

Abstract
The conflicts over sex allocation and male production in insect societies have long served as an important test bed for Hamilton's theory of inclusive fitness, but have for the most part been considered separately. Here, we develop new coevolutionary models to examine the interaction between these two conflicts and demonstrate that sex ratio and colony productivity costs of worker reproduction can lead to vastly different outcomes even in species that show no variation in their relatedness structure. Empirical data on worker-produced males in eight species of Melipona bees support the predictions from a model that takes into account the demographic details of colony growth and reproduction. Overall, these models contribute significantly to explaining behavioural variation that previous theories could not account for.
Keywords: social insects, worker reproduction, worker policing, sex allocation, inclusive fitness
1. Introduction
When discussing the evolution of social insects, Hamilton [1] emphasized various reproductive conflicts among colony members, mentioning that ‘the queen may be inclined to produce more males than the sterile workers regard as ideal. On the other hand a laying worker may want more males than the queen does—provided the extra males are her own (the worker's) offspring’ (p. 72). These insights soon led to mathematical theories of conflicts over sex allocation and male production [2,3]. Most models, however, have considered different kin conflicts separately, which sometimes leads to contradictory predictions. For example, in colonies headed by a single once-mated queen, the workers have been suggested to prefer both female-biased sex ratios and male production by highly related sister workers [2,3]. Worker reproduction also interacts with split sex ratios when the relatedness structure varies among colonies [4]. Hence, it is important to handle the simultaneous interaction of different types of kin conflicts [5–7].
Here, we will revisit the question outlined by Hamilton and develop coevolutionary models [8] to analyse the simultaneous selection for worker reproduction and policing (inhibition of worker reproduction by other workers [9]) and sex allocation biasing. Previously, one model investigated how effects on the sex ratio can alter selection for worker policing, albeit without explicit consideration of worker reproduction [6]. In our models, worker reproduction affects both the colony's sex ratio and its productivity (since worker-produced males can replace female workers). In the light of these models, we interpret empirical data from Melipona stingless bees, which show large variation in the amount of worker reproduction despite displaying minimal variation in kin structure [9].
2. Models
Our models are for male-haploid social insect colonies headed by a single queen. Under random mating, the genetic relatedness (r) among worker nest-mates ranges from 0.75 (monandry) to 0.25 (extreme polyandry). The genetic value of each individual is determined by the product of genetic relatedness, reproductive value (v) and mating success [2]. The mating success is given as an inverse of the number of a given sex that is produced in the population (1/F for females and 1/M for males). The models further assume weak selection (gradual evolution) and focus on the invasion of worker reproduction and worker policing in a situation where there can be simultaneous selection on the proportion of female eggs f laid by the queen and the amount of worker sex allocation biasing K.
We distinguish two main models (see the electronic supplementary material). In Model 1, the workers do not directly manipulate the sex ratio of the queen's offspring but instead replace some of the existing eggs with their own eggs. The basic model (Model 1a) reflects the life cycle of some ant species, such as slave-making ants, where little or no worker sex allocation biasing occurs [5], and where the sexual brood develops as a separate cohort. We then assume that worker-laid eggs replace random male- and gyne-destined eggs with probability w, without altering the total productivity of the colony. Worker reproduction in this situation can result in a suboptimal sex ratio but cannot come at a cost to worker production or colony productivity [7]. Model 1b is tailored to the life cycle of swarm-founding bees, and assumes that, in addition to sexual eggs, workers also randomly replace worker-destined queen-laid eggs. In this case, worker reproduction reduces colony productivity (male and daughter swarm production), with the cost depending on the rate with which workers provision new cells (b), worker mortality (μ) and the fraction of females that develop into queens (q). We solved Model 1b using both inclusive fitness and traditional population genetic approaches (see the electronic supplementary material), with identical results. Subsequently, the theoretical predictions were compared with data from eight species of Melipona bees [10] (see the electronic supplementary material). For both submodels, we determine the conditions under which worker reproduction can invade the population and determine the evolutionarily stable (ESS) level of worker reproduction when the proportion of female eggs laid by the queen f either can or cannot coevolve with the amount of worker reproduction. In addition, we determine the conditions under which worker policing is expected to invade, i.e. when collective worker interests disfavour worker reproduction [11].
Model 2 then takes into account active sex ratio manipulation by the workers and thus reflects the life cycle of many ants such as Formica. Queens lay gyne- and male-destined eggs in a separate cohort in proportions f and 1 – f. Workers can distinguish the sex of young larvae, and kill a fraction K of the brothers to transfer the resources with efficiency E to more highly related sisters. The worker-laid eggs face the same destiny as the other male eggs in the nest. As in Model 1a, we then examine the conditions under which worker reproduction and worker policing are able to invade the population.
3. Results
(a). Model 1: workers randomly replace queen-laid eggs
Invasion of worker reproduction in Model 1a is predicted when
| 3.1 |
which reduces to f > (4rsister − 1)/(4rsister + 1) given that f = F/(F + M), vf/vm = 2 (table 1). This shows that workers benefit from laying eggs when the sex ratio is female-biased and in polyandrous colonies also when the sex ratio is male-biased (figure 1a).
Table 1.
Boundary conditions for the invasion of worker reproduction and worker policing and the ESS proportion of worker-produced males and numerical investment in males (for Model 1) as well as the predicted proportions of removed brood owing to worker sex allocation biasing (for Model 2) (f, proportion of the queen's eggs that are female; r, sister–sister regression relatedness; rwm, regression relatedness of workers to worker-produced males = (1/n).rson + ((n−1)/n).rnephew, where n, number of egg-laying workers in a colony; b, number of new individuals produced per worker per day; q, proportion of females that develop as queens; μ, daily worker mortality rate; Ea, efficiency with which the resources freed by killing male brood is reinvested into the other sex. ESS values of traits are denoted by *.).
| workers randomly replace queen-laid eggs and do not manipulate the sex ratio |
workers reproduce and manipulate sex ratio towards own optimum |
||
|---|---|---|---|
| Model 1a: workers replace male- and gyne-destined eggs and colonies are founded by lone queens | Model 1b: workers replace male-, gyne- and worker-destined eggs and colonies are founded by swarms; worker reproduction reduces colony productivity | Model 2: workers replace male- and gyne-destined eggs and kill male brood to favour more highly related sisters | |
| example taxa in which assumptions apply | slave-making ants (where generally no worker sex ratio biasing occurs) | stingless bees (reproduction by swarming and males, workers and gynes all reared in approximately identical, mass provisioned cells) | ants |
| ESS fraction K of brood killed by workers (for given f) | n.a. | n.a. | 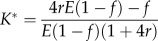 |
| invasion of worker reproduction (for given f and ESS value of K); figure 1 | 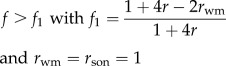 |
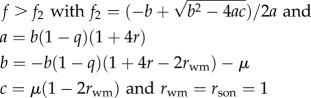 |
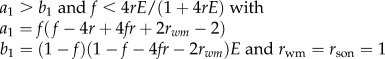 |
| invasion of worker policing (for given f and ESS value of K); figure 1 | 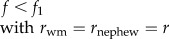 |
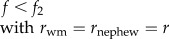 |
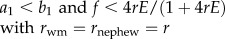 |
| ESS probability for workers to replace queen-laid eggs (for given f) | 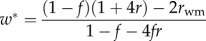 |
 |
See §2.4 in the electronic supplementary material Mathematica notebook |
| ESS proportion of worker-produced males ψ (for given f) |  |
 |
|
| numerical investment in males M (for given f) |  |
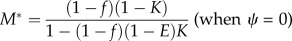 |
|
| joint ESS trait values (when the proportion of female eggs laid by the queen f can coevolve) | 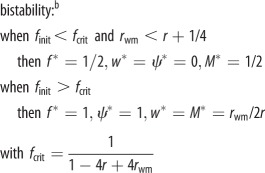 |
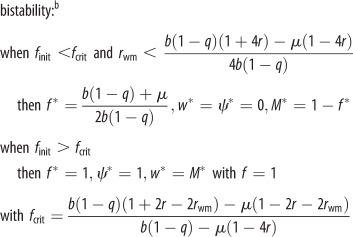 |
 |
| key predictions | very low and very high levels of worker reproduction with an approx. equal sex ratio are both alternative stable states | either very low or very high levels of worker reproduction are alternative stable states; laying of more female eggs by the queen should cause more worker reproduction | even under single mating, workers should frequently be sterile or police each others' reproduction and sex allocation biasing costs should cause colony sex ratios to be less female-biased than expected based purely on relatedness grounds |
| supporting evidence | zero worker reproduction in some slave-making ants, e.g. Epimyrma ravouxi and Harpagoxenus sublaevis, and very high worker reproduction in others, e.g. Polyergus rufescens (ψ = 1) and Protomognathus americanus (ψ = 0.4) [9]; approximately equal sex ratios in most slave-making ants [5] | species of Melipona stingless bees can either have very high (e.g. in M. favosa) or very low (e.g. in M. beecheii) levels of worker reproduction (see electronic supplementary material, table S4), and across eight species of Melipona, levels of worker reproduction are higher in species where queens lay relatively more female eggs (figure 2) | worker sterility and worker policing are common among many monogynous, monandrous ant species [9] and ant sex ratios tend to be less female-biased than expected based on relatedness grounds only, e.g. ca. 2 : 1 as opposed to 3 : 1 across 32 species of monogynous, monandrous ants [12] |
aInvasian conditions for arbitrary values of E are given in the electronic supplementary material Mathematica notebook in §§2.1.4 and 2.1.5.
bAlthough mathematically, a bistable outcome with either high or low worker reproduction is favoured depending on initial conditions, the equilibrium curves of f and ψ actually lie close together (see electronic supplementary material, figure S4), which means that the curves shown in figure 2 represent almost a joint neutral equilibrium from the perspective of both the queens and workers.
cThe lowest value of f that would result in female eggs not becoming limiting for the rearing of new queens.
Figure 1.

Areas of parameter space (defined by the proportion of the queen's eggs that are female f and sister–sister relatedness r) that allow invasion of worker reproduction and policing in a population where worker reproduction is initially absent (ψ ≈ 0) (cf. table 1). The results are shown for (a) Model 1a, where colonies are founded by lone queens and worker-laid eggs randomly compete with male- and gyne-destined queen-laid eggs; (b) Model 1b, where colonies reproduce by swarming and worker-laid eggs randomly compete with male-, gyne- and worker-destined queen-laid eggs (graph is shown for parameter values appropriate for Melipona stingless bees: b = 0.044, q = 0.0847, μ = 0.024; see electronic supplementary material, tables S2–S4); and (c–f) Model 2, where the workers manipulate the sex ratio towards their own optimum via the killing of less related brothers and reinvesting freed-up resources into more related sisters with efficiencies E = 0.3, 0.5, 0.8 and ≈1. The areas in the parameter space are green, no worker reproduction; blue, worker reproduction selected for, but potentially inhibited by worker policing; red, uninhibited worker reproduction. The optimal proportion of female eggs laid by the queen f is indicated with a dashed yellow line. In Models 1a and 1b, two alternative equilibria can be reached depending on initial conditions (initial f greater or smaller than the critical value indicated with the white dotted line, which is here drawn for n = 10; see electronic supplementary material, figure S4).
When worker reproduction is rare (i.e. w ≈ 0), the other workers should police their fellow workers and prevent them from laying eggs when
| 3.2 |
which reduces to f < (2rsister + 1)/(4rsister + 1) (table 1). This condition shows that under single mating (rsister = 3/4), worker policing is selected for as long as the queen lays less than 5/8 = 63% female eggs (figure 1a, blue area). Under multiple mating, conditions become even more relaxed (figure 1a).
When worker reproduction spreads, the sex-specific reproductive values change and their ratio becomes vf/vm = 2 − ψ, where the proportion of males that are workers' sons is ψ = w/[w + (1 − w)(1 − f)] [2]. The predicted level of worker reproduction favoured by collective worker interests can then be solved by assuming that the genetic value of a new nephew equals that of a random existing offspring (table 1). For monandrous colonies, the predicted proportion of males that will be workers' sons, assuming that many workers would reproduce within a colony (resulting in a relatedness to worker-produced males of rwm = rsister), is ψ = (8f − 5)/(5f − 2) (figure 2, solid blue curve and table 1), which will result in a numerical investment in males of M = (5f − 2)/(8f − 2). For example, if the queen lays 25% male eggs (1 − f = 0.25), workers are predicted to produce about a quarter of all eggs and 57% of all the males, thereby raising the fraction of males in the population to M = 44%. Furthermore, if the proportion of female eggs laid by the queen is allowed to coevolve, a bistable outcome is predicted in which either high or low worker reproduction with an approximately equal numerical investment sex ratio are both alternative stable outcomes (table 1 and electronic supplementary material, figure S4). These predictions are consistent with available data from several species of slave-making ants.
Figure 2.
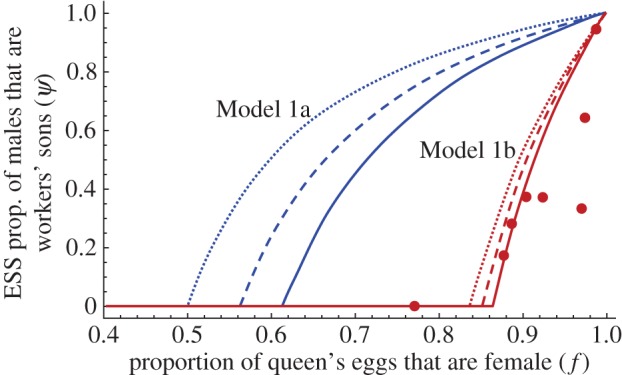
ESS proportions of the males that are workers' sons (ψ) predicted by collective worker interests in colonies headed by a single monandrous queen (r = 3/4) as a function of the proportion of the queen's eggs that are female (f) according to (i) Model 1a (blue curves), where colonies are founded by lone queens and worker-laid eggs randomly compete with male- and gyne-destined queen-laid eggs and (ii) Model 1b (red curves), where colonies reproduce by swarming and worker-laid eggs randomly compete with male-, gyne- as well as worker-destined queen-laid eggs, thereby impacting colony productivity. The curve for Model 1b is drawn for parameter values appropriate for Melipona stingless bees (b = 0.044, q = 0.0847, μ = 0.024, see electronic supplementary material, tables S2–S4), and the dots are observed values in eight species of Melipona (see the electronic supplementary material, table S4). Curves are drawn for the case where n = 1 (dotted lines), 2 (dashed lines) or 10 (full lines) workers would reproduce in a colony.
Invasion conditions for worker reproduction and worker policing under Model 1b for swarming colonies (table 1 and figure 1b) are derived analogously to equations (3.1) and (3.2). Owing to nonlinear effects of worker reproduction on colony productivity, however, the response of small changes in worker reproduction to the costs and benefits to the different classes of relatives are calculated using partial derivatives [13] (see the electronic supplementary material). We also calculate the ESS proportion of worker-derived males as a function of parameters b, f, q and μ, which determine the demographic growth of the colony (table 1). When worker mortality μ approaches 0 and when many workers would reproduce in the colony (rwm = rsister), both the invasion conditions and the predicted ESS are the same as in Model 1a (table 1). This is because worker reproduction then causes a linear tradeoff between male and female investment (swarm production). In addition, coevolution with the primary sex ratio produced by the queen predicts a bistable outcome in which both high and low levels of worker reproduction can be alternative stable states (table 1 and electronic supplementary material, figure S4).
Predictions based on demographic parameters estimated from Melipona bees are compared with the observed proportion of males that are workers' sons, ψ (figure 2 and electronic supplementary material, tables S1–S4). As predicted by our model, species can either have very high or very low levels of worker reproduction and the observed values of ψ increase with the proportion of the queen's eggs that are female (f) (figure 2, red curve; based on raw data: Spearman rank correlation R = 0.90, n = 8, p = 0.002; based on phylogenetic independent contrasts: Pearson R = 0.92, p = 0.001, electronic supplementary material, figure S1; for a sensitivity analysis see electronic supplementary material, figure S3). This occurs because the colony-level cost of worker reproduction is smaller when many of the queen's eggs are female and destined to become workers. Overall, the correlation between observed and predicted values of ψ (see electronic supplementary material, table S4) is also highly significant (Pearson R = 0.84, n = 8, p = 0.009).
(b). Model 2: workers readjust sex ratio to their own optimum
In Model 2, workers collectively remove some of the brood and manipulate sex allocation by moving resources from less valuable males to more highly related sisters with efficiency E. Typically, earlier recycling implies a higher efficiency. Selection on the level of killing (K), the primary sex ratio produced by the queen (f) and the amount of worker reproduction (w) is examined using a similar approach as in Model 1b [13].
When worker reproduction invades, the workers' offspring will be among the other male brood and treated with the same rules, as the workers do not distinguish between worker-derived and queen-derived brood. The conditions for worker reproduction and worker policing to invade a population where the queen initially lays most eggs and workers kill male brood with an optimal probability K* are presented in table 1 and figure 1c–f. When the sex ratio is close to the worker optimum, worker reproduction does not invade because of the attached recycling cost. If the sex ratio departs from the worker optimum, workers are selected to lay eggs. However, worker policing by other workers also invades the population, especially if the workers are not very closely related and the sex ratio is male-biased. If the primary sex ratio produced by the queen is allowed to coevolve with the amount of worker sex allocation biasing (see electronic supplementary material, figure S5), worker policing or even worker sterility are predicted for low values of E, and the traditional prediction of worker policing only being favoured under multiple mating (r < 0.5) is recovered only when E approaches 1. Nevertheless, an alternative equilibrium in which worker reproduction is common (ψ ≈ 1) and worker sex allocation biasing absent is also stable.
4. Discussion
Although a large meta-analysis of ca 100 species of ants, bees and wasps has supported the relatedness hypothesis and the role of polyandry in selecting for worker policing [9], significant unexplained variation in levels of worker reproduction remains [9,10]. Some of this variation has been attributed to productivity costs arising, for instance, from reproductive workers tending to engage less in colony maintenance tasks [10]. Our models, however, show that such direct efficiency costs are not required to explain the occurrence of worker policing or the absence of worker reproduction in monogynous and monandrous species. In fact, we show that this is well explained by either sex ratio costs or colony productivity costs caused by the replacement of worker-brood by worker-derived males. Hence, different species with identical relatedness structure can end up having widely different amounts of worker reproduction, depending on the specifics of sex allocation and costs incurred by the brood rearing dynamics of their colonies. In fact, in Model 2, the traditional result of worker policing being selected for when queens are multiply mated (rsister < 0.5) [9], was recovered, but only when workers can manipulate sex allocation at no cost (E → 1). When reallocation of resources between the sexes is costly, policing becomes favourable even under single mating [9], and if the sex ratio is close to the worker optimum these costs render worker reproduction unfeasible. In addition, in Model 1, worker policing can invade under single mating when worker-laid eggs randomly replace queen-laid eggs. Qualitatively similar conclusions on the effects of sex allocation biasing costs were earlier reached by using specific parameter values [6].
Convincing empirical support for our models is provided by data on Melipona bees, which show large variation in the amount of worker reproduction despite displaying hardly any variation in their kin structure and colonies in most species, being headed by a single once-mated queen [9]. In particular, data from eight species show that the observed level of worker reproduction increases with the female bias among the queen-laid eggs (figure 2). This is in agreement with our prediction that the colony-level cost of worker reproduction is relatively smaller when many of the queen's eggs are female and destined to become workers.
Overall, our study highlights a clear necessity of simultaneously considering several types of kin conflicts in order to arrive at more realistic predictions based on kin selection theory. We further show that, despite suggestions to the contrary [14,15], detailed assumptions on colony demography and colony growth can readily be added to inclusive fitness models, greatly improving the fit between model predictions and empirical observations. In the future, several other models in the social evolution literature would probably benefit from such detailed treatments.
Funding statement
This work was supported by the Centre of Excellence of Biological Interactions and by grants from the Academy of Finland to P.P. (122210) and H.H. (135970), by a grant of the Research Foundation–Flanders (FWO) and an Excellence Center financing grant to T.W. and FAPESP grant 2010/19717-4 to D.A.A.
References
- 1.Hamilton WD. 1972. Altruism and related phenomena, mainly in social insects. Annu. Rev. Ecol. Syst. 3, 193–232 (doi:10.1146/annurev.es.03.110172.001205) [Google Scholar]
- 2.Crozier RH, Pamilo P. 1996. Evolution of social insect colonies. Sex allocation and kin selection. Oxford, UK: Oxford University Press [Google Scholar]
- 3.Ratnieks FLW, Foster KR, Wenseleers T. 2006. Conflict resolution in insect societies. Annu. Rev. Entomol. 51, 581–608 (doi:10.1146/annurev.ento.51.110104.151003) [DOI] [PubMed] [Google Scholar]
- 4.Boomsma JJ, Grafen A. 1991. Colony level sex ratio selection in the eusocial Hymenoptera. J. Evol. Biol. 4, 383–407 (doi:10.1046/j.1420-9101.1991.4030383.x) [Google Scholar]
- 5.Bourke AFG. 2005. Genetics, relatedness and social behaviour in insect societies. In Insect evolutionary ecology (eds Fellowes MDE, Holloway GJ, Rolff J.), pp. 1–30 Wallingford, CT: CABI Publishing [Google Scholar]
- 6.Foster KR, Ratnieks FLW. 2001. The effect of sex-allocation biasing on the evolution of worker policing in hymenopteran societies. Am. Nat. 158, 615–624 (doi:10.1086/323588) [DOI] [PubMed] [Google Scholar]
- 7.Ohtsuki H, Tsuji K. 2009. Adaptive reproduction schedule as a cause of worker policing in social Hymenoptera: a dynamic game analysis. Am. Nat. 173, 747–758 (doi:10.1086/598488) [DOI] [PubMed] [Google Scholar]
- 8.Wenseleers T, Helanterä H, Hart A, Ratnieks FLW. 2004. Worker reproduction and policing in insect societies: an ESS analysis. J. Evol. Biol. 17, 1035–1047 (doi:10.1111/j.1420-9101.2004.00751.x) [DOI] [PubMed] [Google Scholar]
- 9.Wenseleers T, Ratnieks FLW. 2006. Comparative analysis of worker reproduction and policing in eusocial Hymenoptera supports relatedness theory. Am. Nat. 168, E163–E179 (doi:10.1086/508619) [DOI] [PubMed] [Google Scholar]
- 10.Hammond RL, Keller L. 2004. Conflict over male parentage in social insects. PLoS Biol. 2, 1–11 (doi:10.1371/journal.pbio.0020248) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ratnieks FLW. 1988. Reproductive harmony via mutual policing by workers in eusocial Hymenoptera. Am. Nat. 132, 217–236 (doi:10.1086/284846) [Google Scholar]
- 12.Boomsma JJ. 1989. Sex-investment ratios in ants: has female bias been systematically overestimated? Am. Nat. 133, 517–532 (doi:10.1086/284933) [Google Scholar]
- 13.Frank SA. 1998. The foundations of social evolution. Princeton, NJ: Princeton University Press [Google Scholar]
- 14.Nowak MA, Tarnita CE, Wilson EO. 2010. The evolution of eusociality. Nature 466, 1057–1062 (doi:10.1038/nature09205) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fromhage L, Kokko H. 2011. Monogamy and haplodiploidy act in synergy to promote the evolution of eusociality. Nat. Commun. 2, 397 (doi:10.1038/ncomms1410) [DOI] [PubMed] [Google Scholar]