM, Schmeichel BJ. Self-control relies on glucose as a limited energy source: Willpower is more than a metaphor. Journal of Personality and Social Psychology. 2007;92(2):325–36. doi: 10.1037/0022-3514.92.2.325. [aRK] [DOI] [PubMed] [Google Scholar]
- Gallistel CR. The organization of learning: Learning, development, and conceptual change. MIT Press; 1990. [aRK] [Google Scholar]
- Gibson EL. Carbohydrates and mental function: Feeding or impeding the brain? Nutrition Bulletin. 2007;32(suppl I):71–83. [aRK] [Google Scholar]
- Gibson EL, Green MW. Nutritional influences on cognitive function: Mechanisms of susceptibility. Nutrition Research Reviews. 2002;15(1):169–206. doi: 10.1079/NRR200131. [aRK] [DOI] [PubMed] [Google Scholar]
- Gilbert DT, Wilson TD. Prospection: Experiencing the future. Science. 2007;317(5843):1351–54. doi: 10.1126/science.1144161. [aRK] [DOI] [PubMed] [Google Scholar]
- Glimcher PW. Decisions, uncertainty and the brain: The science of neuroeconomics. MIT Press; 2003. [aRK] [Google Scholar]
- Goel V. Anatomy of deductive reasoning. Trends in Cognitive Science. 2007;11(10):435–41. doi: 10.1016/j.tics.2007.09.003. [aRK] [DOI] [PubMed] [Google Scholar]
- Goldman-Rakic PS. Regional and cellular fractionation of working memory. Proceedings of the National Academy of Sciences. 1996;93(24):13473–80. doi: 10.1073/pnas.93.24.13473. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldman-Rakic PS, Muly EC, III, Williams GV. D(1) receptors in prefrontal cells and circuits. Brain Research Reviews. 2000;31(2–3):295–301. doi: 10.1016/s0165-0173(99)00045-4. [aRK] [DOI] [PubMed] [Google Scholar]
- Gonzalez C, Best B, Healy AF, Kole JA, Bourne LE., Jr A cognitive modeling account of simultaneous learning and fatigue effects. Cognitive Systems Research. 2011;12:19–32. [aRK] [Google Scholar]
- Gopher D, Brickner M, Navon D. Different difficulty manipulations interact differently with task emphasis: Evidence for multiple resources. Journal of Experimental Psychology: Human Perception and Performance. 1982;8:146–57. doi: 10.1037//0096-1523.8.1.146. [aRK] [DOI] [PubMed] [Google Scholar]
- Gopher D, Navon D. How is performance limited: Testing the notion of central capacity. Acta Psychologica. 1980;46:161–80. [aRK] [Google Scholar]
- Grill HJ, Kaplan JM. The neuroanatomical axis for control of energy balance. Frontiers in Neuroendocrinology. 2002;23(1):2–40. doi: 10.1006/frne.2001.0224. [aRK] [DOI] [PubMed] [Google Scholar]
- Gunzelmann G, Gross JB, Gluck KA, Dinges DF. Sleep deprivation and sustained attention performance: Integrating mathematical and cognitive modeling. Cognitive Science. 2009;33:880–910. doi: 10.1111/j.1551-6709.2009.01032.x. [aRK] [DOI] [PubMed] [Google Scholar]
- Haber SN. The primate basal ganglia: Parallel and integrative networks. Journal of Chemical Neuroanatomy. 2003;26(4):317–30. doi: 10.1016/j.jchemneu.2003.10.003. [aRK] [DOI] [PubMed] [Google Scholar]
- Haber SN, Knutson B. The reward circuit: Linking primate anatomy and human imaging. Neuropsychopharmacology. 2009;35(1):4–26. doi: 10.1038/npp.2009.129. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagger MS, Wood C, Stiff C, Chatzisarantis NLD. The strength model of self-regulation failure and health-related behavior. Health Psychology Review. 2009;3(2):208–38. [aRK] [Google Scholar]
- Hagger MS, Wood C, Stiff C, Chatzisarantis NLD. Ego depletion and the strength model of self-control: A meta-analysis. Psychological Bulletin. 2010a;136(4):495–525. doi: 10.1037/a0019486. [aRK] [DOI] [PubMed] [Google Scholar]
- Hagger MS, Wood CW, Stiff C, Chatzisarantis NLD. Self-regulation and self-control in exercise: The strength-energy model. International Review of Sport and Exercise Psychology. 2010b;3(1):62–86. [aRK] [Google Scholar]
- Harbaugh WT, Mayr U, Burghart DR. Neural responses to taxation and voluntary giving reveal motives for charitable donations. Science. 2007;316(5831):1622–25. doi: 10.1126/science.1140738. [aRK] [DOI] [PubMed] [Google Scholar]
- Hare TA, Camerer CF, Knoepfle DT, Rangel A. Value computations in ventral medial prefrontal cortex during charitable decision making incorporate input from regions involved in social cognition. Journal of Neuroscience. 2010;30(2):583–90. doi: 10.1523/JNEUROSCI.4089-09.2010. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hare TA, Camerer CF, Rangel A. Self-control in decision-making involves modulation of the vmPFC valuation system. Science. 2009;324(5927):646–48. doi: 10.1126/science.1168450. [aRK] [DOI] [PubMed] [Google Scholar]
- Hayden BY, Pearson JM, Platt ML. Fictive reward signals in the anterior cingulate cortex. Science. 2009;324(5929):948–50. doi: 10.1126/science.1168488. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Head H. The conception of nervous and mental energy (II). Vigilance: A physiological state of the nervous system. British Journal of Psychology. 1923;14:126–47. [aRK] [Google Scholar]
- Healy AF, Kole JA, Buck-Gengler CJ, Bourne LE. Effects of prolonged work on data entry speed and accuracy. Journal of Experimental Psychology: Applied. 2004;10:188–99. doi: 10.1037/1076-898X.10.3.188. [aRK] [DOI] [PubMed] [Google Scholar]
- Hedgcock WM, Vohs KD, Rao AR. Reducing self-control depletion effects through enhanced sensitivity to implementation: Evidence from fMRI and behavioral studies. Journal of Consumer Psychology. 2012;22(4):486–95. [aRK] [Google Scholar]
- Hein G, Silani G, Preuschoff K, Batson CD, Singer T. Neural responses to ingroup and outgroup members’ suffering predict individual differences in costly helping. Neuron. 2010;68(1):149–60. doi: 10.1016/j.neuron.2010.09.003. [aRK] [DOI] [PubMed] [Google Scholar]
- Hickman KL, Stromme C, Lippman LG. Learned industriousness: Replication in principle. Journal of General Psychology. 1998;125(3):213–17. doi: 10.1080/00221309809595545. [aRK] [DOI] [PubMed] [Google Scholar]
- Hillman CH, Erickson KI, Kramer AF. Be smart, exercise your heart: Exercise effects on brain and cognition. Nature Reviews Neuroscience. 2008;9(1):58–65. doi: 10.1038/nrn2298. [aRK] [DOI] [PubMed] [Google Scholar]
- Hillman CH, Pontifex MB, Raine LB, Castelli DM, Hall EE, Kramer AF. The effect of acute treadmill walking on cognitive control and academic achievement in preadolescent children. Neuroscience. 2009;159(3):1044–54. doi: 10.1016/j.neuroscience.2009.01.057. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hockey GRJ. A motivational control theory of cognitive fatigue. In: Ackerman PL, editor. Cognitive fatigue: Multidisciplinary perspectives on current research and future applications. American Psychological Association; 2011. pp. 167–87. [aRK] [Google Scholar]
- Holding DH. Fatigue. In: Hockey G, editor. Stress and fatigue in human performance. Wiley; 1983. pp. 145–67. [aRK] [Google Scholar]
- Hommel JD, Trinko R, Sears RM, Georgescu D, Liu Z, Gao X, DiLeone RJ. Leptin receptor signaling in midbrain dopamine neurons regulates feeding. Neuron. 2006;51(6):801–10. doi: 10.1016/j.neuron.2006.08.023. [aRK] [DOI] [PubMed] [Google Scholar]
- Huxtable ZL, White MH, McCartor MA. A re-performance and re-interpretation of the Arai Experiment in mental fatigue with three subjects. Psychological Monographs. 1946;59(5):52. [aRK] [Google Scholar]
- Inzlicht M, Gutsell JN. Running on empty: Neural signals for self-control failure. Psychological Science. 2007;18(11):933–37. doi: 10.1111/j.1467-9280.2007.02004.x. [aRK] [DOI] [PubMed] [Google Scholar]
- James JM, Bolstein R. Large monetary incentives and their effect on mail survey response rates. Public Opinion Quarterly. 1992;56(4):442–53. [aRK] [Google Scholar]
- Jeukendrup AE, Chambers ES. Oral carbohydrate sensing and exercise performance. Curr Opin Clin Nutr Metab Care. 2010;13(4):447–51. doi: 10.1097/MCO.0b013e328339de83. [aRK] [DOI] [PubMed] [Google Scholar]
- Job V, Dweck CS, Walton GM. Ego depletion – Is it all in your head? Implicit theories about willpower affect self-regulation. Psychological Science. 2010;21(11):1686–93. doi: 10.1177/0956797610384745. [aRK] [DOI] [PubMed] [Google Scholar]
- Just MA, Carpenter PA, Varma S. Computational modeling of high-level cognition and brain function. Human Brain Mapping. 1999;8:128–36. doi: 10.1002/(SICI)1097-0193(1999)8:2/3<128::AID-HBM10>3.0.CO;2-G. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kable JW, Glimcher PW. The neural correlates of subjective value during intertemporal choice. Nature Neuroscience. 2007;10(12):1625–33. doi: 10.1038/nn2007. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kable JW, Glimcher PW. The neurobiology of decision: Consensus and controversy. Neuron. 2009;63(6):733–45. doi: 10.1016/j.neuron.2009.09.003. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahneman D. Attention and effort. Prentice-Hall; 1973. [aRK] [Google Scholar]
- Kanfer R. Determinants and consequences of subjective cognitive fatigue. In: Ackerman PL, editor. Cognitive fatigue: Multidisciplinary perspectives on current research and future applications. American Psychological Association; 2011. pp. 189–207. [aRK] [Google Scholar]
- Kennerley SW, Dahmubed AF, Lara AH, Wallis JD. Neurons in the frontal lobe encode the value of multiple decision variables. Journal of Cognitive Neuroscience. 2009;21(6):1162–78. doi: 10.1162/jocn.2009.21100. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerns JG. Anterior cingulate and prefrontal cortex activity in an fMRI study of trial-to-trial adjustments on the Simon task. NeuroImage. 2006;33(1):399–405. doi: 10.1016/j.neuroimage.2006.06.012. [aRK] [DOI] [PubMed] [Google Scholar]
- Kerns JG, Cohen JD, MacDonald AW, Cho RY, Stenger VA, Carter CS. Anterior cingulate conflict monitoring and adjustments in control. Science. 2004;303(5660):1023–26. doi: 10.1126/science.1089910. [aRK] [DOI] [PubMed] [Google Scholar]
- Kirkpatrick LA, Ellis BJ. An evolutionary-psychological approach to self-esteem: Multiple domains and multiple functions. In: Fletcher GJO, Clark MS, editors. Blackwell handbook of social psychology: Interpersonal processes. Blackwell; 2001. pp. 52–77. [aRK] [Google Scholar]
- Kirkpatrick LA, Waugh CE, Valencia A, Webster GD. The functional domain specificity of self-esteem and the differential prediction of aggression. Journal of Personality and Social Psychology. 2002;82(5):756–67. [aRK] [PubMed] [Google Scholar]
- Klucharev V, Hytonen K, Rijpkema M, Smidts A, Fernandez G. Reinforcement learning signal predicts social conformity. Neuron. 2009;61(1):140–51. doi: 10.1016/j.neuron.2008.11.027. [aRK] [DOI] [PubMed] [Google Scholar]
- Koechlin E, Hyafil A. Anterior prefrontal function and the limits of human decision-making. Science. 2007;318(5850):594–98. doi: 10.1126/science.1142995. [aRK] [DOI] [PubMed] [Google Scholar]
- Kool W, McGuire JT, Rosen ZB, Botvinick MM. Decision making and the avoidance of cognitive demand. Journal of Experimental Psychology: General. 2010;139(4):665–82. doi: 10.1037/a0020198. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krebs RM, Boehler CN, Woldorff MG. The influence of reward associations on conflict processing in the Stroop task. Cognition. 2010;117(3):341–47. doi: 10.1016/j.cognition.2010.08.018. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhnen CM, Knutson B. The neural basis of financial risk taking. Neuron. 2005;47(5):763–70. doi: 10.1016/j.neuron.2005.08.008. [aRK] [DOI] [PubMed] [Google Scholar]
- Kurniawan IT, Guitart-Masip M, Dolan RJ. Dopamine and effort-based decision making. Frontiers in Neuroscience. 2011;5:1–10. doi: 10.3389/fnins.2011.00081. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurniawan IT, Seymour B, Talmi D, Yoshida W, Chater N, Dolan RJ. Choosing to make an effort: The role of striatum in signaling physical effort of a chosen action. Journal of Neurophysiology. 2010;104(1):313–21. doi: 10.1152/jn.00027.2010. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurzban R. The Psychology of Joss Whedon. Dallas, TX: BenBella Books; 2007. Mal’s Morals; pp. 7–19. [aRK] [Google Scholar]
- Kurzban R. Does the brain consume additional glucose during self-control tasks? Evolutionary Psychology. 2010a;8(2):244–59. [aRK] [PubMed] [Google Scholar]
- Kurzban R. Why everyone (else) is a hypocrite: Evolution and the modular mind. Princeton University Press; 2010b. [aRK] [Google Scholar]
- Kurzban R. Cheatin’ hearts and loaded guns: The high fitness stakes of country music lyrics. Review of General Psychology (Special Issue: The Intersection of Evolution and Popular Culture) 2012;16:187–91. [aRK] [Google Scholar]
- Laird AR, McMillan KM, Lancaster JL, Kochunov P, Turkeltaub PE, Pardo JV, Fox PT. A comparison of label-based review and ALE meta-analysis in the Stroop task. Human Brain Mapping. 2005;25(1):6–21. doi: 10.1002/hbm.20129. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laran J, Janiszewski C. Work or fun? How task construal and completion influence regulatory behavior. Journal of Consumer Research. 2011;37(6):967–83. [aRK] [Google Scholar]
- Lau B, Glimcher PW. Value representations in the primate striatum during matching behavior. Neuron. 2008;58(3):451–63. doi: 10.1016/j.neuron.2008.02.021. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazarus RS. From psychological stress to the emotions: A history of changing outlooks. Annual Review of Psychology. 1993;44(1):1–22. doi: 10.1146/annurev.ps.44.020193.000245. [aRK] [DOI] [PubMed] [Google Scholar]
- Leary MR, Baumeister RF. The nature and function of self-esteem: Sociometer theory. Advances in Experimental Social Psychology. 2000;32:1–62. [aRK] [Google Scholar]
- Leary MR, Tambor ES, Terdal SK, Downs DL. Self-esteem as an interpersonal monitor: The sociometer hypothesis. Journal of Personality and Social Psychology. 1995;68(3):518–30. [aRK] [Google Scholar]
- Lee D, Rushworth MF, Walton ME, Watanabe M, Sakagami M. Functional specialization of the primate frontal cortex during decision making. Journal of Neuroscience. 2007;27(31):8170–73. doi: 10.1523/JNEUROSCI.1561-07.2007. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim J, Wu WC, Wang J, Detre JA, Dinges DF, Rao H. Imaging brain fatigue from sustained mental workload: An ASL perfusion study of the time-on-task effect. NeuroImage. 2010;49(4):3426–35. doi: 10.1016/j.neuroimage.2009.11.020. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lorist MM, Boksem MAS, Ridderinkhof KR. Impaired cognitive control and reduced cingulate activity during mental fatigue. Cognitive Brain Research. 2005;24(2):199–205. doi: 10.1016/j.cogbrainres.2005.01.018. [aRK] [DOI] [PubMed] [Google Scholar]
- Lorist MM, Klein M, Nieuwenhuis S, Jong R, Mulder G, Meijman TF. Mental fatigue and task control: Planning and preparation. Psychophysiology. 2000;37(5):614–25. [aRK] [PubMed] [Google Scholar]
- Lynn M, McCall M. Unpublished manuscript. School of Hotel Administration, Cornell University; 2000. Beyond gratitude and gratuity: A meta-analytic review of the predictors of restaurant tipping. [aRK] [Google Scholar]
- Mackworth N. The breakdown of vigilance during prolonged visual search. The Quarterly Journal of Experimental Psychology. 1948;1:6–21. [aRK] [Google Scholar]
- Magen E, Gross JJ. Harnessing the need for immediate gratification: Cognitive reconstrual modulates the reward value of temptations. Emotion. 2007;7(2):415–28. doi: 10.1037/1528-3542.7.2.415. [aRK] [DOI] [PubMed] [Google Scholar]
- Marr D. Vision: A computational investigation into the human representation and processing of visual information. W. H. Freeman; 1982. [aRK] [Google Scholar]
- Martijn C, Tenbult P, Merckelbach H, Dreezens E, de Vries NK. Getting a grip on ourselves: Challenging expectancies about loss of energy after self-control. Social Cognition. 2002;20(6):441–60. [aRK] [Google Scholar]
- Masicampo EJ, Baumeister RF. Toward a physiology of dual-process reasoning and judgment: Lemonade, willpower, and expensive rule-based analysis. Psychological Science. 2008;19(3):255–60. doi: 10.1111/j.1467-9280.2008.02077.x. [aRK] [DOI] [PubMed] [Google Scholar]
- McClure SM, Berns GS, Montague PR. Temporal prediction errors in a passive learning task activate human striatum. Neuron. 2003;38(2):339–46. doi: 10.1016/s0896-6273(03)00154-5. [aRK] [DOI] [PubMed] [Google Scholar]
- McGuire JT, Botvinick MM. Prefrontal cortex, cognitive control, and the registration of decision costs. Proceedings of the National Academy of Science. 2010;107(17):7922–26. doi: 10.1073/pnas.0910662107. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messier C. Glucose improvement of memory: A review. European Journal of Pharmacology. 2004;490(1–3):33–57. doi: 10.1016/j.ejphar.2004.02.043. [aRK] [DOI] [PubMed] [Google Scholar]
- Miller EK, Cohen JD. An integrative theory of prefrontal cortex function. Annual Review of Neuroscience. 2001;24(1):167–202. doi: 10.1146/annurev.neuro.24.1.167. [aRK] [DOI] [PubMed] [Google Scholar]
- Miller GA. The magical number seven, plus or minus two: Some limits on our capacity for processing information. Psychological Review. 1956;63(2):81–97. [aRK] [PubMed] [Google Scholar]
- Minsky M. The society of mind. Simon & Schuster; 1985. [aRK] [Google Scholar]
- Mitchell JP, Heatherton TF, Kelley WM, Wyland CL, Wegner DM, Neil Macrae C. Separating sustained from transient aspects of cognitive control during thought suppression. Psychological Science. 2007;18(4):292–97. doi: 10.1111/j.1467-9280.2007.01891.x. [aRK] [DOI] [PubMed] [Google Scholar]
- Miyake A, Friedman NP, Emerson MJ, Witzki AH, Howerter A, Wager TD. The unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: A latent variable analysis. Cognitive Psychology. 2000;41(1):49–100. doi: 10.1006/cogp.1999.0734. [aRK] [DOI] [PubMed] [Google Scholar]
- Molden DC, Hui CM, Scholer AA, Meier BP, Noreen EE, D’Agostino PR, Martin V. The motivational versus metabolic effects of carbohydrates on self-control. Psychological Science. 2012;23(10):1137–44. doi: 10.1177/0956797612439069. [aRK] [DOI] [PubMed] [Google Scholar]
- Montague PR, Dayan P, Sejnowski TJ. A framework for mesencephalic dopamine systems based on predictive Hebbian learning. Journal of Neuroscience. 1996;16(5):1936–47. doi: 10.1523/JNEUROSCI.16-05-01936.1996. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moray N. Where is capacity limited? A survey and a model. Acta Psychologica. 1967;27:84–92. doi: 10.1016/0001-6918(67)90048-0. [aRK] [DOI] [PubMed] [Google Scholar]
- Muraven M, Baumeister RF. Self-regulation and depletion of limited resources: Does self-control resemble a muscle? Psychological Bulletin. 2000;126(2):247–59. doi: 10.1037/0033-2909.126.2.247. [aRK] [DOI] [PubMed] [Google Scholar]
- Muraven M, Shmueli D, Burkley E. Conserving self-control strength. Journal of Personality and Social Psychology. 2006;91(3):524–37. doi: 10.1037/0022-3514.91.3.524. [aRK] [DOI] [PubMed] [Google Scholar]
- Muraven M, Slessareva E. Mechanisms of self-control failure: Motivation and limited resources. Personality and Social Psychology Bulletin. 2003;29(7):894–906. doi: 10.1177/0146167203029007008. [aRK] [DOI] [PubMed] [Google Scholar]
- Muraven M, Tice DM, Baumeister RF. Self-control as a limited resource: Regulatory depletion patterns. Journal of Personality and Social Psychology. 1998;74(3):774–89. doi: 10.1037//0022-3514.74.3.774. [aRK] [DOI] [PubMed] [Google Scholar]
- Naccache L, Dehaene S, Cohen L, Habert MO, Guichart-Gomez E, Galanaud D, Willer J. Effortless control: Executive attention and conscious feeling of mental effort are dissociable. Neuropsychologia. 2005;43(9):1318–28. doi: 10.1016/j.neuropsychologia.2004.11.024. [aRK] [DOI] [PubMed] [Google Scholar]
- Nassi JJ, Callaway EM. Parallel processing strategies of the primate visual system. Nature Reviews Neuroscience. 2009;10(5):360–72. doi: 10.1038/nrn2619. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navon D. Resources – A theoretical soup stone? Psychological Review. 1984;91(2):216–34. [aRK] [Google Scholar]
- Navon D, Gopher D. On the economy of the human-processing system. Psychological Review. 1979;86(3):214–55. [aRK] [Google Scholar]
- Nee DE, Wager TD, Jonides J. Interference resolution: Insights from a meta-analysis of neuroimaging tasks. Cognitive, Affective, & Behavioral Neuroscience. 2007;7(1):1–17. doi: 10.3758/cabn.7.1.1. [aRK] [DOI] [PubMed] [Google Scholar]
- Neumann J, Lohmann G, Derrfuss J, von Cramon DY. Meta-analysis of functional imaging data using replicator dynamics. Human Brain Mapping. 2005;25(1):165–73. doi: 10.1002/hbm.20133. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newberg AB, Wang J, Rao H, Swanson RL, Wintering N, Karp JS, Detre JA. Concurrent CBF and CMRG1c changes during human brain activation by combined fMRI-PET scanning. NeuroImage. 2005;28(2):500–506. doi: 10.1016/j.neuroimage.2005.06.040. [aRK] [DOI] [PubMed] [Google Scholar]
- Nieder A, Dehaene S. Representation of number in the brain. Annal Review of Neuroscience. 2009;32:185–208. doi: 10.1146/annurev.neuro.051508.135550. [aRK] [DOI] [PubMed] [Google Scholar]
- Niv Y, Daw ND, Joel D, Dayan P. Tonic dopamine: Opportunity costs and the control of response vigor. Psychopharmacology (Berl) 2007;191(3):507–20. doi: 10.1007/s00213-006-0502-4. [aRK] [DOI] [PubMed] [Google Scholar]
- Niv Y, Joel D, Dayan P. A normative perspective on motivation. Trends in Cognitive Sciences. 2006;10:375–81. doi: 10.1016/j.tics.2006.06.010. [aRK] [DOI] [PubMed] [Google Scholar]
- Nix GA, Ryan RM, Manly JB, Deci EL. Revitalization through self-regulation: The effects of autonomous and controlled motivation on happiness and vitality. Journal of Experimental Social Psychology. 1999;35(3):266–84. [aRK] [Google Scholar]
- O’Connell KA, Schwartz JE, Shiffman S. Do resisted temptations during smoking cessation deplete or augment self-control resources? Psychology of Addictive Behaviors. 2008;22(4):486–95. doi: 10.1037/0893-164X.22.4.486. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Doherty JP, Dayan P, Friston K, Critchley H, Dolan RJ. Temporal difference models and reward-related learning in the human brain. Neuron. 2003;38(2):329–37. doi: 10.1016/s0896-6273(03)00169-7. [aRK] [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Gross JJ. The cognitive control of emotion. Trends in Cognitive Sciences. 2005;9(5):242–49. doi: 10.1016/j.tics.2005.03.010. [aRK] [DOI] [PubMed] [Google Scholar]
- Orians GH, Heerwagen JH. Evolved responses to landscapes. In: Barkow JH, Cosmides L, Tooby J, editors. The adapted mind: Evolutionary psychology and the generation of culture. Oxford University Press; 1992. pp. 555–79. [aRK] [Google Scholar]
- Orne MT. On the social psychology of the psychological experiment: With particular reference to demand characteristics and their implications. American Psychologist. 1962;17(11):776–83. [aRK] [Google Scholar]
- Padoa-Schioppa C, Assad JA. Neurons in the orbitofrontal cortex encode economic value. Nature. 2006;441(7090):223–26. doi: 10.1038/nature04676. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Padoa-Schioppa C, Assad JA. The representation of economic value in the orbitofrontal cortex is invariant for changes of menu. Nature Neuroscience. 2008;11(1):95–102. doi: 10.1038/nn2020. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paus T, Zatorre RJ, Hofle N, Caramanos Z, Gotman J, Petrides M, Evans AC. Time-related changes in neural systems underlying attention and arousal during the performance of an auditory vigilance task. Journal of Cognitive Neuroscience. 1997;9(3):392–408. doi: 10.1162/jocn.1997.9.3.392. [aRK] [DOI] [PubMed] [Google Scholar]
- Persson J, Reuter-Lorenz P. Training and depletion of executive functions: The case of interference control. Paper presented at the Cognitive Neuroscience Society Annual Meeting; Montreal, Quebec. April 17–20, 2010; 2010. [aRK] [Google Scholar]
- Persson J, Welsh KM, Jonides J, Reuter-Lorenz PA. Cognitive fatigue of executive processes: Interaction between interference resolution tasks. Neuropsychologia. 2007;45(7):1571–79. doi: 10.1016/j.neuropsychologia.2006.12.007. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pessiglione M, Seymour B, Flandin G, Dolan RJ, Frith CD. Dopamine-dependent prediction errors underpin reward-seeking behaviour in humans. Nature. 2006;442(7106):1042–45. doi: 10.1038/nature05051. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrides M. The role of the mid-dorsolateral prefrontal cortex in working memory. Experimental Brain Research. 2000;133(1):44–54. doi: 10.1007/s002210000399. [aRK] [DOI] [PubMed] [Google Scholar]
- Pinker S. How the mind works. W. W. Norton; 1997. [aRK] [Google Scholar]
- Plassmann H, O’Doherty J, Rangel A. Orbitofrontal cortex encodes willingness to pay in everyday economic transactions. Journal of Neuroscience. 2007;27(37):9984–88. doi: 10.1523/JNEUROSCI.2131-07.2007. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Posner MI, Petersen SE. The attention system of the human brain. Annual Review of Neuroscience. 1990;13:25–42. doi: 10.1146/annurev.ne.13.030190.000325. [aRK] [DOI] [PubMed] [Google Scholar]
- Posner MI, Snyder CRR. Attention and cognitive control. In: Solso RL, editor. Information processing and cognition: The Loyola Symposium. Erlbaum; 1975. pp. 55–85. [aRK] [Google Scholar]
- Posner MI, Snyder CRR, Davidson BJ. Attention and the detection of signals. Journal of Experimental Psychology: General. 1980;109:160–74. [aRK] [PubMed] [Google Scholar]
- Prevost C, Pessiglione M, Metereau E, Clery-Melin ML, Dreher JC. Separate valuation subsystems for delay and effort decision costs. Journal of Neuroscience. 2010;30(42):14080–90. doi: 10.1523/JNEUROSCI.2752-10.2010. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raichle ME, Gusnard DA. Appraising the brain’s energy budget. Proceedings of the National Academy of Sciences. 2002;99(16):10237–39. doi: 10.1073/pnas.172399499. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proceedings of the National Academy of Sciences. 2001;98(2):676–82. doi: 10.1073/pnas.98.2.676. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rainer G, Asaad WF, Miller EK. Selective representation of relevant information by neurons in the primate prefrontal cortex. Nature. 1998;393(6685):577–79. doi: 10.1038/31235. [aRK] [DOI] [PubMed] [Google Scholar]
- Rangel A, Camerer C, Montague PR. A framework for studying the neurobiology of value-based decision making. Nature Reviews Neuroscience. 2008;9(7):545–56. doi: 10.1038/nrn2357. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rangel A, Hare T. Neural computations associated with goal-directed choice. Current Opinion in Neurobiology. 2010;20(2):262–70. doi: 10.1016/j.conb.2010.03.001. [aRK] [DOI] [PubMed] [Google Scholar]
- Rao SC, Rainer G, Miller EK. Integration of what and where in the primate prefrontal cortex. Science. 1997;276(5313):821–24. doi: 10.1126/science.276.5313.821. [aRK] [DOI] [PubMed] [Google Scholar]
- Regan DT. Effects of a favor and liking on compliance. Journal of Experimental Social Psychology. 1971;7(6):627–39. [aRK] [Google Scholar]
- Robinson MD, Schmeichel BJ, Inzlicht M. A cognitive control perspective of self-control strength and its depletion. Social and Personality Psychology Compass. 2010;4(3):189–200. [aRK] [Google Scholar]
- Rolfe JM. The secondary task as a measure of mental load. In: Singleton WT, Easterby RS, Whitfield DE, editors. Measurement of man at work. Taylor & Francis; 1971. pp. 135–48. [aRK] [Google Scholar]
- Rousselet GA, Fabre-Thorpe M, Thorpe SJ. Parallel processing in high-level categorization of natural images. Nature Neuroscience. 2002;5:629–30. doi: 10.1038/nn866. [aRK] [DOI] [PubMed] [Google Scholar]
- Rozin P, Vollmecke TA. Food likes and dislikes. Annual Review of Nutrition. 1986;6(1):433–56. doi: 10.1146/annurev.nu.06.070186.002245. [aRK] [DOI] [PubMed] [Google Scholar]
- Rudebeck PH, Behrens TE, Kennerley SW, Baxter MG, Buckley MJ, Walton ME, Rushworth FS. Frontal cortex subregions play distinct roles in choices between actions and stimuli. The Journal of Neuroscience. 2008;28(51):13775–85. doi: 10.1523/JNEUROSCI.3541-08.2008. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudebeck PH, Walton ME, Smyth AN, Bannerman DM, Rushworth MF. Separate neural pathways process different decision costs. Nature Neuroscience. 2006;9(9):1161–68. doi: 10.1038/nn1756. [aRK] [DOI] [PubMed] [Google Scholar]
- Rummelhart DE, McClelland JL The PDP Research Group. Parallel distributed processing: Explorations in the microstructure of cognition. Vol. 1. MIT Press; 1986. [aRK] [DOI] [PubMed] [Google Scholar]
- Rutledge RB, Lazzaro SC, Lau B, Myers CE, Gluck MA, Glimcher PW. Dopaminergic drugs modulate learning rates and perseveration in Parkinson’s patients in a dynamic foraging task. Journal of Neuroscience. 2009;29(48):15104–14. doi: 10.1523/JNEUROSCI.3524-09.2009. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salamone JD, Correa M, Farrar AM, Nunes EJ, Pardo M. Dopamine, behavioral economics, and effort. Frontiers in Behavioral Neuroscience. 2009;3:13. doi: 10.3389/neuro.08.013.2009. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samejima K, Ueda Y, Doya K, Kimura M. Representation of action-specific reward values in the striatum. Science. 2005;310(5752):1337–40. doi: 10.1126/science.1115270. [aRK] [DOI] [PubMed] [Google Scholar]
- Scerbo MW. Stress, workload, and boredom in vigilance: A problem and an answer. In: Hancock PA, Desmond PA, editors. Stress, workload, and fatigue. Lawrence Erlbaum; 2001. pp. 267–78. [aRK] [Google Scholar]
- Scerbo MW, Holcomb JR. Effects of signal conspicuity and time on boredom in vigilance. Paper presented at the First Mid-Atlantic Human Factors Conference; Virginia Beach, Virginia. February 25–26, 1993; 1993. [aRK] [Google Scholar]
- Schimmack U. The ironic effect of significant results on the credibility of multiple study articles. Psychological Methods. doi: 10.1037/a0029487. in press. [aRK] [DOI] [PubMed] [Google Scholar]
- Schmeichel BJ. Attention control, memory updating, and emotion regulation temporarily reduce the capacity for executive control. Journal of Experimental Psychology: General. 2007;136:241–55. doi: 10.1037/0096-3445.136.2.241. [aRK] [DOI] [PubMed] [Google Scholar]
- Schmeichel BJ, Vohs K. Self-affirmation and self-control: Affirming core values counteracts ego depletion. Journal of Personality and Social Psychology. 2009;96:770–82. doi: 10.1037/a0014635. [aRK] [DOI] [PubMed] [Google Scholar]
- Schneider F, Gur RE, Alavi A, Seligman MEP, Mozley LH, Smith RJ, Mozley PD, Gur RC. Cerebral blood flow changes in limbic regions induced by unsolvable anagram tasks. American Journal of Psychiatry. 1996;153(2):206–12. doi: 10.1176/ajp.153.2.206. [aRK] [DOI] [PubMed] [Google Scholar]
- Scholey AB, Harper S, Kennedy DO. Cognitive demand and blood glucose. Physiology & Behavior. 2001;73(4):585–92. doi: 10.1016/s0031-9384(01)00476-0. [aRK] [DOI] [PubMed] [Google Scholar]
- Schultz W, Dayan P, Montague PR. A neural substrate of prediction and reward. Science. 1997;275(5306):1593–99. doi: 10.1126/science.275.5306.1593. [aRK] [DOI] [PubMed] [Google Scholar]
- Sigman M, Dehaene S. Brain mechanisms of serial and parallel processing during dual-task performance. The Journal of Neuroscience. 2008;28(30):7585–98. doi: 10.1523/JNEUROSCI.0948-08.2008. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singer T, Seymour B, O’Doherty JP, Kaube H, Dolan RJ, Frith CD. Empathy for pain involves the affective but not sensory components of pain. Science. 2004;303(5661):1157–62. doi: 10.1126/science.1093535. [aRK] [DOI] [PubMed] [Google Scholar]
- Singer T, Seymour B, O’Doherty JP, Stephan KE, Dolan RJ, Frith CD. Empathic neural responses are modulated by the perceived fairness of others. Nature. 2006;439(7075):466–69. doi: 10.1038/nature04271. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh D. Adaptive significance of female physical attractiveness: Role of waist-to-hip ratio. Journal of Personality and Social Psychology. 1993;65(2):293–307. doi: 10.1037//0022-3514.65.2.293. [aRK] [DOI] [PubMed] [Google Scholar]
- Smith EE, Jonides J. Neuroimaging analyses of human working memory. Proceedings of the National Academy of Sciences. 1998;95(20):12061–68. doi: 10.1073/pnas.95.20.12061. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sperber D. The modularity of thought and the epidemiology of representations. In: Hirschfeld L, Gelman SA, editors. Mapping the mind: Domain specificity in cognition and culture. Cambridge University Press; 1994. pp. 39–67. [aRK] [Google Scholar]
- Stucke TS, Baumeister RF. Ego depletion and aggressive behavior: Is the inhibition of aggression a limited resource? European Journal of Social Psychology. 2006;36(1):1–13. [aRK] [Google Scholar]
- Stuss DT, Alexander MP. Executive functions and the frontal lobes: A conceptual view. Psychological Research. 2000;63(3):289–98. doi: 10.1007/s004269900007. [aRK] [DOI] [PubMed] [Google Scholar]
- Sutton RS, Barto AG. Reinforcement learning: An introduction. MIT Press; 1998. [aRK] [Google Scholar]
- Symons D. On the use and misuse of Darwinism in the study of human behavior. In: Barkow JH, Cosmides L, Tooby J, editors. The adapted mind: Evolutionary psychology and the generation of culture. Oxford University Press; 1992. pp. 137–59. [aRK] [Google Scholar]
- Thompson-Schill SL, Bedny M, Goldberg RF. The frontal lobes and the regulation of mental activity. Current Opinion in Neurobiology. 2005;15(2):219–24. doi: 10.1016/j.conb.2005.03.006. [aRK] [DOI] [PubMed] [Google Scholar]
- Thorndike EL. An introduction to the theory of mental and social measurements. Science Press; 1904. [aRK] [Google Scholar]
- Thornhill R. Darwinian aesthetics. In: Crawford C, Krebs DL, editors. Handbook of evolutionary psychology: Ideas, issues, and applications. Lawrence Erlbaum; 1998. pp. 543–72. [aRK] [Google Scholar]
- Tice DM, Baumeister RF, Shmueli D, Muraven M. Restoring the self: Positive affect helps improve self-regulation following ego depletion. Journal of Experimental Social Psychology. 2007;43(3):379–84. [aRK] [Google Scholar]
- Tom SM, Fox CR, Trepel C, Poldrack RA. The neural basis of loss aversion in decision-making under risk. Science. 2007;315(5811):515–18. doi: 10.1126/science.1134239. [aRK] [DOI] [PubMed] [Google Scholar]
- Tomporowski PD. Effects of acute bouts of exercise on cognition. Acta Psychologica. 2003;112(3):297–324. doi: 10.1016/s0001-6918(02)00134-8. [aRK] [DOI] [PubMed] [Google Scholar]
- Tooby J, Cosmides L. The psychological foundations of culture. In: Barkow JH, Cosmides L, Tooby J, editors. The adapted mind: Evolutionary psychology and the generation of culture. Oxford University Press; 1992. pp. 19–136. [aRK] [Google Scholar]
- Tooby J, Cosmides L, Sell A, Lieberman D, Sznycer D. Internal regulatory variables and the design of human motivation: A computational and evolutionary approach. In: Elliot A, editor. Handbook of approach and avoidance motivation. Lawrence Erlbaum; 2008. pp. 251–71. [aRK] [Google Scholar]
- Tops M, Lorist MM, Wijers AA, Meijman TF. To stress or relax: Neurochemical aspects of activity and rest. Gedrag en Organisatie. 2004;17:32–42. [aRK] [Google Scholar]
- Tyler JM, Burns KC. After depletion: The replenishment of the self’s regulatory resources. Self and Identity. 2008;7:305–21. [aRK] [Google Scholar]
- van der Linden D. The urge to stop: The cognitive and biological nature of acute mental fatigue. In: Ackerman PL, editor. Cognitive fatigue: Multidisciplinary perspectives on current research and future applications. American Psychological Association; 2011. pp. 149–64. [aRK] [Google Scholar]
- van der Linden D, Frese M, Meijman TF. Mental fatigue and the control of cognitive processes: Effects on perseveration and planning. Acta Psychologica. 2003;113(1):45–65. doi: 10.1016/s0001-6918(02)00150-6. [aRK] [DOI] [PubMed] [Google Scholar]
- Vohs KD, Baumeister RF, Schmeichel BJ, Twenge JM, Nelson NM, Tice DM. Making choices impairs subsequent self-control: A limited-resource account of decision making, self-regulation, and active initiative. Journal of Personality and Social Psychology. 2008;94(5):883–98. doi: 10.1037/0022-3514.94.5.883. [aRK] [DOI] [PubMed] [Google Scholar]
- Wager TD, Smith EE. Neuroimaging studies of working memory: A meta-analysis. Cognitive, Affective, & Behavioral Neuroscience. 2003;3(4):255–74. doi: 10.3758/cabn.3.4.255. [aRK] [DOI] [PubMed] [Google Scholar]
- Walton ME, Bannerman DM, Alterescu K, Rushworth MF. Functional specialization within medial frontal cortex of the anterior cingulate for evaluating effort-related decisions. The Journal of Neuroscience. 2003;23(16):6475–79. doi: 10.1523/JNEUROSCI.23-16-06475.2003. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warm JS. An introduction to vigilance. In: Warm JS, editor. Sustained attention in human performance. Wiley; 1984. pp. 1–14. [aRK] [Google Scholar]
- Warm JS, Matthews G, Finomore VS., Jr . Vigilance, workload, and stress. In: Hancock PA, Szalma J, editors. Performance under stress. Ashgate; 2008. pp. 115–41. [aRK] [Google Scholar]
- Webb TL, Sheeran P. Can implementation intentions help to overcome ego-depletion? Journal of Experimental Social Psychology. 2003;39(3):279–86. [aRK] [Google Scholar]
- Wickens CD. Multiple resources and performance prediction. Theoretical Issues in Ergonomics Science. 2002;3(2):159–77. [aRK] [Google Scholar]
- Williams GC. Adaptation and natural selection. Princeton University Press; 1966. [aRK] [Google Scholar]
- Wright RA, Junious TR, Neal C, Avello A, Graham C, Herrmann L, Junious S, Walton N. Mental fatigue on effort-related cardiovascular response: Difficulty effects and extension across cognitive performance domains. Motivation and Emotion. 2007;31:219–31. [aRK] [Google Scholar]
- Wyland CL, Kelley WM, Macrae CN, Gordon HL, Heatherton TF. Neural correlates of thought suppression. Neuropsychologia. 2003;41(14):1863–67. doi: 10.1016/j.neuropsychologia.2003.08.001. [aRK] [DOI] [PubMed] [Google Scholar]
- Zaghloul KA, Blanco JA, Weidemann CT, McGill K, Jaggi JL, Baltuch GH, Kahana MJ. Human substantia nigra neurons encode unexpected financial rewards. Science. 2009;323(5920):1496–99. doi: 10.1126/science.1167342. [aRK] [DOI] [PMC free article] [PubMed] [Google Scholar]