

Abstract
Purpose
To identify the role of chemokine receptor 6 (D6) expression by dendritic cells (DCs) and its role in corneal transplant immunity.
Methods
Flow cytometry analysis was used to assess the expression level of the D6 chemokine receptor in different leukocyte populations and DC maturation following lipopolysaccharides (LPS) stimulation of bone marrow–derived DCs isolated from wild-type (WT) or D6−/− mice (C57BL/6 background). Mixed-lymphocyte reactions and delayed-type hypersensitivity assays were performed with bone marrow–derived DCs from WT or D6−/− mice to evaluate T-cell alloreactivity. Adoptive transfer experiments with T cells from WT or D6−/− hosts with BALB/c corneal allografts were performed. Graft opacity was assessed over an 8-week period, and graft survival was plotted using Kaplan–Meier survival curves.
Results
Expression of the D6 chemokine receptor was significantly higher in DCs compared to other leukocyte subpopulations, including neutrophils, lymphocytes, and monocytes/macrophages. LPS challenge of D6−/− bone marrow–derived DCs elicited significantly lower levels of major histocompatibility complex II and costimulatory molecules (CD40, CD80, and CD86) compared to WT bone marrow–derived DCs, indicating the role of the D6 chemokine receptor in DC maturation. Further, DCs isolated from D6−/− mice induced less T-cell proliferation (p≤0.001) and interferon-gamma production in T cells of draining lymph nodes compared to WT mice following corneal transplantation (p≤0.001). Moreover, adoptively transferred T cells from D6−/− corneal transplanted mice to WT mice led to impaired graft rejection, compared to the hosts that received T cells from the WT transplanted mice.
Conclusions
We demonstrated D6 chemokine receptor expression by DCs and identified its critical function in multiple aspects of DC biology, including maturation and consequent elicitation of alloreactive T-cell responses that are responsible for corneal allograft rejection.
Introduction
The unique family of receptors known as chemokine “scavenging receptors” was originally defined by their ability to ligate chemokines in a non-signaling fashion. These decoy receptors include chemokine receptor 6 (D6), the Duffy antigen receptor for chemokines (DARC), and CCX-CKR. The best described of these is D6, known to be expressed in placenta [1] and lymphatic endothelial cells [1,2] (in gut, skin, and lung) and capable of scavenging more than 12 chemokines—mostly agonists of inflammatory CC chemokine receptors from CCR1 to CCR5 [1]. Thus, in contrast to conventional chemokine receptors by which chemokine ligation induces leukocyte recruitment in inflammation, ligation via D6 leads to targeted chemokine degradation and consequent reduction of their bioavailability [2-4]. Consistent with this, the absence of D6 expression has been associated with uncontrolled and sustained inflammation in multiple inflammatory models, including mycobacterium tuberculosis infection, phorbol esters–induced cutaneous inflammatory response, and inflammatory bowel disease [3,5-7]. This has led to the theory that expression of D6 by lymphatic endothelial cells plays a crucial role in mediating resolution of inflammation.
Recent data indicate that in addition to lymphatic endothelial cells, various populations of immune cells, including B cells, macrophages, and dendritic cells, express the D6 chemokine receptor [8-12]. Interestingly, and in contrast to its function when expressed in the lymphatic endothelium, D6 expression on leukocytes has been reported to promote inflammatory responses in some experimental models [8,9,12-14]. One key report came from Liu et al., who demonstrated that D6−/− mice develop milder forms of experimental autoimmune encephalomyelitis compared to their wild-type (WT) counterparts, which was mediated by an impaired T-cell response [14]. Thus, an alternate role for D6 via leukocyte expression that potentially involves the elicitation of adaptive responses has been suggested.
In the current study, we examined the specific function of D6 expression on corneal allograft rejection, which is mediated primarily by alloreactive CD4+ T cells. We accomplished this by using the T-cell adoptive transfer assay, similarly described by Liu et al. [14], using D6−/− mice in corneal transplantation to evaluate the effect of D6 absence on immune rejection. Here, we demonstrate that host D6 expression actually amplifies alloreactive T-cell responses that mediate immune rejection. Furthermore, we have identified a positive contribution of D6 expression to DCs promoting their maturation, including upregulation of major histocompatibility complex (MHC) II and costimulatory (CD80/86, CD40) molecules. Taken together, our data identify a novel role for D6 in mediating alloimmune responses by promoting the maturation of dendritic cells (DCs) and allosensitization.
Methods
Animals
Male 6- to 8-week-old C57BL/6 and BALB/c mice (Taconic Farms, Germantown, NY) were used. C57BL/6 D6 null mice were kindly provided by Dr. Donald Cook (NIH/NIEHS) and bred in house at the Schepens Eye Research Institute (Boston, MA) animal facility. The institutional Animal Care and Use Committee approved all procedures. All animals were treated according to the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research.
Flow cytometry
The following antibodies (Abs) were used: fluorescein isothiocyanate (FITC)-conjugated rat anti-mouse CD11b (monocyte/macrophage marker), FITC-conjugated hamster anti-mouse CD11c (dendritic cell marker), allophycocyanin (APC)-conjugated rat anti-mouse CD45 (leukocyte common marker), FITC-conjugated hamster anti-mouse CD80 (costimulatory molecule, B7–1), FITC-conjugated rat anti-mouse CD86 (costimulatory molecule, B7–2), FITC-conjugated rat anti-mouse GR-1 (neutrophil marker), FITC-conjugated rat anti-mouse CD8, Phycoerythrin (PE)-conjugated mouse anti-mouse IAb (major histocompatibility complex class II), and goat anti-mouse D6 (Abcam, Cambridge, MA). Secondary Abs were rhodamine-conjugated donkey anti-goat immunoglobulin G (IgG), FITC-conjugated donkey anti-rabbit IgG, and FITC-conjugated goat anti-rabbit IgG. All primary and secondary Abs (except where noted) and isotype matched controls were purchased from BD PharMingen (San Diego, CA). To avoid non-specific staining, cells were blocked with anti-FcR mAb (CD16/CD32) for 15 min at 4 °C before they were stained with primary antibodies or isotype-matched control antibodies (BD PharMingen).
Cultivation of bone marrow–derived dendritic cells
Bone marrow–derived DCs were generated as described elsewhere [15]. Briefly, 6- to 8-week-old WT or D6−/− mice were scarified. The femurs were collected, and bone marrow cells were flushed with 28G syringes and seeded (2×106) in 10 ml Petri dishes with cultured medium (RPMI-1640 supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 100 U/ml of penicillin, 100 µg/ml of streptomycin, 50 mM 2-ME) containing 20 ng/ml of granulocyte-macrophage colony stimulating factor (GM-CSF; PeproTech, Rocky Hill, NJ). On days 3 and 6, new medium containing GM-CSF (20 ng/ml) was added, and non-adherent and loosely adherent immature DCs were collected on day 8. To induce maturation, 100 ng/ml of lipopolysaccharides (LPS; R&D Systems, Minneapolis, MN) was added 24 h before the cells were harvested.
Mixed leukocyte reaction
CD90+ splenic T cells (105/well) from BALB/c mice were MACS sorted (Miltenyi Biotec, Auburn, CA) and cocultured at 1:1 with mitomycin C–treated (50 μg/ml for 30 min at 37 °C; Sigma-Aldrich, St. Louis, MO) bone marrow-derived dendritic cells (BMDC) or splenic antigen presenting cells (APC; CD90-) for 4 days in 96-well round-bottom plates. After 3 days, the bromodeoxyuridine (BrdU) reagent (Sigma-Aldrich) was added for approximately 16 h. Then, the plates were centrifuged, and the medium was removed. Following fixation and denaturation of each well, the plates were washed and incubated with peroxidase-anti-BrdU conjugate solution at room temperature for 60 min. After washing, the horseradish peroxidase substrate was added, and optical density (OD) was analyzed at 450 nm using a microplate reader (MRX, Dynatech, Chantilly, VA).
Local adoptive transfer assay
BALB/c mice received a subcutaneous injection into the nape of the neck containing C57BL/6 splenocytes (105). After 7 days, lymph node (LN) T cells (105) from these mice were coinjected at a 1:1 ratio with WT or D6−/− BMDCs into the ear pinnae of the naïve BALB/c mice. Local adoptive transfer (LAT) assay was also performed with T cells from the C57BL/6 WT or D6−/− mice 3 weeks post-transplantation with BALB/c donors. T cells (105) were coinjected at a ratio of 1:1 with BALB/c APCs into the ear pinnae of naïve C57BL/6 mice. LAT assays were performed twice, and each group consisted of n=5 mice. In both LAT assays, ear thickness was measured 48 h after cells were injected in a masked fashion using a micrometer (Mitutoyo; MTI, Paramus, NJ) [16].
Corneal transplantation and adoptive transfer
The corneal transplant protocol we used here has been described elsewhere [16]. Briefly, the central cornea (2 mm diameter) was marked by a trephine and excised from a BALB/c or C57BL/6 donor mouse using Vannas scissors (Storz Instruments, San Dimas, CA), and then placed on ice in Optisol-GS (Bausch and Lomb, Rochester, NY). The graft bed was prepared by excising a 1.5-mm site in the central cornea of a C57BL/6 WT or D6−/− recipient mouse. A donor button was then placed onto the graft bed and secured with eight interrupted 11–0 nylon sutures (Sharpoint, Reading, PA). For the adoptive transfer experiments, splenic and draining lymph node T cells were isolated from D6−/− or WT mice 3 weeks after transplantation with BALB/c cornea, and 1×106 T cells were injected intravenously into C57BL/6 WT transplanted (BALB/c cornea) mice. Graft survival was evaluated twice a week over the course of 8 weeks using a slit-lamp biomicroscope. Experiments were performed twice with five mice per group. We used the following standardized opacity-grading (ranges 0–5+): 0=clear graft; 1+=minimal superficial (nonstromal) opacity; 2+=minimal deep stromal opacity; 3+=moderate deep stromal opacity; 4+=intense deep stromal opacity; 5+=maximum stromal opacity. Rejection is defined as a score of 2+ or greater for two consecutive examinations.
Cauterization-induced corneal inflammation
Thermal cauterization, a non-antigen-specific acute stimulation, was applied to the mouse corneal surface of either D6−/− mice or C57BL/6 WT mice to induce mobilization of inflammatory cells into corneal tissue. Under anesthesia, six light burns were applied to the central 50% of the cornea, using the tip of a hand-held thermal cautery (Aaron Medical Industries, St. Petersburg, FL). Afterward, bacitracin-neomycin-polymyxin (Vetropolycin; Dechra Veterinary Products, Overland Park, KS) ophthalmic ointment was applied. Corneas were excised 7 days after being cauterized, harvested, and wholemount corneas were stained with CD11c to enumerate dendritic cell infiltration.
Statistical analysis
Statistical analyses including the Student t test were performed throughout the study, and are indicated in the respective figure legends. Error bars displayed in the figures were calculated from the standard error of the mean (±SEM). Kaplan–Meier analysis constructed survival curves and respective log-rank tests compared the rates of corneal graft survival. P values of <0.05 were considered significant.
Results
D6 expression is required for phenotypic maturation of dendritic cells
Using flow cytometry, we examined D6 expression in different leukocyte populations. D6 expression was detected in CD4+ and CD8+ T cells, CD11b+ leukocytes (primarily composed of macrophages), Gr-1high leukocytes (primarily comprised of neutrophils), and CD11c+ DCs. Interestingly, DCs were found to express the highest level of D6 among leukocyte subpopulations (Figure 1A), consistent with several previous reports [8-12,17].
Figure 1.
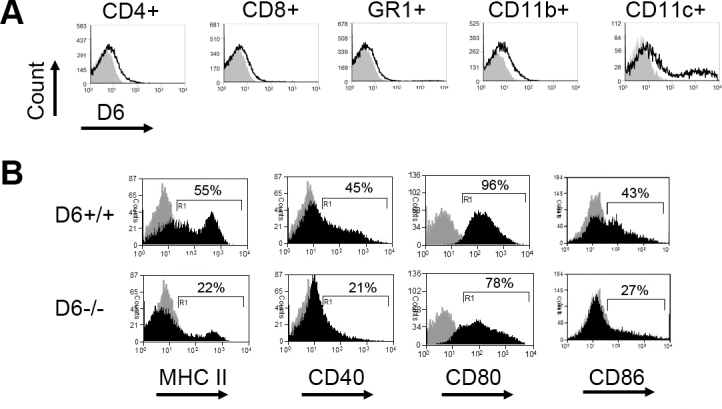
D6 chemokine receptor expression by dendritic cells promotes their maturation. A: Flow cytometry analysis of whole spleen cells for D6 expression on CD4+ and CD8+ T cells, Gr-1high neutrophils, CD11b+ macrophages, and CD11c+ dendritic cells (DCs). The black line represents D6, the gray area represents the isotype control. D6 expression was higher in DCs compared to other leukocyte populations. B: Flow cytometry analysis of bone marrow–derived DCs for major histocompatibility complex (MHC) II and co-stimulatory molecules, CD40, CD80, and CD86 (black area) versus the isotype control (gray area) is shown. Bone marrow–derived DCs from D6−/− and wild-type (WT; D6+/+) mice were analyzed 24 h after lipopolysaccharides (LPS; 100 ng/ml) treatment.
Next, we hypothesized that the robust expression of D6 chemokine receptor by DCs suggests important functionality in DCs. Antigen presentation and T-cell priming are two principal aspects of mature DC function, both enabled by upregulation of MHC II and costimulatory molecules. Thus, we next evaluated the function of the D6 chemokine receptor in phenotypic maturation (acquisition of high expression of MHC II and costimulatory factors) of DCs by comparing the maturation capacity of bone marrow–derived DCs from WT versus D6−/− mice in response to in vitro LPS stimulation. As expected, LPS elicited marked upregulation of MHC II (I-Ab) and costimulatory molecules, i.e., CD40, CD80, and CD86, in bone marrow–derived DCs isolated from WT mice. However, the bone marrow–derived DCs isolated from D6 chemokine receptor null mice demonstrated defective upregulation of all maturation markers assessed in our study (Figure 1B). Taken together, these findings demonstrate that DCs express high levels of D6 chemokine receptor, which is associated with higher phenotypic maturation.
D6 expression by dendritic cells promotes T-cell allosensitization
Because maturation of DCs is critical to their role in priming alloreactive T cells, we next investigated whether D6-mediated maturation of DCs promotes T-cell allosensitization. A modified mixed leukocyte reaction assay was used, in which DCs from C57BL/6 WT or D6−/− mice were cocultured with purified naïve T cells from allogeneic BALB/c mice, and subsequent T-cell proliferation was measured using a BrdU incorporation assay. Corroborating our phenotypic maturation observations, we found that DCs from D6−/− mice failed to induce proper alloreactive T-cell proliferation (p≤0.01), in contrast to DCs from WT mice (Figure 2A).
Figure 2.
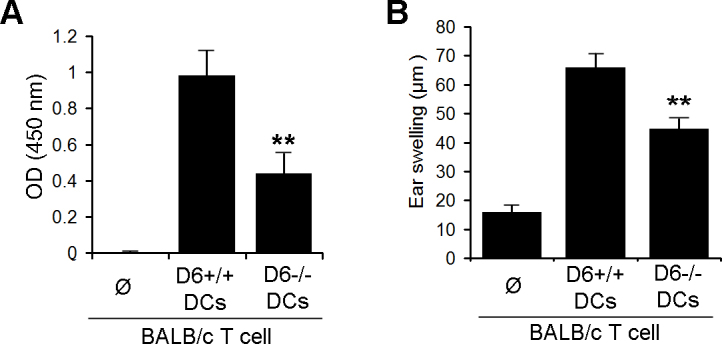
D6−/− dendritic cells generate impaired alloreactive T-cell responses. A: Mixed leukocyte reaction (MLR) assay of bone marrow (BM)-derived mitomycin C–treated dendritic cells (DCs) from wild-type (WT; D6+/+) or D6−/− mice. Cells were cocultured with allodisparate T cells (BALB/c), and less T-cell proliferation was evaluated using the bromodeoxyuridine (BrdU) assay. B: Delayed-type hypersensitivity (DTH) reaction in WT BALB/c mice receiving either splenic T cells from BALB/c mice (1×105) in 10 μl Hanks’ balanced salt solution alone or with 1×105 BM-derived DCs of either D6−/− or WT C57BL/6 mice. Cells were injected into the right pinna, and ear thickness was measured 48 h after injection. The experiment was performed twice, and representative data of one experiment are shown here; mean±standard error of the mean (SEM) of n=5 mice per group. P values are indicated comparing WT with D6−/− mice: **p≤0.01.
Next, we evaluated the role of D6 chemokine receptor expression by DCs on the alloreactive T-cell-mediated delayed-type hypersensitivity (DTH) reaction using the LAT assay [18]. Purified T cells from BALB/c mice primed against C57BL/6 were coinjected with WT or D6−/− DCs into the ear pinnae of the naïve BALB/c mice, and then the ear swelling was measured. Coinjection of T cells with DCs isolated from D6−/− mice elicited a significantly lower ear swelling compared to coinjection with WT DCs (p≤0.01) indicating that expression of D6 chemokine receptor is involved in the development of a DTH reaction (Figure 2B). These findings clearly suggested a functional role of D6 chemokine receptor expression in promoting DC phenotypic maturation and allostimulatory capacity.
D6 expression promotes T-cell alloreactivity in the corneal transplant model
Based on our findings that D6 expression promotes the phenotypic and functional maturation of DCs, we hypothesize that D6 plays a key role in T-cell alloreactivity and thus regulates the transplant outcome. We began our assessment by examining graft survival from C57BL/6 WT and D6−/− hosts transplanted with either fully allodisparate BALB/c (I-Ad) corneas or syngeneic grafts (I-Ab). Surprisingly, the fate of corneal grafts from BALB/c mice to WT or D6−/− recipients was not significantly different. Moreover, D6−/− mice rejected even syngeneic grafts (Figure 3A). Since opacity-grading scales to evaluate the rejected grafts in D6−/− mice demonstrated similar outcomes even in syngeneic mice, we next evaluated whether there are any differences in the generation of an alloreactive T-cell response in allotransplanted mice. Previously, we have reported a direct correlation between the frequency of CD4+/ interferon-γ+ (IFN-γ+) alloreactive T cells and immune rejection in the corneal transplant model [19,20]. Therefore, we harvested host lymph nodes from D6−/− or WT mice 3 weeks post-transplantation and enumerated alloreactive T-cell frequency via flow cytometry. Alloreactive IFN-γ-producing CD4 T cells were significantly lower in allografted D6−/− recipients compared to the WT hosts (p≤0.05; Figure 3B). In addition, we have previously demonstrated that host T-cell proliferation is directly correlated with corneal allograft rejection [19]. In this study, we observed significantly decreased proliferation of T cells from transplanted D6−/− recipients compared to WT hosts using the BrdU incorporation assay (Figure 3C).
Figure 3.
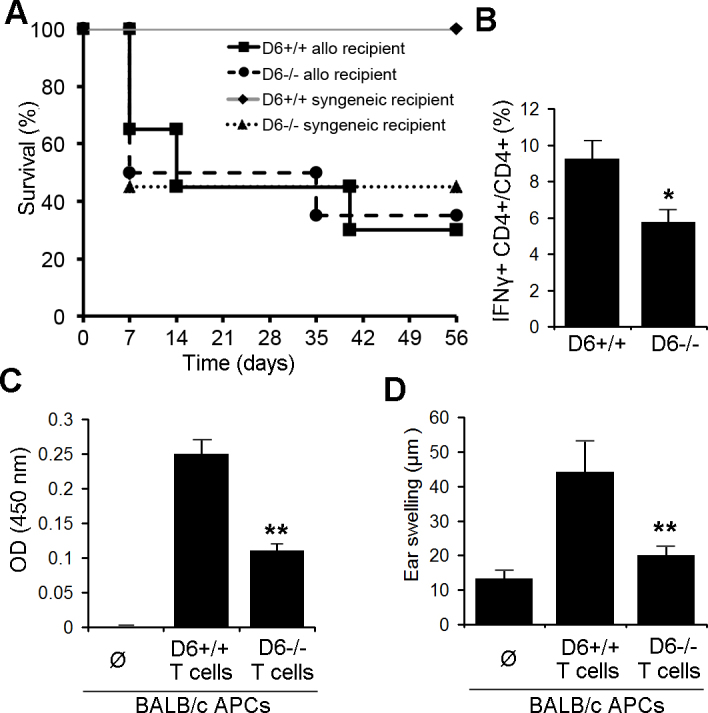
Transplantation in D6−/− hosts leads to a reduction in alloreactive T-cell frequencies. A: BALB/c or C57BL/6 wild-type (WT) corneas were engrafted onto C57BL/6 D6−/− or WT (D6+/+) mice. Kaplan–Meier survival curves are plotted here (n=10 per group). B: Flow cytometry analysis of CD4+ interferon-γ+ (IFN-γ+) T cells of lymph nodes from D6−/− and WT hosts 3 weeks post-transplantation is shown. *P≤0.05 (n=10 per group). C: Proliferation assay of purified T cells from D6−/− or WT mice; T cells were isolated 3 weeks post-transplantation of BALB/c corneas onto C57BL/6 D6−/− or WT mice, and cocultured with BALB/c antigen presenting cells (APCs). Subsequent T-cell proliferation was measured with the bromodeoxyuridine (BrdU) incorporation assay. D: Delayed-type hypersensitivity (DTH) reaction in naïve WT C57BL/6 mice receiving purified T cells from D6−/− or WT allografted mice and BALB/c APC is shown. Cells were injected into the right pinna, and ear thickness was measured 48 h after injection. The experiment was performed twice, and representative data of one experiment are shown here; mean±standard error of the mean (SEM) of n=5 mice per group. P values indicate comparison of WT and D6−/−: *p≤0.05, **p≤0.01.
Host T cells were also tested for DTH responses using the LAT assay. Immune rejection is mediated by DTH, and Miyazaki et al. previously demonstrated a positive correlation between DTH responses and corneal allograft rejection [21]. Purified T cells from D6−/− or WT mice 3 weeks post-transplantation were coinjected with BALB/c APCs into the ear pinnae of the naïve WT C57BL/6 mice. We observed that the T cells from the transplanted D6−/− hosts elicited a significantly lower DTH response compared to the T cells from the WT hosts (Figure 3D). These data provide evidence that D6 expression in corneal grafted hosts plays a critical role in triggering T-cell alloreactivity.
Increased infiltration of CD11c+ cells in D6−/− inflamed corneas
We investigated whether D6−/− mice display an increased innate inflammatory response and used a standardized model of cauterization-induced corneal inflammation. Seven days after cauterization, the corneas were stained for CD11c+ and enumerated using confocal microscopy. The inflamed corneas of the D6−/− mice showed a significantly increased number of CD11c+ cells compared to the inflamed corneas of the WT mice (p<0.05; Figure 4).
Figure 4.

A larger number of dendritic cells infiltrate the cornea of D6−/− mice following inflammation induced by cauterization. A: Confocal microscopy of the cornea from wild-type (WT; D6+/+) and D6−/− mice 7 days post-cornea cauterization. The cornea was stained for CD11c (green) cells (magnification 40X). B: The bar chart represents the quantification of the CD11c-infiltrated cells in the D6−/− and WT corneas 7 days post-cornea cauterization. *p≤0.05.
T cells from allografted D6−/− hosts have an impaired capacity to affect immune rejection following adoptive transfer
Though we did not detect a difference in corneal allograft rejection between the WT and D6−/− hosts, additional experiments that directly examined T-cell function (i.e., LAT and DTH assays) demonstrated impaired alloreactivity in D6−/− allografted hosts. Therefore, we adoptively transferred T cells from the allografted WT versus D6−/− hosts to the naïve wild-type C57BL/6 mice challenged with a fresh BALB/c corneal allograft. The allografted D6−/− T cells with reduced alloreactivity attenuated the immune rejection after adoptive transfer in C57BL/6 WT mice challenged with a fresh BALB/c cornea (p=0.015; Figure 5).
Figure 5.
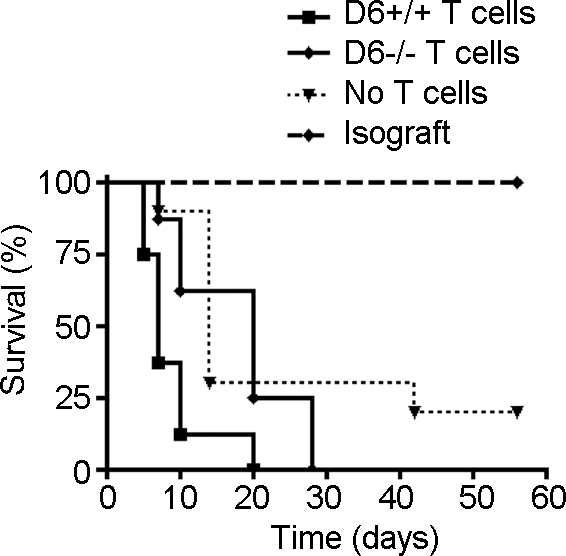
Immune rejection is attenuated after adoptive T-cell transfer from allografted D6−/−mice. Purified T cells from D6−/− or wild-type (WT; D6+/+) allografted mice were adoptively transferred into naïve wild-type C57BL/6 mice challenged with a fresh BALB/c cornea allograft. Graft survival is plotted in Kaplan–Meier curves (n=10 per group). P=0.015 D6-/- T cell vs. D6+/+ T cell group.
Discussion
This study for the first time demonstrates the role of D6 chemokine receptor expression by DCs in the development of alloimmune responses in transplanted hosts. We showed that D6 expression promotes DC maturation and is functionally relevant in enhancing alloimmune responses against the corneal graft demonstrated by adoptive transfer experiments.
The proresolving effect of D6 by chemokine scavenging has been well described. However, we demonstrated that D6 expression by DCs also has a proinflammatory effect by supporting T-cell-mediated alloimmunity. Studies supporting an immune amplifying function for D6 showed that D6−/− mice have an impaired ability to resolve a cutaneous inflammatory response after either topical application of the phorbol ester TPA or induction of granuloma development by subcutaneous injection of complete Freund's adjuvant (CFA) [3,5-7]. Nevertheless, little work has been done to dissect this role on the level of lymphatic endothelial cells or leukocytic expression of D6. One attempt was done in an animal model of colitis, where D6-deficient mice showed enhanced susceptibility to colitis [12]. However, restoration of D6 chemokine receptor expression in leukocytes and lymphatics through bone marrow transplantation demonstrated that the observed effect in this colitis mouse model is specifically related to the absence of D6 in the lymphatics, whereas leukocytes do not contribute to this phenotype.
The role of D6 in regulating the adaptive immune response was indirectly shown in alloimmune lung injury. Evaluation of D6−/− mice in the Th2 cell-dependent OVA model of lung inflammation showed reduced airway reactivity as well as increased numbers of DCs and T cells in the inflamed lung compared to WT mice. In addition, D6−/− mice demonstrated a rapid mortality subsequent to intranasal administration of low-dose Mycobacterium tuberculosis [5]. The conclusion of the study was that D6-deficient mice in this model displayed opposing effects on inflammation and airway reactivity, which resulted in a dysregulated, multiorgan, and systemic inflammatory response. Likewise, another study reported reduced airway hyperactivity to methalcholine in D6−/− mice using a Th2 cell-mediated model of experimental allergic asthma [13]. Because T cells are key mediators in allergic asthma such observations may be in part due to an impaired allergen-reactive T-cell response in D6−/− mice. In accordance to our data, Liu et al. reported reduced pathology in D6−/− mice to experimental autoimmune encephalomyelitis (EAE) due to impaired autoreactive T cell response providing a direct connection between D6 expression and T-cell response [14].
Previously, several studies have implicated DC maturation in mediating corneal alloimmunity [19,20,22]. Correspondingly, several lines of evidence showed that DC maturation, which we found to be positively regulated by D6 expression, plays a key role in promoting effector T-cell responses to the graft. The conclusion that D6 expression plays a role in phenotypic maturation of DCs is supported by our findings that BMDCs from D6−/− mice have consistently impaired expression of MHC II, CD80, CD86, and CD40 upon in vitro LPS stimulation. Further, DCs from D6−/− mice showed impaired capacity to stimulate T-cell proliferation and DTH responses suggesting that D6-mediated functional maturation of DCs is directly linked to the capacity to stimulate T-cell responses. Thus, it is likely that impaired DC maturation in engrafted D6−/− hosts contributes to impaired T-cell response reflected in lowered alloreactive frequencies, mixed leukocyte reaction and DTH responses, and decreased rejection.
Our findings are not in conflict with the current understanding that D6 is important in inflammatory resolution. Indeed, the opposing roles of D6 in inflammatory resolution and adaptive immune response activation may explain why corneal graft outcome in D6−/− and WT mice showed no significant difference in our study. In previous studies, cytokine expression analyses of corneal transplants have shown a biphasic pattern with no difference in the cytokine expression pattern between syngeneic and allogeneic recipients in the early phase (3–9 days postoperatively) [23]. Analyzing inflamed corneas after cauterization injury revealed increased infiltration of innate non-allospecific cells in the cornea of D6−/− mice as early as 1 week after injury. We hypothesize that these innate immune cells express high levels of inflammatory cytokines that might cause graft failure in allografted D6−/− mice. Yamada et al. have shown that corneal allografts were rejected by infiltration of innate immune cells independently of IFN-γ/IL-17 producing CD4 T cells, and the researchers proposed a still elusive pathway involved in graft rejection [24]. Likewise, we predict that a hyper-innate immune response in our transplanted D6−/− hosts may have caused the allograft to appear opaque in syngeneic hosts and thus be considered a failed transplant. We found that transplanted D6−/− hosts experience an approximate 50% immune rejection rate (Figure 3A) as early as 1 week post-transplantation—a process that typically requires 2 to 3 weeks to generate a prominent alloreactive T-cell response in corneal transplant [16,19,20].
In addition to the defective maturation of DCs, reduced migration of DCs from the grafted beds to lymph nodes is another potential factor that may have contributed to the blunted allosensitivity of T cells in D6−/− mice. It remains to be determined whether lymphatic vessel-expressed D6 contributes to T-cell alloreactive responses by facilitating DC migration to the lymph nodes. This is, in fact, conceivable since the neutralization of certain chemokines (e.g., CCR1–5) by D6 may consequently increase the reactivity to other chemokines that are not neutralized by D6, such as CCL19/21. CCL19/21 is important for the migration of DCs to the LN, where mature DCs elicit T-cell activation, and finally promote alloreactivity. Whether D6 promotes transplant survival not only by regulating DC maturation but also DC mobilization to the LNs requires further investigation.
Based on our data, we conclude that host D6 expression is important in the induction of alloreactive T-cell responses and immune rejection. In addition, we demonstrated that D6 expression by DCs plays a key role in their maturation and capacity to prime T cells. Further study is needed to clarify the connection between the proresolving effect of D6 in the lymphatic endothelium and its proinflammatory effect in DC maturation and on T-cell activation.
Acknowledgments
We thank Dr. Donald N. Cook from National Institute of Environmental Science for providing us with D6-deficient mice and Dr. Daniel Saban of Duke University for helpful scientific discussions. This work was supported by the NIH RO1EY12963 and the Eye Bank Association of America.
References
- 1.Nibbs RJ, Kriehuber E, Ponath PD, Parent D, Qin S, Campbell JD, Henderson A, Kerjaschki D, Maurer D, Graham GJ, Rot A. The beta-chemokine receptor D6 is expressed by lymphatic endothelium and a subset of vascular tumors. Am J Pathol. 2001;158:867–77. doi: 10.1016/s0002-9440(10)64035-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fra AM, Locati M, Otero K, Sironi M, Signorelli P, Massardi ML, Gobbi M, Vecchi A, Sozzani S, Mantovani A. Cutting edge: scavenging of inflammatory CC chemokines by the promiscuous putatively silent chemokine receptor D6. J Immunol. 2003;170:2279–82. doi: 10.4049/jimmunol.170.5.2279. [DOI] [PubMed] [Google Scholar]
- 3.Jamieson T, Cook DN, Nibbs RJ, Rot A, Nixon C, McLean P, Alcami A, Lira SA, Wiekowski M, Graham GJ. The chemokine receptor D6 limits the inflammatory response in vivo. Nat Immunol. 2005;6:403–11. doi: 10.1038/ni1182. [DOI] [PubMed] [Google Scholar]
- 4.Locati M, Torre YM, Galliera E, Bonecchi R, Bodduluri H, Vago G, Vecchi A, Mantovani A. Silent chemoattractant receptors: D6 as a decoy and scavenger receptor for inflammatory CC chemokines. Cytokine Growth Factor Rev. 2005;16:679–86. doi: 10.1016/j.cytogfr.2005.05.003. [DOI] [PubMed] [Google Scholar]
- 5.Di Liberto D, Locati M, Caccamo N, Vecchi A, Meraviglia S, Salerno A, Sireci G, Nebuloni M, Caceres N, Cardona PJ, Dieli F, Mantovani A. Role of the chemokine decoy receptor D6 in balancing inflammation, immune activation, and antimicrobial resistance in Mycobacterium tuberculosis infection. J Exp Med. 2008;205:2075–84. doi: 10.1084/jem.20070608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Martinez de la Torre Y, Locati M, Buracchi C, Dupor J, Cook DN, Bonecchi R, Nebuloni M, Rukavina D, Vago L, Vecchi A, Lira SA, Mantovani A. Increased inflammation in mice deficient for the chemokine decoy receptor D6. Eur J Immunol. 2005;35:1342–6. doi: 10.1002/eji.200526114. [DOI] [PubMed] [Google Scholar]
- 7.Collins CB, McNamee EN, Wermers JD, Lebsack MD, Rivera-Nieves J. Chemokine decoy receptor D6 in inflammatory bowel disease (IBD) and IBD-associated colon cancer. Gut. 2010;59:151–2. doi: 10.1136/gut.2009.192708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.McKimmie CS, Graham GJ. Leucocyte expression of the chemokine scavenger D6. Biochem Soc Trans. 2006;34:1002–4. doi: 10.1042/BST0341002. [DOI] [PubMed] [Google Scholar]
- 9.Bordon Y, Hansell CA, Sester DP, Clarke M, Mowat AM, Nibbs RJ. The atypical chemokine receptor D6 contributes to the development of experimental colitis. J Immunol. 2009;182:5032–40. doi: 10.4049/jimmunol.0802802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hansell CA, Schiering C, Kinstrie R, Ford L, Bordon Y, McInnes IB, Goodyear CS, Nibbs RJ. Universal expression and dual function of the atypical chemokine receptor D6 on innate-like B cells in mice. Blood. 2011;117:5413–24. doi: 10.1182/blood-2010-11-317115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bradford L, Marshall H, Robertson H, Kirby JA, Graham G, Ali S, O'Boyle G. Cardiac allograft rejection: examination of the expression and function of the decoy chemokine receptor D6. Transplantation. 2010;89:1411–6. doi: 10.1097/TP.0b013e3181da604b. [DOI] [PubMed] [Google Scholar]
- 12.Vetrano S, Borroni EM, Sarukhan A, Savino B, Bonecchi R, Correale C, Arena V, Fantini M, Roncalli M, Malesci A, Mantovani A, Locati M, Danese S. The lymphatic system controls intestinal inflammation and inflammation-associated Colon Cancer through the chemokine decoy receptor D6. Gut. 2010;59:197–206. doi: 10.1136/gut.2009.183772. [DOI] [PubMed] [Google Scholar]
- 13.Whitehead GS, Wang T, DeGraff LM, Card JW, Lira SA, Graham GJ, Cook DN. The chemokine receptor D6 has opposing effects on allergic inflammation and airway reactivity. Am J Respir Crit Care Med. 2007;175:243–9. doi: 10.1164/rccm.200606-839OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Liu L, Graham GJ, Damodaran A, Hu T, Lira SA, Sasse M, Canasto-Chibuque C, Cook DN, Ransohoff RM. Cutting edge: the silent chemokine receptor D6 is required for generating T cell responses that mediate experimental autoimmune encephalomyelitis. J Immunol. 2006;177:17–21. doi: 10.4049/jimmunol.177.1.17. [DOI] [PubMed] [Google Scholar]
- 15.Lutz MB, Kukutsch N, Ogilvie AL, Rossner S, Koch F, Romani N, Schuler G. An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J Immunol Methods. 1999;223:77–92. doi: 10.1016/s0022-1759(98)00204-x. [DOI] [PubMed] [Google Scholar]
- 16.Hamrah P, Yamagami S, Liu Y, Zhang Q, Vora SS, Lu B, Gerard CJ, Dana MR. Deletion of the chemokine receptor CCR1 prolongs corneal allograft survival. Invest Ophthalmol Vis Sci. 2007;48:1228–36. doi: 10.1167/iovs.05-1483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.McKimmie CS, Fraser AR, Hansell C, Gutierrez L, Philipsen S, Connell L, Rot A, Kurowska-Stolarska M, Carreno P, Pruenster M, Chu CC, Lombardi G, Halsey C, McInnes IB, Liew FY, Nibbs RJ, Graham GJ. Hemopoietic cell expression of the chemokine decoy receptor D6 is dynamic and regulated by GATA1. J Immunol. 2008;181:8171–81. doi: 10.4049/jimmunol.181.11.8170-a. [DOI] [PubMed] [Google Scholar]
- 18.Lin HH, Faunce DE, Stacey M, Terajewicz A, Nakamura T, Zhang-Hoover J, Kerley M, Mucenski ML, Gordon S, Stein-Streilein J. The macrophage F4/80 receptor is required for the induction of antigen-specific efferent regulatory T cells in peripheral tolerance. J Exp Med. 2005;201:1615–25. doi: 10.1084/jem.20042307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Saban DR, Chauhan SK, Zhang X, El Annan J, Jin Y, Dana R. 'Chimeric' grafts assembled from multiple allodisparate donors enjoy enhanced transplant survival. Am J Transplant. 2009;9:473–82. doi: 10.1111/j.1600-6143.2008.02535.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hattori T, Saban DR, Emami-Naeini P, Chauhan SK, Funaki T, Ueno H, Dana R. Donor-derived, tolerogenic dendritic cells suppress immune rejection in the indirect allosensitization-dominant setting of corneal transplantation. J Leukoc Biol. 2012;91:621–7. doi: 10.1189/jlb.1011500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Miyazaki D, Inoue Y, Yao YF, Okada AA, Shimomura Y, Hayashi K, Tano Y, Ohashi Y. T-cell-mediated immune responses in alloepithelial rejection after murine keratoepithelioplasty. Invest Ophthalmol Vis Sci. 1999;40:2590–7. [PubMed] [Google Scholar]
- 22.Huq S, Liu Y, Benichou G, Dana MR. Relevance of the direct pathway of sensitization in corneal transplantation is dictated by the graft bed microenvironment. J Immunol. 2004;173:4464–9. doi: 10.4049/jimmunol.173.7.4464. [DOI] [PubMed] [Google Scholar]
- 23.King WJ, Comer RM, Hudde T, Larkin DF, George AJ. Cytokine and chemokine expression kinetics after corneal transplantation. Transplantation. 2000;70:1225–33. doi: 10.1097/00007890-200010270-00017. [DOI] [PubMed] [Google Scholar]
- 24.Yamada J, Hamuro J, Fukushima A, Ohteki T, Tetai K, Iwakura Y, Yagita H, Kinoshita S. MHC-Matched Corneal allograft rejection in an IFN-γ/IL-17-independent manner in 57&BL/6 mice. Invest Ophthalmol Vis Sci. 2009;50:2139–46. doi: 10.1167/iovs.08-2993. [DOI] [PubMed] [Google Scholar]