

Abstract
Fluorescent dyes have been designed for internal cellular component specificity, and have been used extensively in the scientific community as a means to monitor cell growth, location, morphology, and viability. However, it is possible that the introduction of these dyes influences the basal function of the cell and, in turn, the results of these studies. Electrochemistry provides a non-invasive method for probing the unintended cellular affects of these dyes. The multianalyte microphysiometer (MAMP) is capable of simultaneous electrochemical measurement of extracellular metabolites in real-time. In this study, analytes central to cellular metabolism, glucose, lactate, oxygen, as well as extracellular acidification were monitored to determine the immediate metabolic effects of nuclear stains, including SYTO®, DAPI dilactate, Hoechst 33342, and FITC dyes upon the pheochromocytoma PC-12 cells and RAW 264.7 macrophages. The experimental results revealed that SYTO dye 13 significantly decreased glucose and oxygen consumption and increased extracellular acidification and lactate production in both cell lines, indicating a shift to anaerobic respiration. No other dyes caused significantly definitive changes in cellular metabolism upon exposure. This study shows that fluorescent dyes can have unintended effects on cellular metabolism and care should be taken when using these probes to investigate cellular function and morphology.
Keywords: Fluorescent dyes, cellular metabolism, bioenergetics, microphysiometry
INTRODUCTION
Cellular processes have been researched extensively by utilizing fluorescent dyes to aid in monitoring cellular changes; however, there has been little investigation to determine whether or not these dyes induce changes in cellular metabolism. Minute changes in basal cellular metabolism have the potential to manipulate overall cell health by controlling the amount of energy sequestered for aerobic versus anaerobic respiration, thereby influencing standard cellular functions. Potential fluctuations in basal cellular metabolism caused by fluorescent dye-based monitoring experiments could promote unintentional metabolic changes, giving rise to misleading data and faulty conclusions. To observe any minute changes in cellular metabolism caused by fluorescent dye exposure, we employed the multianalyte microphysiometer (MAMP).
The MAMP was developed by modifying sensor heads from the Cytosensor® Microphysiometer (Molecular Devices, Inc.) to include additional platinum electrodes for the simultaneous collection of metabolic analytes.1–4 Our lab has previously utilized the MAMP as a novel instrument to study metabolic changes caused by islet activation, toxin exposure, atherosclerotic development, and even to derive absolute basal cell rates for cancer cell lines.3, 5–9 In the present study, we are using this technology for the simultaneous, real-time measurement of the cellular consumption of glucose and oxygen (aerobic respiration) as well as their production of lactate (anaerobic respiration) and extracellular acidification rates.10
In these experiments, two cell types that are commonly used in research that utilize fluorescent dyes were studied: adrenal cells and macrophages. The various dyes used were SYTO dye 13, DAPI, dilactate, Hoechst 33342, and FITC. These commonly used fluorescent dyes allow researchers to track and monitor cell growth, location, morphology, and viability.11–14 In this study, the interaction of each dye with a cell line was investigated in real-time using the MAMP to observe any basal metabolic rate aberrations caused by fluorescent dye exposure. The overall goal of this study is to compare the resulting cellular metabolic profiles from exposure to each fluorescent dye.
MATERIALS AND METHODS
Chemicals
All materials were used as obtained from their commercial sources, unless otherwise noted. Lyophilized alamethicin from A.G. Scientific, Inc (San Diego, CA) was used to induce cell death at the end of each experiment. Seeds of PC-12 (ATCC CRL-1721) and RAW 264.7 (ATCC CRL-2278) cell lines were purchased through American Type Culture Collection (ATCC, Manassas, VA) and cultured in our lab using the appropriate ATCC protocols for each line. Experimental media supplements were obtained from the Media Core at Vanderbilt University (Nashville, TN) unless otherwise noted. Previously commercially available consumables were obtained through Molecular Devices Corporation (Sunnyvale, CA). For the hand-coated enzyme films, lactate oxidase (LOx) was purchased from Applied Enzyme Technology (Pontypool, UK). Other materials for the enzyme films, including bovine serum albumin (BSA, fraction V, 96%), glutaraldehyde (glutaric dialdehyde, 25 wt% solution in water) as a crosslinker to promote film stability, and glucose oxidase (GOx, Type IIS from Aspergillus niger) were purchased from Sigma. SYTO dye 13 and FITC were purchased through Life Technologies (Grand Island, NY). Additional materials are discussed in methodologies, as appropriate.
Plating Cells in Capsule Cups
Roughly 24 h prior to each experiment, the cell line to be used were placed in Corning cell culture inserts coated with Collagen I at a density of 5 × 105 cells/mL per capsule cup. The inserts were appropriately plated with one mL of this new cell suspension and given roughly 24 h to adhere to the cell capsule before subjecting the cell capsule to media flow in the microphysiometer.
MAMP Fluorescent Dye Dosage Experiments
For each experiment, running media was made using low-buffered (1 mM phosphate) RPMI-1640 with sterile water, which was adjusted to a pH of 7.40. Glucose was added to the running media at a concentration of 5 mM and was perfused through the MAMP chambers at 100 μL/min throughout the experiment at a temperature of 37°C. A spacer was inserted into the cell capsule, to create a chamber volume of 3 μL and height of 100 μm, while the insert membrane protected the cells from perturbations from the flow and from direct contact with the electrodes. The MAMP functions in continuous periods of 120 s cycles: 80 s flow followed by 40 s stop flow. The stop flow allows for the accumulation of lactate and other acidic by-products, and the consumption of glucose and oxygen within the chamber.1–4 The changes in extracellular acidification are measured by a light-addressable potentiometric sensor (LAPS).
Glucose and lactate are measured amperometrically via platinum electrodes coated with enzyme films, containing GOx and LOx, respectively. In each film, the analyte is oxidized by the enzyme, producing hydrogen peroxide. The H2O2 is then further oxidized at the electrode held at a potential of +0.6 V, resulting in a change in current. Oxygen is measured at a platinum electrode held at a potential of −0.45 V, which reduces O2 at the electrode to H2O2 and causes a change in current. The changes in current resulting from the consumption of glucose and oxygen and production of lactate are measured continuously throughout the experiment and plotted as current vs. time.
The running media was allowed to flow through for 60 min. Afterwards, a solution of 5 mM DMSO in running media was placed on the microphysiometer for 20 min as a control. Each dye was allowed to run through for 30 min, and running media was subsequently replaced for an additional 60 min to allow the cells to recover. SYTO dye 13 was used at the suggested concentration of 0.5 μL/mL, and FITC was used at the suggested concentration of 10 μM.11–14 Afterwards, the cells were lysed with 75 μL alamethicin to ensure proper calibrations occur for the data conversion into metabolic rates. Successive calibrations were performed, including 0 mM to 5 mM glucose and 0 mM to 0.2 mM lactate. After calibrations, the dye was reintroduced into the chambers to ensure no interaction with the enzymatic sensors.
Data Analysis
For each analyte measured, the maximum change in current was calculated by determining the difference in the peak height during stop flow periods and baseline values during flow periods, yielding Δip. Each Δip value for glucose and lactate is then calibrated against the corresponding baseline shifts acquired during the calibrations performed at the end of each experiment, after cell death. The linear range of glucose oxidase is often exceeded in these conditions, thus glucose calibrations are calculated using Michaelis-Menten kinetics with double-reciprocal plots. Oxygen data were treated in the same manner, however instead of performing in-line calibrations for the oxygen sensor, the baseline signal is assumed to be the concentration of dissolved oxygen, 0.24 mM and calibrated accordingly.15, 16 These data series can then be converted to molar consumption rates by using the size of the chamber, flow rate, and number of cells, providing values as mole per cell per second (mol/cell*s). Data are then normalized against the 10 min baseline prior to dye exposure to provide a dynamic trace of each experiment. Extracellular acidification is calculated by the Cytosoft® program based on the slope of stop flow peaks and each time point is subsequently normalized and included in the dynamic trace.
The percent change in peak heights were calculated from the maximum change in peak height during exposure compared to the baseline basal values prior to exposure. To determine statistical significance, a paired or unpaired t-test was used as appropriate. Any p-value of less than 0.05 was considered statistically significant.
RESULTS AND DISCUSSION
The SYTO fluorescent dye is a cell-permeant nucleic acid stain and can be used to stain RNA and DNA in both live and dead cells.11–14 Its various uses in the field of research include flow cytometric enumeration of live and fixed cells, increased efficiency in fluorescence signal/noise ratio, detection of apoptosis in live cells, and fluorescent tracking of neuronal orientation and function.11–14 The recommended dosages of SYTO 13 for fluorescent live cell experiments is 0.5 μL dye/mL media and was used in these experiments. Due to its widespread use in a variety of research fields, it was imperative to ensure cellular metabolic viability during its use.
After analyzing and reviewing the data from these MAMP experiments, it was evident that a significant change had occurred in the metabolic rates of glucose and oxygen consumption, lactate production, as well as extracellular acidification of both the PC-12 and RAW 264.7 cells, upon the addition of the SYTO 13 fluorescent dye. During SYTO exposure, there was a marked increase in extracellular acidification and lactate production rates concurrent with a decrease in glucose and oxygen consumption for both PC-12 and RAW 264.7 cells (Figures 1 and 2). During the 60 min recovery period, after dye exposure, the metabolic rates never reached their previous basal values; thereby revealing the long-lasting effects the dye had produced.
Figure 1.
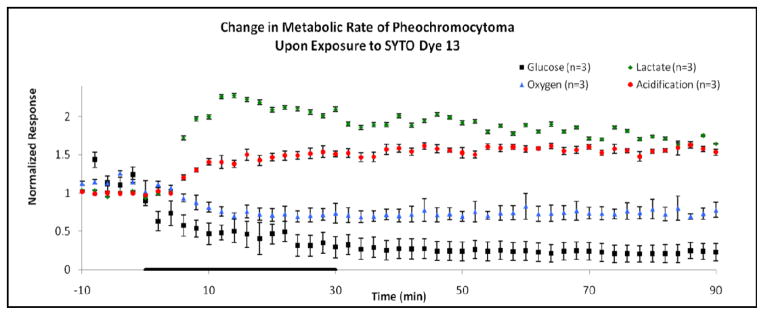
Normalized metabolic responses for 0.5 μL dye/mL SYTO dye 13 exposure to pheochromocytoma (PC-12) cells. Basal baseline measurements were obtained 10 min prior to exposure, with standard error variation in the baseline follows: glucose ± 27.1% (−8 to 0 min time points used for baseline), lactate ± 1.90%, oxygen ± 3.80%, and acidification ± 0.34%. The SYTO dye was administered for thirty minutes during the experiment, as indicated by the black line on the horizontal axis of the graph. Afterwards, running media was applied for a 60 min recovery period. Significant increases in lactate production and extracellular acidification suggest the increasing utilization of the anaerobic respiratory pathways for ATP production. Decreases in glucose and oxygen consumption highlight the impairment of the aerobic respiratory pathway by SYTO dye exposure.
Figure 2.
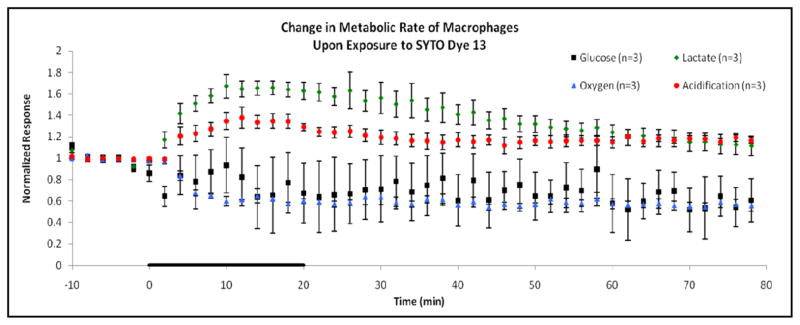
Normalized metabolic responses for 0.5 μL dye/mL SYTO dye 13 exposure to macrophage (RAW 264.7) cells. Basal baseline measurements were obtained 10 min prior to exposure, with standard error variation in the baseline follows: glucose ± 3.70%, lactate ± 1.50%, oxygen ± 0.34%, and acidification ± 0.11%. The cells were exposed to SYTO dye for 20 min, as indicated by the black line on the horizontal axis of the graph. Afterwards, running media was applied for a 60 min recovery period. Again, significant increases in anaerobic respiration and decreases in aerobic respiration suggest a metabolic shift in ATP production brought on by the cellular exposure to SYTO dye.
For PC-12 cells, there was a 93 ± 14 % decrease in glucose consumption, coupled with a 47 ± 7 % decrease in oxygen consumption. These data can be seen in Figure 1 and Table 1. The onset of this decrease occurred within the first 6 min of exposure time and continued throughout the exposure. Aerobic respiration never regained its basal metabolic rates during recovery, indicating that the SYTO dye prohibited proper functioning of the aerobic respiration pathway. On the other hand, lactate production increased (126 ± 3 %) during exposure, as did extracellular acidification (54 ± 8 %). These rapid changes also occurred near the onset of dye exposure and had a prolonged effect well after recovery, suggesting a metabolic shift to anaerobic respiration due to the presence of SYTO dye.
Table 1.
Percent Changes in Metabolic Rates Due to Fluorescent Dye Exposure. PC-12 and RAW cell lines are compared with percent changes during SYTO dye 13, and FITC exposure.
| Cell Line | Dye | Glucose (%) | Lactate (%) | Oxygen (%) | Acidification (%) |
|---|---|---|---|---|---|
| PC-12 | FITC (10 μM) | −22 ± 36 | 3 ± 10 | 1 ± 3 | 2 ± 5 |
| SYTO Dye 13 (0.5 μL dye/mL) | −93 ± 14 | 126 ± 3* | −47 ± 7* | 54 ± 8* | |
| RAW 264.7 | SYTO Dye 13 (0.5 μL dye/mL) | −36 ± 29 | 67 ± 11* | −43 ± 1* | 38 ± 10* |
Asterisks (*) denote a p value of less than 0.05, for a comparison between peak height percent changes and basal measurements.
These responses were mimicked in the macrophage cell line as well, as can be seen in Figure 2 and Table 1. Glucose and oxygen consumption decreased by 36 ± 29 % and 43 ± 1 %, respectively, again suggesting a shift away from typical aerobic respiration rates for ATP production. Simultaneously, there was an increase in lactate production by 67 ± 11 % and an extracellular acidification increase of 38 ± 10 %. In both cell lines, there is a rapid shift from anaerobic respiration toward anaerobic respiration due to the introduction of SYTO dye to the cells.
SYTO dye was confirmed to cause significant changes in cellular metabolism of both cell lines. It was determined that the DMSO treatment alone did not cause any statistically significant change in the basal metabolic rate of the cells (data not shown). The dye was also run after the cells were killed with alamethicin to determine if there was any sensor interference from the dye, but there was no change in signal from this application, thereby ruling out this possibility. Overall, there were no factors indicating that these marked metabolic rate changes could have been caused by any parameters other than the dye exposure. The SYTO dye 13 results carry considerable impact on any research that has utilized this specific, or a similar functioning, fluorescent dye.
Alternate dyes were tested as well, including Hoechst 33342, a cell-permeable DNA stain that binds preferentially to adenine-thymine regions of the minor groove of DNA. Hoechst 33342 is used to specifically stain nuclei of live or fixed cells and tissues. Hoechst 33342 has been used extensively in scientific research including studies concerning in vivo stem cell research and transport kinetics.17 In addition to Hoechst 33342, alternate dyes studied were DAPI, dilactate and FITC. These dyes were commercially available and sold in a research kit for fluorescent dyes, however these three dyes did not produce any statistically significant responses during cellular exposure. The lack of significant response from these dyes suggests that they do not alter the metabolic rates of either PC-12 or RAW 264.7 cell lines.
Based on the research presented here, utilization of fluorescent dyes for the monitoring of cellular functioning can inhibit cellular bioenergetics. In the dyes studied, SYTO dye 13 exhibited significant metabolic disturbances when exposed to both PC-12 and RAW 264.7 cell lines. These data suggests that the mechanism of action of these dyes interferes in some way with glucose conversion to ATP for cellular energy, thus potentially altering any cellular monitoring experiments. Future studies should focus on investigating the mechanism of action of these dyes and how their compositions contribute to metabolic pathway alteration, and if there is a safe concentration to use these dyes that will adequately avoid metabolic perturbation.
Acknowledgments
We would like to thank Dr. Jennifer McKenzie, Dr. Rachel Snider, Dr. Jason Greene, the Beckman’s Scholars Program, and the Vanderbilt Institute for Integrative Biosystems Research and Engineering. Funding: Systems Biology and Bioengineering Undergraduate Research Experience: NIH U01 AI 061223, Beckman’s Scholars Foundation, and HDTRA 1-09-01-0013.
Funding Sources
Funding for this project came from: NIH U01 AI 061223, Beckman’s Scholars Foundation, and HDTRA 1-09-01-0013.
Footnotes
Author Contributions
The manuscript was written through contributions of all authors. All authors have given approval to the final version of the manuscript.
References
- 1.Eklund SE, Cliffel DE, Kozlov E, Prokop A, Wikswo J, Baudenbacher F. Anal Chim Acta. 2003;496:93–101. [Google Scholar]
- 2.Eklund SE, Kozlov E, Taylor DE, Baudenbacher F, Cliffel DE. Methods in Molecular Biology (Totowa, NJ, United States) 2005;303:209–223. doi: 10.1385/1-59259-901-X:209. [DOI] [PubMed] [Google Scholar]
- 3.Eklund SE, Snider RM, Wikswo J, Baudenbacher F, Prokop A, Cliffel DE. J Electroanal Chem. 2006;587:333–339. [Google Scholar]
- 4.Eklund SE, Taylor D, Kozlov E, Prokop A, Cliffel DE. Anal Chem. 2004;76:519–527. doi: 10.1021/ac034641z. [DOI] [PubMed] [Google Scholar]
- 5.Eklund SE, Thompson RG, Snider RM, Carney CK, Wright DW, Wikswo J, Cliffel DE. Sensors. 2009;9:2117–2133. doi: 10.3390/s90302117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ciobanu M, Taylor DE, Jr, Wilburn JP, Cliffel DE. Anal Chem. 2008;80:2717–2727. doi: 10.1021/ac7021184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Snider RM, McKenzie JR, Kraft L, Kozlov E, Wikswo JP, Cliffel DE. Toxins. 2010;2:632–648. doi: 10.3390/toxins2040632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Snider RM, Ciobanu M, Rue AE, Cliffel DE. Anal Chim Acta. 2008;609:44–52. doi: 10.1016/j.aca.2007.12.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zeiger SL, McKenzie JR, Stankowski JN, Martin JA, Cliffel DE, McLaughlin B. Biochim Biophys Acta. 2010 doi: 10.1016/j.bbadis.2010.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Eklund SE, Taylor D, Kozlov E, Prokop A, Cliffel DE. Anal Chem. 2004;76:519–527. doi: 10.1021/ac034641z. [DOI] [PubMed] [Google Scholar]
- 11.Ullal AJ, Pisetsky DS, Reich CF. Cytometry A. 2010;77:294–301. doi: 10.1002/cyto.a.20833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Guindulain T, Comas J, Vives-Rego J. Appl Environ Microbiol. 1997;63:4608–4611. doi: 10.1128/aem.63.11.4608-4611.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Guindulain T, Vives-Rego J. Letters in Applied Microbiology. 2002;34:182–188. doi: 10.1046/j.1472-765x.2002.01069.x. [DOI] [PubMed] [Google Scholar]
- 14.Wlodkowic D, Skommer J, Darzynkiewicz Z. Cytometry A. 2008;73A doi: 10.1002/cyto.a.20535. [DOI] [PubMed] [Google Scholar]
- 15.Owicki JC, Bousse LJ, Hafeman DG, Kirk GL, Olson JD, Wada HG, Parce JW. Annu Rev Biophys Biomol Struct. 1994;23 doi: 10.1146/annurev.bb.23.060194.000511. [DOI] [PubMed] [Google Scholar]
- 16.Casciari JJ, Sotirchos SV, Sutherland RM. J Cellular Physiology. 1992;151:386–394. doi: 10.1002/jcp.1041510220. [DOI] [PubMed] [Google Scholar]
- 17.Darzynkiewicz Z, Huang X. Current Protocols in Immunology. 2004;60:5.7.1–5.7.18. doi: 10.1002/0471142735.im0507s60. [DOI] [PubMed] [Google Scholar]