

Abstract
Bovine tuberculosis is a bacterial disease caused by Mycobacterium bovis in livestock and wildlife with hosts that include Eurasian badgers (Meles meles), brushtail possum (Trichosurus vulpecula), and white-tailed deer (Odocoileus virginianus). Risk-assessment efforts in Michigan have been initiated on farms to minimize interactions of cattle with wildlife hosts but research on M. bovis on cattle farms has not investigated the spatial context of disease epidemiology. To incorporate spatially explicit data, initial likelihood of infection probabilities for cattle farms tested for M. bovis, prevalence of M. bovis in white-tailed deer, deer density, and environmental variables for each farm were modeled in a Bayesian hierarchical framework. We used geo-referenced locations of 762 cattle farms that have been tested for M. bovis, white-tailed deer prevalence, and several environmental variables that may lead to long-term survival and viability of M. bovis on farms and surrounding habitats (i.e., soil type, habitat type). Bayesian hierarchical analyses identified deer prevalence and proportion of sandy soil within our sampling grid as the most supported model. Analysis of cattle farms tested for M. bovis identified that for every 1% increase in sandy soil resulted in an increase in odds of infection by 4%. Our analysis revealed that the influence of prevalence of M. bovis in white-tailed deer was still a concern even after considerable efforts to prevent cattle interactions with white-tailed deer through on-farm mitigation and reduction in the deer population. Cattle farms test positive for M. bovis annually in our study area suggesting that the potential for an environmental source either on farms or in the surrounding landscape may contributing to new or re-infections with M. bovis. Our research provides an initial assessment of potential environmental factors that could be incorporated into additional modeling efforts as more knowledge of deer herd factors and cattle farm prevalence is documented.
Introduction
Bovine tuberculosis (bTB) is a bacterial disease (Mycobacterium bovis) in livestock and wildlife that results in United States Department of Agriculture-mandated depopulation of cattle herds costing farmers millions in lost revenue throughout the world [1], [2]. Preliminary efforts by the Michigan Department of Agriculture-Animal Industry Division (MDA) have created protocols that farmers could follow to reduce potential for M. bovis infection of cattle in Michigan’s Modified Accredited Zone (MAZ) [3]. Basic risk-assessment efforts were needed, however, to address the spatial context of disease epidemiology (i.e., infection probability if a farm is adjacent to a bTB-infected farm) and dynamics of primary reservoirs in the MAZ (i.e., white-tailed deer [Odocoileus virginianus]).
The influence of wildlife activity on transmission of M. bovis depends on possible hosts and their ability to transmit disease [4]–[6]. Direct observation of farms in Michigan, USA documented that indirect interactions between cattle and white-tailed deer were dominated by use of pastures and silage storage areas but deer fed from hay racks or silage troughs on only one occasion [7]. Visitation of farm yards and cattle-use areas by sixteen GPS-collared white-tailed deer was documented in Michigan’s MAZ and deer were documented using confined feeding areas, water tubs, and pastures [8]. Prevalence of M. bovis in deer was as high as 10–12% in some townships but currently can range from 2 to ≥5% in some townships due to changes in management regulation for deer and feeding on some cattle farms [3], [9], [10].
Reoccurrence of M. bovis in farms depopulated of cattle in Michigan would suggest an environmental or mammalian host source of re-infection as several farms have become re-infected with M. bovis on ≥2 separate occasions often spanning 3–7 years between re-infection [3], [11]. Under natural shaded conditions on pastures, survival of M. bovis in cattle feces was documented to span up to 5 months post-application during winter but only up to 2 months during spring and summer [12]. Effluent plots tested positive for M. bovis for up to 29, 13, and 35 days post application for soil, radishes, and lettuce, respectively, in a study in raised garden plots (lined plywood boxes) [13]. Although environmental and anthropogenic variables that influence odds of contracting a disease have been addressed in North America [3], [14], only recently has the spatial matrices incorporating proximity to adjacent infected individuals been successfully modeled in disease epidemiology research with advances in software (i.e., WinBUGS; [15]–[17]). Understanding the spatial dynamics of M. bovis will increase our ability to predict future spread or occurrences and variables influencing these occurrences across the MAZ in the northern, lower peninsula of Michigan.
To incorporate spatially explicit data, likelihood of infection probabilities within a geographically designed grid can be determined for cattle herds that tested positive for M. bovis and incorporated into a Bayesian hierarchical framework. Although on-farm management practices are believed to influence M. bovis transmission, consensus on the most important farm-level factor responsible for transmission is absent and varied across studies in Europe and North America ([11], [18], [19] but see [3] for a detailed summary). Spatially explicit data on environments that cattle farms occupy is often lacking for researchers attempting to understand the underlying distribution of disease in the landscape and has not been modeled in this system since discovery of M. bovis in a free-ranging white-tailed deer in 1975. Our objectives were to model odds of infection with M. bovis in cattle farms at the herd level using Bayesian hierarchical analysis by incorporating prevalence of M. bovis in the deer population, environmental variables, spatial structure, and unstructured spatial heterogeneity across the MAZ in Michigan. An understanding of conditions that sustain survival of M. bovis in the environment would be valuable to our ability to focus surveillance for the disease and predict future spread or occurrences outside of the MAZ in Michigan.
Materials and Methods
Study area
We conducted our study in the northern, lower peninsula of Michigan in the MAZ. The 8,062 km2 study area included the entirety of Alcona, Alpena, Montmorency, Oscoda, and Presque Isle counties (Fig. 1). The area encompassed the majority of the cattle farms where M. bovis has been found in Michigan. Our study area surrounds Deer Management Unit 452 that has been defined as the bovine tuberculosis core area by the Michigan Department of Natural Resources (MDNR) due to the high prevalence of M. bovis in free-ranging deer and the presence of M. bovis-positive cattle on farms (Fig. 1; [20], [21]). Vegetation categories present in our study area included: developed that included roads, development, and barren land; grass that included pasture/hay fields and native grasses; agriculture that included crops other than forage; forest that included upland hardwood stands (Quercus alba, Acer rubrum, and A. saccharum), aspen stands (Populus tremuloides and P. grandidentata), hardwood/aspen mixed stands, upland conifer stands (Pinus glauca, P. banksiana, and P. resinosa), and hardwood/conifer mixed stands, and swamp that included lowland conifer forests/swamps (P. glauca, P. mariana, Thuja occidentalis, Abies balsamea. and Latrix laricinea). Elevations in the area ranged from 150–390 m above sea level and the mean annual temperature was 6.6°C, the mean rainfall was 72.5 cm, and there was a mean snowfall of 175 cm [22].
Figure 1. Sampling grid (25 km2 cells) that contained all cattle farms tested for bovine tuberculosis within the Modified Accredited Zone (5 counties in bold) in the upper, lower peninsula of Michigan.

Deer Management Unit 452 (dashed polygon) is considered the bovine tuberculosis core area for surveillance in white-tailed deer (Outset). Cattle farms that tested negative (yellow circles) and positive (black crosses) for Mycobacterium bovis used in Bayesian Hierarchical models overlayed on percent sand within a portion of the study area in the upper, lower peninsula of Michigan (Inset).
To link the disease status (positive or negative) of each farm in the sample to deer herd and environmental-level predictors, we first overlaid a 5×5 km square grid having a resolution of 25 square kilometers (hereafter referred to as grid cell), which is equal to a quarter township in size. We selected quarter townships as the proper resolution given that township would likely be too coarse a scale and section would be too fine a resolution for model convergence based on previous research with Bayesian hierarchical models [17]. There were a total of 368 grid cells covering the MAZ and we assigned each farm in our study to its appropriate grid cell.
Observation component
Cattle farm data
Our data included 762 cattle farms of known infection status (observation component) provided by the Michigan Department of Agriculture and Rural Development (MDA) from mandatory testing of cattle on an annual basis. Based on current knowledge that indirect transmission (i.e., environmental source) of M. bovis to cattle may be important, we used replicates for each farm that tested positive on ≥1 occasion over the 11 year span of our study (58 positive, 704 negative) with positive farms coded as 1 and negative farms coded as 0. We included a farm each time it tested positive for M. bovis and this was deemed warranted because conditions of that farm or host characteristics in the area were responsible for continued infections of M. bovis and replicates would weigh environmental characteristics of farms that tested positive on >1 occasion. Because farms that tested positive for M. bovis were depopulated of cattle then re-populated prior to subsequently testing positive, each farm was considered an independent observation for the purposes of our study design. Each farm was tested annually for M. bovis but we did not include additional negatives as replicates because that would have likely masked the effects of the positives that we were attempting to model for odds of infection.
We included all cattle farms in this region because we wanted to determine the environmental drivers of disease that were not associated with farm practices and remained unaltered when cattle farms were depopulated or permanently closed (e.g., surrounding habitats, soil composition). Furthermore, we did not include any farm-level covariates (e.g., herd size, feeding practices) in our models because farm mitigation strategies were initiated by the MDA during our study [3], would be difficult to quantify and standardize across the study region, and would only be considered a contamination source (e.g., cattle fed in deer habitat) but would not influence environmental persistence or survival of M. bovis in the landscape.
Process component
Host-level variables
The MDNR provided section-level data on deer prevalence for M. bovis from 1995 to 2009. We limited our analysis to white-tailed deer prevalence for 2005 to 2009 because deer herd management, ban on baiting deer, and mitigation of on-farm practices indicated that deer prevalence has stabilized within the past 5 years thus, more reflective of current deer prevalence [10], [23]. Apparent prevalence of M. bovis in deer was determined for each grid cell by dividing the total number of deer testing positive by the total number of deer tested resulting in percent prevalence that was entered into models. Annual deer densities were provided by the MDNR from 2005 to 2009 at the county level for the MAZ based on Sex-Age-Kill reconstruction technique or additional methods if available [24], [25]. We averaged deer density over the time period to match deer prevalence (2005–2009) that resulted in a single estimate of deer density per grid cell. Due to the logistical constraints of estimating deer densities at a fine scale, such as to the section-level, we used the only available data to represent deer densities in our study site as deer per square kilometer. We did not select mean deer prevalence or mean deer density for the entire span of sampling of cattle farms because we were interested in modeling the effects of more recent deer prevalence that likely would be influenced by initiation of deer and on-farm management practices in 1996.
Environmental-level variables
We hypothesized a priori that M. bovis patterns on farms were structured in part by spatial heterogeneities in features of the landscape, therefore, we identified four environmental-level predictors of infection based on optimal survival characteristics of M. bovis identified in the literature [3]. Proportion of sand content in the soil was selected because dry sandy loam soils at the proper pH and moisture promoted bacteria growth [26], [27]. Proportion of the landscape ponding frequently and proportion of swamp/wetland were selected because the duration of standing water occurring in non-wetlands (ponding) and soil characteristics of inundated areas (wetlands) were conducive to long-term survival of M. bovis [27], [28]. Mean soil pH was selected because soil pH from 5.8 to 6.9 was conducive to culture of M. bovis at the optimum temperature (37°C) for survival [27], [29]. Sand (where “sand” was defined as soil particles with size >2 μm), landscape ponding frequently, and soil pH was determined using the Advanced Mode of the Soil Data Viewer available through the National Resources Conservation Service of the US Department of Agriculture in ArcMap 9.x (ArcMap; Environmental Systems Research Institute, Redlands, CA, USA). Soil Data Viewer provides interactive mapping software to query the Soil Survey Geographic (SSURGO) database and descriptive characteristics for each soil type. Sand was expressed as the percent sand within a soil type polygon (∼2 ha resolution) for each soil map unit [30]. Each grid cell was thus potentially composed of multiple soil type polygons with varying sand contents. Therefore, we calculated the mean percent sand for each 25 km2 grid cell using a weighted average based on the area of the various soil type polygons and their associated sand content. Similarly, in the Advanced Mode of the Soil Data Viewer, we identified the proportion of each grid cell that ponded frequently with frequently defined as “ponding occurs, on the average, more than once in 2 years and the chance of ponding is >50% in any year [30].” Similar to sand and ponding frequently, we calculated the mean soil pH for each 25 km2 grid cell using a weighted average based on the area of the various pH polygons and their associated pH value.
We used the National Land Cover Database of 2006 (NLCD) that was created from Landsat 7 imagery to determine the proportion of swamp/wetland across the study site (MRLC 2007). To standardize analyses across the MAZ, we reclassified land cover from the NLCD into 8 categories used in Kaneene et al. [11]: hardwood forest, coniferous forest, mixed forest, open areas and shrubs, wetland/swamp, agricultural use, open water, and other (industrial, residential). We extracted the proportion of wetland/swamp from NLCD within each grid cell with all environmental-level variables, except soil pH, presented as a percentage in a grid cell in modeling efforts. Skewness of data and correlation among covariates was assessed but data transformations and exclusions were not considered necessary prior to entering into models.
Statistical analysis
We used a Bayesian hierarchical model structure [31], [32] with logistic regression models (described below) to examine how recent TB prevalence at the deer herd-level and landscape factors influenced the probability of a farm being infected, while adjusting for the other covariates and spatial structure in the data [33]. To adjust for latent spatial effects we included two types of random effects that captured both the influence of the local neighborhood (i.e., cells sharing a border or vertex with each 25-km2 grid cell; CAR) in determining spatial clustering of M. bovis, as well as any spatially independent influences occurring at the 25-km2 spatial resolution of our grid (HET). Our models were constructed hierarchically to accommodate the fact that information from multiple levels (i.e., fixed-effects and spatial random effects) was being used to estimate individual-level infection probabilities. Taking a Bayesian approach, we used Markov Chain Monte Carlo (MCMC) simulation methods available within the program WinBUGS [34] to produce the unnormalized joint posterior density for the parameters of interest across all models examined based on the product of the data likelihood and the prior densities for each parameter. We used this approach to estimate the posterior marginal probability distributions for the parameters governing the influence of the host- and environmental-level, and spatial random effects predictors on the probability of infection. For each model we ran three independent Markov chains with varying initial values for 350,000 iterations and discarded the first 100,000. We thinned the Markov chains by keeping every twentieth iteration for inference. To determine if the three Markov chains used for each model had converged on the same posterior distribution, we used the statistical program R with the package boa [35] and employed several graphical and quantitative diagnostics, including autocorrelation plots, trace plots, and univariate corrected scale reduction factors for each parameter. To assess simultaneous convergence of all parameters for the top models, we calculated the multivariate potential scale reduction factor [32], [36], [37]. All inferences were based on the mean of each parameter’s marginal posterior distribution.
Likelihood functions
The observation component of the data likelihood specifies each farm’s observed M. bovis infection status as a Bernoulli random variable with parameter 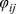 :
:
where Yij is the infection status of the i
th farm for i = 1, …, n from the j
th grid cell for j = 1, …, m, and 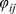 represents the probability of infection. Thus, given the probability of infection we assume each farm’s infection status is conditionally independent.
represents the probability of infection. Thus, given the probability of infection we assume each farm’s infection status is conditionally independent.
The process component of the data likelihood models, via the logit link function, defined as the probability of infection as a function of individual, environmental and spatial covariates as well as random effects that account for spatial variability:
| (1) |
where 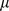 defines the baseline M. bovis infection probability,
defines the baseline M. bovis infection probability, 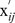 is the transpose of a
is the transpose of a 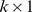 matrix of covariates for the i
th farm from the j
th grid cell,
matrix of covariates for the i
th farm from the j
th grid cell, 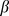 is a
is a 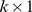 vector of parameter estimates for these covariates,
vector of parameter estimates for these covariates, 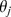 is a random effect for the j
th grid cell capturing extra-binomial variability over the entire study region at the individual quarter township scale (HET; i.e., 25 km2), and
is a random effect for the j
th grid cell capturing extra-binomial variability over the entire study region at the individual quarter township scale (HET; i.e., 25 km2), and 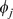 is a random effect term for the j
th grid cell that models the extra-binomial variability associated with local disease clustering (CAR; i.e., grid cells closer together will have similar infection probabilities due to proximity of cattle farms and pastures).
is a random effect term for the j
th grid cell that models the extra-binomial variability associated with local disease clustering (CAR; i.e., grid cells closer together will have similar infection probabilities due to proximity of cattle farms and pastures).
Prior distributions
We assumed non-informative N (0, 100,000) prior distributions for each of the 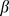 parameters, and an improper (flat) prior over the entire real line for µ. For the random effect describing region-wide heterogeneity (HET), we assumed the following:
parameters, and an improper (flat) prior over the entire real line for µ. For the random effect describing region-wide heterogeneity (HET), we assumed the following:
| (2) |
To describe the spatial structure, we assumed an intrinsic Gaussian conditional autoregressive prior with a sum to zero constraint [31] for the local clustering random effect (CAR):
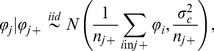 |
(3) |
where 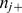 is the number of grid cells that share a border or vertex with the j
th grid cell. Thus, the random effect of the j
th grid cell is conditional on the values of its
is the number of grid cells that share a border or vertex with the j
th grid cell. Thus, the random effect of the j
th grid cell is conditional on the values of its 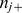 (usually = 8) neighboring cells. Adjacency matrices were created with the Adjacency for WinBUGS Tool in ArcMap that provides a matrix relating one areal unit to a collection of neighboring areal units in text files for use in WinBUGS.
(usually = 8) neighboring cells. Adjacency matrices were created with the Adjacency for WinBUGS Tool in ArcMap that provides a matrix relating one areal unit to a collection of neighboring areal units in text files for use in WinBUGS.
Because of the marginal specification for 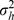 and conditional specification for
and conditional specification for 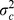 of the random effects, we generated prior distributions for the precisions (i.e.,
of the random effects, we generated prior distributions for the precisions (i.e., 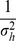 and
and 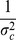 ) using simulations in WinBUGS where we varied the values of the parameters and determined the parameter values that created an expectation of ∼0.5 for the psi metric [38], where psi is as follows:
) using simulations in WinBUGS where we varied the values of the parameters and determined the parameter values that created an expectation of ∼0.5 for the psi metric [38], where psi is as follows:
| (4) |
.
These simulations attempted to ensure an equal emphasis on the priors of the standard deviations of the random effects [31]. Based on our simulation results we defined
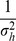 ∼ Gamma(10.368,3.22) and
∼ Gamma(10.368,3.22) and 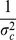 ∼ Gamma(1.0,1.0).
∼ Gamma(1.0,1.0).
Model Selection
To test our original hypothesis that environmental variables would influence the odds of M. bovis infection, our set of candidate models for logistic regression consisted of 12 different structures with strictly additive effects on the logit scale (Table 1). The 12 logistic regression models represented all possible combinations of deer and environmental variables, as well as inherent regional and local spatial structure of the data. Viewing these 12 models as competing hypotheses, we used deviance information criterion (DIC) [15], [33], [39] to compare the models’ respective fits to the data from sampled farms, and then estimated parameters and examined goodness-of-fit and other metrics for the top models. For model comparison we used DIC weights [33], which allow for an intuitive comparison of the evidence in the data for each candidate model. The weights are considered a measure of the strength of evidence in the data for i th model being the “best” model of those within the candidate set, and therefore provide a measure of model selection uncertainty [33], [39].
Table 1. Model selection results for the candidate set of models investigating the effect of covariates on the probability of bovine tuberculosis infection from 2005–2010 in Modified Accredited Zone in Michigan, USA using non-informative N (0, 0.00001) prior distributions for the fixed effects parameters and diffuse gamma priors for the random effects with farm-level factors removed.
| Model Terms | Dbar | Dhat | pD | DIC | ΔDIC | Weights |
| Deer + ------ + HET + Envir | 272.9 | 226.4 | 46.5 | 319.4 | 0.0 | 0.6386 |
| Deer + ------ + HET + ------ | 276.2 | 231.5 | 44.7 | 320.9 | 1.5 | 0.3073 |
| ------ + ------ + HET + Envir | 275.9 | 227.5 | 48.4 | 324.4 | 4.9 | 0.0540 |
| Deer + CAR + ------ + ------ | 289.0 | 235.5 | 53.5 | 342.4 | 23.0 | 0.0000 |
| ------ + CAR + ------ + Envir | 290.5 | 231.7 | 58.8 | 349.3 | 29.8 | 0.0000 |
| Deer + CAR + ------ + Envir | 298.1 | 232.0 | 66.1 | 364.2 | 44.7 | 0.0000 |
| Deer + CAR + HET + ------ | 302.3 | 224.8 | 77.5 | 379.8 | 60.4 | 0.0000 |
| ------ + CAR + HET + Envir | 309.3 | 226.2 | 83.1 | 392.3 | 72.9 | 0.0000 |
| Deer + ------ + ------ + Envir | 387.9 | 380.9 | 7.0 | 394.9 | 75.5 | 0.0000 |
| Deer + ------ + ------ + ------ | 394.2 | 391.2 | 2.9 | 397.1 | 77.6 | 0.0000 |
| ------ + ------ + ------ + Envir | 405.5 | 400.7 | 4.8 | 410.3 | 90.8 | 0.0000 |
| Deer + CAR + HET + Envir | 317.1 | 222.1 | 95.0 | 412.0 | 92.6 | 0.0000 |
“Deer” represents deer herd factors: apparent prevalence of deer and deer density. “Envir” represents the environmental variables: percent sand, percent ponding frequently, percent swamp/wetland, and mean soil pH in each sampled farms quarter township grid cell. “HET” represents the random effect capturing region-wide heterogeneity and “CAR” is the random effect capturing local clustering.
We used parameter estimates from the top model to calculate odds ratios for the effect of variables on M. bovis infection odds among farms in that area. Model averaging was not appropriate because DIC, unlike BIC and AIC, is not based on any assumption of a “true” model and is primarily concerned with short-term predictive ability [33]. We treated host-level (deer prevalence, deer density) and environmental-level predictors (percent sand, soil pH, proportion of wetland/swamp, and area that ponded frequently) as a group of variables, such that they were all entered or removed from the models together (Table 1). To examine the goodness-of-fit of the top model from our candidate set we conducted a numerical posterior predictive check [32]. We examined correlation and trace plots, as well as the estimates of the corrected scale reduction factor for each parameter and multivariate potential scale reduction factors and determined that that the three chains for each model had converged (data not shown). For each dataset, the top models selected via our model selection procedures and their corresponding estimates were similar regardless of prior specification (data not shown; [33]).
Results
Out of the 762 cattle farms tested on an annual basis, 704 were negative while 37, 9, and 1 tested positive for M. bovis on 1, 2, and 3 occasions, respectively. Of our 12 models determined a priori, the top two models combined to account for over 95% of the summed weights of all models considered. Models weights of 95% provided strong assurance that some combination of these 2 models and their parameters reflected the underlying infection-generating process far better than other models in the candidate set (Table 1). Parameters in the top model included deer herd factors and local landscape features suggesting that these factors increased the odds of M. bovis infection to cattle in the northern, lower peninsula of Michigan (Table 2). As documented since initial diagnosis of a positive white-tailed deer in 1975, deer apparent prevalence ranged from 0.0% to 5.2% and was the most supported variable in the top model (odd ratio = 1.004, 95% CI = 1.001 to 1.007; Table 2). Sand within the vicinity of sampled farms was by far the most supported environmental variable and ranged from 37% to 79% on farms that test positive whereas sand ranged from 17% to 88% for cattle farms that tested negative for M. bovis (Fig. 1). The odds of infection for M. bovis increased by about 4% for every 1% increase in sand in the area (odd ratio = 1.036, 95% CI = 1.01 to 1.07; Table 2).
Table 2. Mean parameter estimates, standard deviation (SD), Monte Carlo error (MC error), odds ratios (OR), and 95% credible intervals for best-fitting model investigating the effect of covariates on the probability of bovine tuberculosis infection from 2005–2010 in Modified Accredited Zone in Michigan, USA.
| Parameter | Mean | SD | MC error | 2.50% | Median | 97.5% | OR | 95% CI |
| Intercept | −3.401 | 2.886 | 0.02 | −9.219 | −3.317 | 2.031 | 0.0333 | 0.00 to 7.622 |
| Deer density | −0.2219 | 0.1454 | 0.00 | −0.515 | 0.219 | 0.056 | 0.8001 | 0.60 to 1.06 |
| Deer prevalence | 0.4147 | 0.1412 | 0.00 | 0.137 | 0.414 | 0.697 | 1.004 | 1.001 to 1.007 |
| Percent wetland | −0.0209 | 0.0262 | 0.00 | −0.074 | −0.020 | 0.029 | 0.9793 | 0.93 to 1.03 |
| Percent sand | 0.0357 | 0.0152 | 0.00 | 0.007 | 0.035 | 0.067 | 1.0363 | 1.01 to 1.07 |
| Soil pH | 0.04212 | 0.3368 | 0.00 | −0.591 | 0.035 | 0.732 | 1.0430 | 0.55 to 2.08 |
| Percent ponding | 0.0240 | 0.035 | 0.00 | −0.043 | 0.023 | 0.095 | 1.0243 | 0.96 to 1.10 |
| HET | 1.213 | 0.4819 | 0.00 | 0.524 | 1.128 | 2.39 | 3.3636 | 1.69 to 10.91 |
“HET” represents the random effect capturing region-wide.
Our analyses also identified that an unstructured random effect (HET) dominated over spatial structure (CAR) in influencing the odds of M.bovis infection (odd ratio = 3.36, 95% CI = 1.69 to 10.91; Table 2). Inclusion of the unstructured random effect in both our top models would suggest that additional covariates are driving odds of M. bovis infection and not spatial occurrence of M. bovis-positive farms in our study area.
Discussion
Our findings support the premise that deer herd-related factors play an important role in sustaining M. bovis presence in the northern, lower peninsula of Michigan similar to that found in previous research [11], [40]. Because mitigation measures have been implemented to reduce deer access to feeding and cattle use areas [3], we focused our analysis simply on deer density and deer prevalence in the area without reference to farm practices. Farm practices have been the primary focus of most efforts to control M. bovis transmission between reservoirs and hosts [11], [18], [41], [42] but are difficult to standardize and document for inclusion in modeling efforts. Bayesian hierarchical models provide the ability to assess spatially the influence of region-wide cattle farm and host-level variables while adjusting for additional covariates [31], [43]. The bias that may have been introduced by entering 36% of cattle farms more than once into our models was deemed warranted to achieve our objectives of assessing environmental factors that may lead to continued presence of M. bovis on cattle farms. Furthermore, we don’t deny that farm-management practices are important in M. bovis infection on cattle farms, however, it is very difficult to accurately represent or measure one of these farm practices in a standardized format for hundreds of farms to include in our models. For this reason, we selected objectives that would look at determining new facet in understanding M. bovis infection (i.e., environmental variables) as opposed to conducting another study that suggested a different farm practice was responsible for bTB infection in cattle farms as documented in previous research [11], [18], [41], [42].
Although deer densities in the area have been reduced to about 10–15 deer/km2 since 1994 and deer prevalence has remained at just below 2% overall at the DMU 452-level [10], the role of a primary host for M. bovis in this region is still influencing transmission of M. bovis based on our study. Including mean deer prevalence for the duration of the study (i.e., 1998–2009) in our models would likely have yielded similar results to our current modeling effort but the role of the host would still be supported nonetheless. Due to public pressure about low deer densities and continued occurrence of cattle positive for M. bovis on an annual basis, reducing deer densities further is likely not possible. Relatedly, even with our conservative estimate of prevalence of M. bovis in deer (i.e., 2005–2009), deer prevalence was in the most supported model even though the odds ratio would likely have been greater if we included 1998–2009.
Our study identified environmental variables that were not possible to assess in previous modeling efforts that can further assist agencies in their attempts to eradicate M. bovis in Michigan. Environmental variables have been documented to contribute to presence or viability of infectious agents of disease in several areas in North American and Europe [17], [44], [45] although environmental sampling has yet to identify M. bovis on cattle farms in this region [46], [47]. Variables to consider should be based on a priori knowledge of the disease agent studied and mechanisms that may hinder or promote survival. Survival of M. bovis has been linked to moist, humid environments that maintain the proper soil type and pH [48]–[50]. Our top model indicated that a combination of landscape variables played an important role in determining infection probability for M. bovis on farms which was the impetus for us to select a combination of covariates that were conducive to moist, humid environments with low sunlight exposure on the landscape (e.g., wetlands, ponding frequency, soil types). Percent sand was a significant predictor and increased the odds of M. bovis infection by 3.6% for every 1% increase in proportion of sandy soil in the local area (Table 2). Survival of M. bovis has been linked to soil type, temperature and pH [3], [29], [48] but research in natural settings has limited the advancement of knowledge in this area.
The exact composition of sandy soils or functional role of these soils that make them conducive to the survival of M. bovis likely requires further research. Sandy soils are defined as having loose particle sizes (> 2 μm) with structural integrity during desiccation that may promote survival of M. bovis when associated with wetlands and areas that routinely have standing water. Moraines that have steep slopes and sandy, well-drained soils dominated by northern hardwood forests were linked to infection of white-tailed with M. bovis [51] but are confounded by the fact that they characterize preferred deer habitat in years with heavy oak mast production. Based on prevalence studies on white-tailed deer and our current study of farms positive for M. bovis, environmental or landscape-specific characteristics would appear to be a logical focus of future studies and potential assessment of viable M. bovis detection in soils or water. Areas that contain these sandy, well-drained soils in northern hardwood forests likely provide moist, humid microclimates conducive to survival of M. bovis for extended periods of time and should be considered for focused surveillance for M. bovis in deer and cattle as well as the focus of future on-farm mitigation measures.
Since initial detection of M. bovis in farms over a decade ago, farms have tested positive on an annual basis. Even after considerable efforts have been implemented to reduce deer densities, limiting or preventing aggregation of deer (e.g., ban on baiting of deer), and limiting deer-cattle interactions through on-farm mitigation measures, Michigan still does not have M. bovis-free status. Although modeling efforts are unable to include movements of cattle between farms and its influence on movements and spread of M. bovis, repeated positive tests have occurred on numerous farms since the first farm tested positive in 1998 [28]. Repeated positive tests would suggest that potentially an environmental source conducive to survival of the bacteria may be responsible for maintaining M. bovis in the region. Mycobacteria have waxy, lipid-rich cell walls that are relatively resistant to biocides used in decontamination procedures thus complicating management of the disease.
Unlike previous work in this region, we were able to assess spatial processes that may be influencing the transmission or presence of M. bovis using a Bayesian hierarchical modeling framework [31], [43]. We identified that unstructured spatial heterogeneity (HET) was included in the top two models explaining infection of M. bovis on farms. If probability of infection was driven by spatial structure or clustering of disease (i.e., contiguous grid cells more alike than 2 arbitrary grid cells) at our site, we would have expected spatial structure (CAR) to be included in our top models but the opposite occurred [43]. Unstructured spatial heterogeneity would suggest that additional covariates that we may not have accounted for in our models were also influencing the disease across our study region. As stated previously, movement of cattle between farms and additional on-farm practices is controlled or mitigated to some extent for some farms but is very difficult to enforce and has been documented to be the cause of contamination of several cattle farms within and outside of the MAZ [21]. Our unstructured spatial heterogeneity could simply be on-farm practices not included in our modeling effort or additional environmental covariates conducive to survival or destruction of M. bovis such as slope/aspects conducive to direct exposure to ultraviolet light.
Although host prevalence in DMU 452 and clustering of cattle farms were considered important in M. bovis infection in previous research [11], [52], our results suggest that less spatially structured components in the landscape are influencing continued occurrence of M. bovis in cattle on farms in Michigan. Considering the logistics of managing the host and reservoir through the northern, lower peninsula of Michigan, management efforts could focus on environmental and landscape characteristics that are potentially supporting the continued presence of M. bovis in Michigan. Landscape characteristics that are conducive to survival of M. bovis such as habitat inundated with standing water, soil composition, and prime deer habitat should be the focus of future management and research. On-farm mitigations should focus efforts in areas at high risk for continued survival of M. bovis prior to mandating complete risk mitigation of farms for an entire area such as DMU 452 (1,479 km2) or the 5 county study area (8,062 km2). Focused surveillance and management of on-farm practices in reservoirs for disease in domestic livestock would provide a logistically feasible approach to combating a disease in an endemic area as well as new areas that the disease has recently been introduced or spread from the endemic area.
Acknowledgments
We thank the Michigan Department of Natural Resources for deer-related data used in this manuscript. We thank Daniel J. O’Brien, Michigan Department of Natural Resources, for a helpful review of an earlier draft of this manuscript. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Funding Statement
Funding for this research was provided by the National Wildlife Research Center of the United States Department of Agriculture, Animal and Plant Health Inspection Service, Wildlife Services. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. de Lisle GW, Bengis RG, Schmitt SM, O'Brien DJ (2002) Tuberculosis in free-ranging wildlife: detection, diagnosis and management. Review Science Technology, OIE 21: 317–334. [DOI] [PubMed] [Google Scholar]
- 2. Kaneene J, Thoen CO (2004) Tuberculosis. Journal of the American Veterinary Medical Association 224: 685–691. [DOI] [PubMed] [Google Scholar]
- 3. Walter WD, Anderson CW, Smith R, Vanderklok M, Averill JJ (2012) On-farm mitigation of transmission of tuberculosis from white-tailed deer to cattle: literature review and recommendations. Veterinary Medicine International 2012: 1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Clifton-Hadley RS, Wilesmith JW, Stuart FA (1993) Mycobacterium bovis in the European badger ( Meles meles ): epidemiological findings in tuberculosis badgers from a naturally infected population. Epidemiology and Infection 111: 9–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Griffin JM, Hahesy T, Lynch K, Salman MD, McCarthy J, et al. (1993) The association of cattle husbandry practices, environmental factors and farmer characteristics with the occurrence of chronic bovine tuberculosis in dairy herds in the Republic of Ireland. Preventive Veterinary Medicine 17: 145–160. [Google Scholar]
- 6. Corner LAL (2006) The role of wild animal populations in the epidemiology of tuberculosis in domestic animals: How to assess the risk. Veterinary Microbiology 112: 303–312. [DOI] [PubMed] [Google Scholar]
- 7.Hill JA (2005) Wildlife-cattle interactions in northern Michigan: implications for the transmission of bovine tuberculosis Thesis. Logan: Utah State University. 1–58 p. [Google Scholar]
- 8.Berentsen AR, Miller RS, Misiewicz R, Malmberg JL, Dunbar MR (2013) Characteristics of white-tailed deer visits to cattle farms: implications for disease transmission at the wildlife-livestock interface. European Journal of Wildlife Research: In press.
- 9. O'Brien DJ, Schmitt SM, Fierke JS, Hogle SA, Winterstein SR, et al. (2002) Epidemiology of Mycobacterium bovis in free-ranging white-tailed deer, Michigan, USA, 1995-2000. Preventive Veterinary Medicine 54: 47–63. [DOI] [PubMed] [Google Scholar]
- 10. O'Brien DJ, Schmitt SM, Fitzgerald SD, Berry DE (2011) Management of bovine tuberculosis in Michigan wildlife: Current status and near term prospects. Veterinary Microbiology 151: 179–187. [DOI] [PubMed] [Google Scholar]
- 11. Kaneene JB, Bruning-Fann CS, Granger LM, Miller R, Porter-Spalding BA (2002) Environmental and farm management factors associated with tuberculosis on cattle farms in northeastern Michigan. Journal of the American Veterinary Medical Association 221: 837–842. [DOI] [PubMed] [Google Scholar]
- 12. Williams RS, Hoy WA (1930) The viability of B. tuberculosis (bovinus) on pasture land, in stored faeces and in liquid manure. Journal of Hygeine 30: 413–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Van Donsel DJ, Larkin EP (1977) Persistence of Mycobacterium bovis BCG in soil and on vegetables spray-irrigated with sewage effluent and sludge. Journal of Food Protection 40: 160–163. [DOI] [PubMed] [Google Scholar]
- 14. Carstensen M, DonCarlos MW (2011) Preventing the establishment of a wildlife disease reservoir: a case study of bovine tuberculosis in wild deer in Minnesota, USA. Veterinary Medicine International 2011: 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Farnsworth ML, Hoeting JA, Hobbs NT, Miller MW (2006) Linking chronic wasting disease to mule deer movement scales: a hierarchical bayesian approach. Ecological Applications 16: 1026–1036. [DOI] [PubMed] [Google Scholar]
- 16. Osnas EE, Heisey DM, Rolley RE, Samuel MD (2009) Spatial and temporal patterns of chronic wasting disease: fine-scale mapping of a wildlife epidemic in Wisconsin. Ecological Applications 19: 1311–1322. [DOI] [PubMed] [Google Scholar]
- 17. Walter WD, Walsh DP, Farnsworth ML, Winkelman DL, Miller MW (2011) Soil clay content underlies prion infection odds. Nature Communications 2: 1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Hutchings MR, Harris S (1997) Effects of farm management practices on cattle grazing behaviour and the potential for transmission of bovine tuberculosis from badgers to cattle. The Veterinary Journal 153: 149–162. [DOI] [PubMed] [Google Scholar]
- 19. Mathews F, Lovett L, Rushton S, Macdonald DW (2006) Bovine tuberculosis in cattle: reduced risk on wildlife-friendly farms. Biology Letters 2: 271–274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. O'Brien DJ, Schmitt SM, Berry DE, Fitzgerald SD, Vanneste JR, et al. (2004) Estimating the true prevalence of Mycobacterium bovis in hunter-harvested white-tailed deer in Michigan. Journal of Wildlife Diseases 40: 42–52. [DOI] [PubMed] [Google Scholar]
- 21.Okafor CC, Grooms DL, Bruning-Fann CS, Averill JJ, Kaneene JB (2011) Descriptive epidemiology of bovine tuberculosis in Michigan (1975–2010): lessons learned. Veterinary Medicine International 2011. [DOI] [PMC free article] [PubMed]
- 22.Hughey BD (2003) Are there "hot spots" of bovine tuberculosis in the free-ranging white-tailed deer ( Odocoileus virginianus ) herd of northeastern Michigan? Thesis. East Lansing: Michigan State University. 1–86 p. [Google Scholar]
- 23. Okafor CC, Grooms DL, Bruning-Fann CS, Averill JJ, Kaneene JB (2011) Descriptive epidemiology of bovine tuberculosis in Michigan(1975-2010): lessons learned. Veterinary Medicine International 2011: 874924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Eberhardt LL (1960) Estimation of vital characteristics of Michigan deer herds. East Lansing: Department of Conservation.
- 25.Creed WA, Haberland F, Kohn BE, McCaffery KR (1984) Harvest management: the Wisconsin experience. In: Halls LK, editor. White-tailed deer ecology and management. Harrisburg.
- 26. Phillips CJC, Foster CRW, Morris PA, Teverson R (2002) Genetic and management factors that influence the susceptibility of cattle to Mycobacterium bovis infection. Animal Health Research Reviews 3: 3–13. [DOI] [PubMed] [Google Scholar]
- 27.Mitscherlich E, Marth EH (1984) Microbial survival in the environment: bacteria and rickettsiae important in human and animal health. Berlin: Springer-Verlag. 1–802 p. [Google Scholar]
- 28. Miller R, Kaneene JB (2006) Evaluation of historical factors influencing the occurrence and distribution of Mycobacterium bovis infection among wildlife in Michigan. American Journal of Veterinary Research 67: 604–615. [DOI] [PubMed] [Google Scholar]
- 29. Phillips CJC, Foster CRW, Morris PA, Teverson R (2003) The transmission of Mycobacterium bovis infection to cattle. Research in Veterinary Science 74: 1–15. [DOI] [PubMed] [Google Scholar]
- 30.USDA NRCS (2007) Soil Data Viewer 5.2 User Guide. United States Department of Agriculture, Natural Resources Conservation Science.
- 31.Banerjee S, Carlin BP, Gelfand AE (2004) Hierarchical modeling and analysis for spatial data. New York: Chapman and Hall/CRC. 1–448 p. [Google Scholar]
- 32.Gelman A, Carlin JB, Stern HS, Rubin DB (2004) Bayesian data analysis. New York: Chapman and Hall/CRC. 1–696 p. [Google Scholar]
- 33. Spiegelhalter DJ, Best NG, Carlin BP, van der Linde A (2002) Bayesian measures of model complexity and fit. Journal of the Royal Statistical SocietySeries B (Statistical Methodology) 64: 583–639. [Google Scholar]
- 34.Spiegelhalter D, Thomas A, Best N, Lunn D (2003) WinBUGS Version 1.4 user manual. Cambridge: MRC Biostatistics Unit. 1–60 p. [Google Scholar]
- 35. Smith BJ (2007) boa: an R package for MCMC output convergence assessment and posterior inference. Journal of Statistical Software 21: 1–37. [Google Scholar]
- 36. Gelman A, Rubin DB (1992) Inference from iterative simulation using multiple sequences. Statistical Science 7: 457–472. [Google Scholar]
- 37. Brooks SP, Gelman A (1998) General methods for monitoring convergence of iterative simulations. Journal of Computational and Graphical Statistics 7: 434–455. [Google Scholar]
- 38. Eberly LE, Carlin BP (2000) Identifiability and convergence issues for Markov chain Monte Carlo fitting of spatial models. Statistics in Medicine 19: 2279–2294. [DOI] [PubMed] [Google Scholar]
- 39.Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. New York: Springer-Verlag. 1–488 p. [Google Scholar]
- 40. Miller R, Kaneene JB, Fitzgerald SD, Schmitt SM (2003) Evaluation of the influence of supplemental feeding of white-tailed deer ( Odocoileus virginianus ) on the prevalence of bovine tuberculosis in the Michigan wild deer population. Journal of Wildlife Diseases 39: 84–95. [DOI] [PubMed] [Google Scholar]
- 41. Knust BM, Wolf PC, Wells SJ (2011) Characterization of the risk of deer-cattle interactions in Minnesota by use of an on-farm environmental assessment tool. American Journal of Veterinary Research 72: 924–931. [DOI] [PubMed] [Google Scholar]
- 42. Brook RK (2010) Incorporating farmer observations in efforts to manage bovine tuberculosis using barrier fencing at the wildlife-livestock interface. Preventive Veterinary Medicine 94: 301–305. [DOI] [PubMed] [Google Scholar]
- 43. Besag J, York J, Mollie A (1991) Bayesian image restoration, with two applications in spatial statistics. Annals of the Institute of Statistical Mathematics 43: 1–59. [Google Scholar]
- 44. Imrie CE, Korre A, Munoz-Melendez G (2009) Spatial correlation between the prevalence of transmissible spongiform diseases and British soil geochemistry. Environmental Geochemistry and Health 31: 133–145. [DOI] [PubMed] [Google Scholar]
- 45. Fine AE, Bolin CA, Gardiner JC, Kaneene JB (2011) A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan, USA. Veterinary Medicine International 2011: 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Witmer G, Fine AE, Gionfriddo J, Pipas M, Shively K, et al. (2010) Epizootiological survey of bovine tuberculosis in northern Michigan. Journal of Wildlife Diseases 46: 368–378. [DOI] [PubMed] [Google Scholar]
- 47.Fine AE, O'Brien DJ, Winterstein SR, Kaneene JB (2011) An effort to isolate Mycobacterium bovis from environmental substrates during investigations of bovine tuberculosis transmission sites (cattle farms and wildlife areas) in Michigan, USA. ISRN Veterinary Science. [DOI] [PMC free article] [PubMed]
- 48. Duffield BJ, Young DA (1985) Survival of Mycobacterium bovis in defined environmental conditions. Veterinary Microbiology 10: 193–197. [DOI] [PubMed] [Google Scholar]
- 49. Young JS, Gormley E, Wellington EMH (2005) Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil. Applied and Environmental Microbiology 71: 1946–1952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Jackson R, de Lisle GW, Morris RS (1995) A study of the environmental survival of Mycobacterium bovis on a farm in New Zealand. New Zealand Veterinary Journal 43: 346–352. [DOI] [PubMed] [Google Scholar]
- 51. Miller R, Kaneene JB, Schmitt SM, Lusch DP, Fitzgerald SD (2007) Spatial analysis of Mycobacterium bovis infection in white-tailed deer ( Odocoileus virginianus ) in Michigan, USA. Preventive Veterinary Medicine 82: 111–122. [DOI] [PubMed] [Google Scholar]
- 52. O'Brien DJ, Schmitt SM, Fitzgerald SD, Berry DE, Hickling GJ (2006) Managing the wildlife reservoir of Mycobacterium bovis: The Michigan, USA, experience. Veterinary Microbiology 112: 313323. [DOI] [PubMed] [Google Scholar]